

Line × Tester analysis for morpho-physiological traits of Zea mays L. seedlings
Qurban Ali1,2, Arfan Ali2*, Muhammad Ahsan1, Sajed Ali2, Nazar Hussain Khan1, Sher Muhammad4, Hafiz Ghazanfar Abbas3, Idrees Ahmad Nasir2, Tayyab Husnain2
Adv. life sci., vol. 1, no. 4, pp. 242-253, August 2014
*- Corresponding Author: Arfan Ali (Email: Arfan.ali@cemb.edu.pk)
Author Affiliations
2- CEMB, University of Punjab, Pakistan
3- Cotton Research Institute, Ayub Agricultural Research Institute – Faisalabad
4- CABB, University of Agriculture Faisalabad – Pakistan
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Zea mays L. is one of the most imperative cereal crop in world after wheat and rice. It is a used as food for human and feed for livestock. To meet the ever increasing demand, maize production can be increased by application of improved agronomic techniques to get varieties with higher qualitative and quantitative traits and resilience to abiotic stresses.
Methods: The genetic material was comprised of 8 parents and 12 F1 hybrids. The genotypes were sown in the iron treys filled with sand in three replications following completely randomized design. The data was recorded for fresh root length (FRL), fresh shoot length (FSL), fresh root-to-shoot length ratio (FRSLR), fresh root weight (FRW), fresh shoot weight (FSW), total fresh weight (TFW), fresh root-to-shoot weight ratio (FRSWR), dry root weight (DRW), dry shoot weight (DSW) and total dry weight (TDW), dry root-to-shoot weight ratio (DRSWR), chlorophyll contents (Ch.C), leaf temperature (LT), transpiration rate (E), photosynthetic rate (A), stomata conductance (gs), water use efficiency (WUS) and sub-stomata CO2 concentration (Ci). GCA (general combining ability) and SCA (specific combining ability) were calculated by using Kepmthorn, (1957) technique.
Results: Higher GCA of B-336 variety was recorded for FRL, FRW, FSW, TFW, A and Ci. Higher SCA of EV-1097Q × Pop/209, Sh-139 × Pop/209, EV-1097Q × B-316 and Sh-139 × B-316 varieties was recorded for FRL, FSL, FRW, FSW, TFW, A, Ci, WUE, LT, E and gs.
Conclusion: Higher heritability, genetic advance, GCA and SCA had decisive role in selection of drought tolerant maize varieties. F1 hybrids EV-1097Q × Pop/209, Sh-139 × Pop/209, EV-1097Q × B-316 and Sh-139 × B-316 showed higher SCA for all traits that persuaded that these hybrids may be used for higher grain and fodder yield under drought conditions.
Key words: Heritability, Genetic advance, GCA, SCA, Line × Tester analysis, Zea mays
Introduction
Maize (Zea mays L.) is one of the most important cereal crop in world after wheat and rice. It is grown in Pakistan as a major cash crop under the area of 1.083 million hectares with production of 3990 thousand tones [1]. Maize is dual propose crop, used as a food for human and feed for livestock. It is also used as a raw material in medicine, food and textile industries. Maize was exported about 300 million tons during the year of 2013-2014. It contains 72 % starch, 10 % protein, 4.80 % oil, 9.50 % fiber, 3.0 % sugar, and 1.70 % ash [2]. There are many traits involved in the improvement of grain and fodder yield of maize and by improving those traits yield may be increased. Various morphological traits like fresh root weight (FRW), dry root weight (DRW), fresh shoot weight (FSW), dry shoot weight (DSW), fresh root length (FRL), fresh shoot length (FSL), and physiological traits like photosynthetic rate (A), chlorophyll contents (Ch.C), transpiration rate (E), leaf temperature (LT), stomata conductance (gs), water use efficiency (WUE) and sub-stomata CO2 concentration (Ci) play an important role in yield [3-7]. The present study was conducted to evaluate various maize parents and F1 hybrids for morpho-physiological traits at seedling stage for their general combining ability (GCA) and specific combining ability (SCA).
Methods
The present study was conducted in the glasshouse of the Department of Plant Breeding and Genetics, University of Agriculture Faisalabad, Pakistan during cropping season of spring 2012-13. The genetic material was comprises of B-316 and POP/209 (male), B-11, B-327, Raka-poshi, B-336, Sh-139 and EV-1097Q (females) and their F1 developed by using 6 × 2 Line × Tester analysis design viz., B-11 x Pop/209, B-336 x Pop/209, EV-1097Q x Pop/209, B-327 x Pop/209, Raka-poshi x Pop/209, Sh-139 x Pop/209, B-11 x B-316, B-336 x B-316, EV-1097Q x B-316, B-327 x B-316, Raka-poshi x B-316 and Sh-139 x B-316. The genotypes were sown in the iron treys filled with sand in three replications following completely randomized design. The data was recorded for fresh root length, fresh shoot length (by using Vernier Caliper (Model RS232)), fresh root-to-shoot length ratio, fresh root weight, fresh shoot weight, total fresh weight, fresh root-to-shoot weight ratio, dry root weight, dry shoot weight and total dry weight, dry root-to-shoot weight ratio (by using electronic balance (OHAUS-GT4000, USA), chlorophyll contents (by using Chlorophyll meter), leaf temperature (by using Infrared leaf meter), transpiration rate, photosynthetic rate, stomata conductance, water use efficiency and sub-stomata CO2 concentration (by using IRGA, Infrared Gas Analyzer). The data was subjected for analysis of variance. GCA and SCA were calculated by using Kepmthorn technique [8]. The genetic advance was calculated by using Falconer formula [9].
Results
It is swayed from table 1 that significant differences were found for parents and F1 hybrids, and it is cleared that higher genotypic and phenotypic variance was recorded for sub-stomata CO2 concentration (104207.33, 104305.33) and water use efficiency (256.213, 343.333) respectively, genotypic (1778.151, 1778.987) and phenotypic (360.909, 417.787) coefficients of variances, respectively were also higher followed by photosynthetic rate (61.450, 63.772) and (288.324, 293.721) respectively. The range of heritability was found to be 50.0% to 99.90%. The genetic advance was found to be 7.29% to 478.12%.
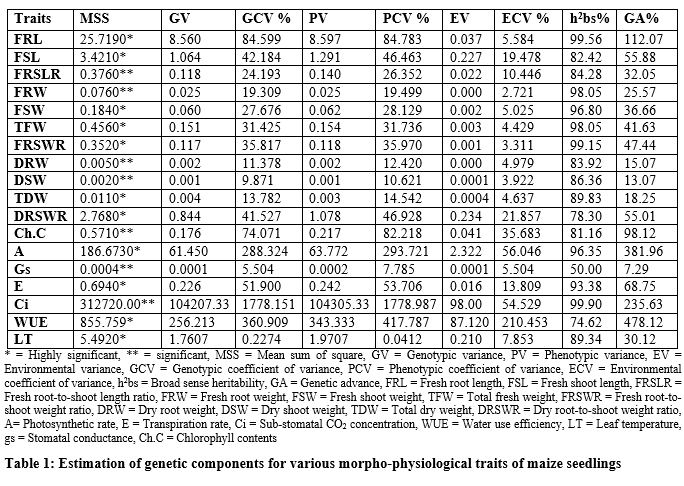
Combining ability analysis
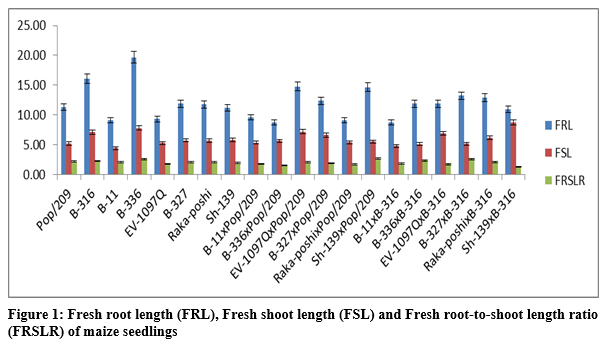
It is persuaded from tables 2 and 2a that the significant differences were found for parents and F1 hybrids. The figure1 indicated that higher fresh root length was recorded for B-336 and B-316 from parents while from F1 hybrids EV-1097Q x Pop/209 and Sh-139 x Pop/209 followed by B-327 x B-316 and Raka-poshi x B-316. The higher GCA for fresh root length was recorded for B-336 (2.0022) and Raka-poshi (0.9822) followed by B-316 (0.4106) and Sh-139 (0.3689).
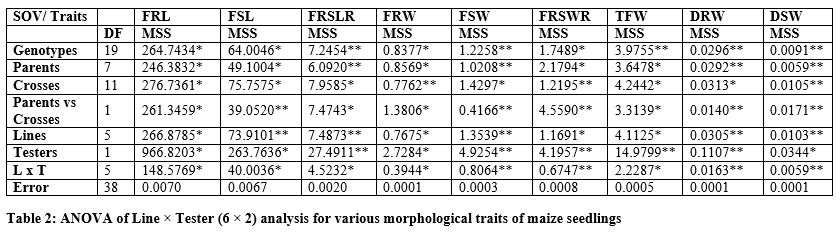
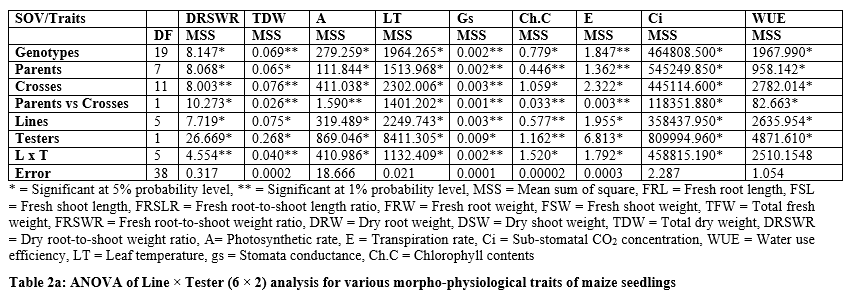
The higher SCA effects were recorded for EV-1097Q x Pop/209 (8.5078) and Sh-139 x B-316 (8.1344) followed by EV-1097Q x B-316 (7.6078) and Sh-139 x Pop/209 (7.0433). The additive effects were found to be negative as -1.0088 while very higher dominance effects (600.8853) were found for fresh root length. The contribution of lines (females) was 510.5848 greater as compared to testers (males) and line x tester interactions (Table 3). The figure1 indicated that higher fresh shoot length was recorded for B-336 and B-316 from parents while from F1 hybrids EV-1097Q x Pop/209 and Sh-139 x B-316 followed by B-327 x Pop/209 and EV-1097Q x B-316.
The higher GCA for fresh shoot length was recorded for B-336 (0.8606) and Sh-139 (1.3922) followed by B-316 (0.0772). The higher SCA effects were recorded for EV-1097Q x Pop/209 (4.2161) and Sh-139 x B-316 (4.3950) followed by EV-1097Q x B-316 (4.2339) and Sh-139 x Pop/209 (3.7472). The additive effects were found to be negative as -1.1641 while very higher dominance effects (152.3269) were found for fresh shoot length. The contribution of lines (females) was 946.3039 greater as compared to testers (males) and line x tester interactions (table 3).

The figure1 indicated that higher fresh root-to-shoot length ratio was recorded for B-336 from parents while from F1 hybrids Sh-139 x Pop/209. The higher GCA for fresh root-to-shoot length ratio was recorded for EV-1097Q (0.2251) and Raka-poshi (0.1963) followed by B-316 (0.0902). The higher SCA effects were recorded for EV-1097Q x Pop/209 (1.3912) and Sh-139 x B-316 (1.2596) followed by EV-1097Q x B-316 (1.2235) and Sh-139 x Pop/209 (1.2947).
The additive effects were found to be as 2.7477 while very higher dominance effects (54.1903) were found for fresh root-to-shoot length ratio. The contribution of lines (females) was 500.8744 greater as compared to testers (males) and line x tester interactions (table 3). Figure 2 & 3 indicated that higher fresh and dry root weight was recorded for B-336 from parents while from F1 hybrids Raka-poshi x Pop/209, EV-1097Q x B-316 and Sh-139 x Pop/209 respectively. The higher GCA for fresh and dry root weight was recorded for EV-1097Q (0.1958), B-336 (0.0048), Sh-139 (0.0181) and Raka-poshi (0.1963) respectively.
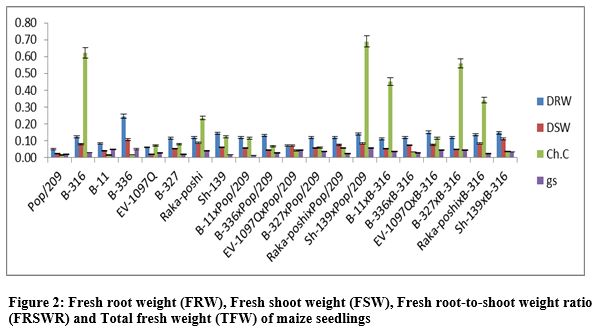
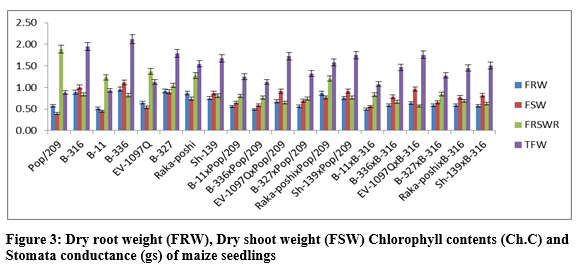
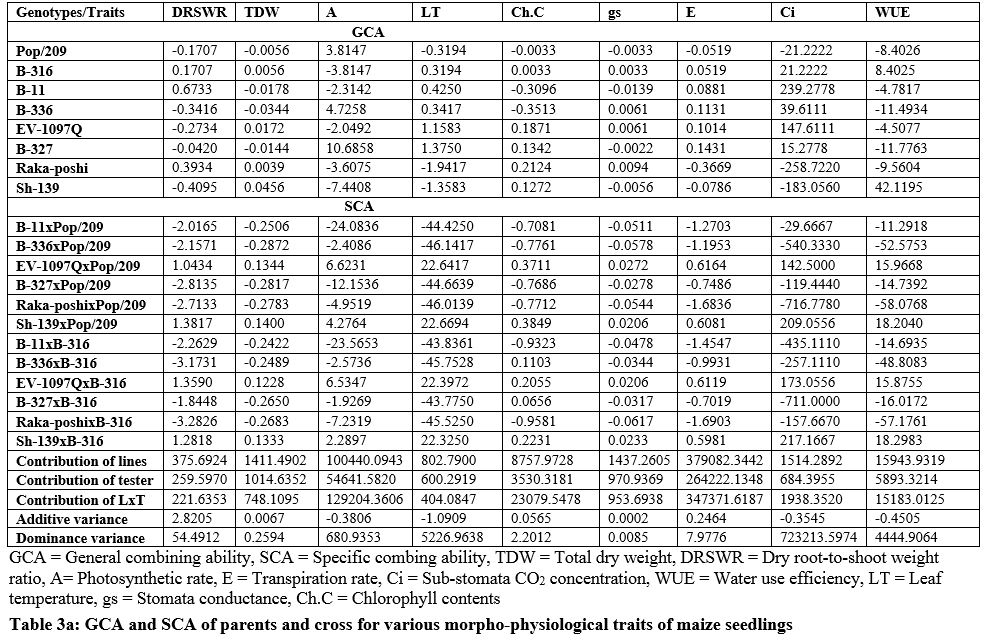
The higher SCA effects were recorded for EV-1097Q x Pop/209 (0.4401, 0.0819) and Sh-139 x B-316 (0.4349, 0.0853) followed by EV-1097Q x B-316 (0.3444, 0.0803) and Sh-139 x Pop/209 (0.4080, 0.0919) respectively. The additive effects were found to be 0.0732 while very higher dominance effects (2.7050) were found higher for fresh root weight as compared to dry root weight 0.0027 and 0.1065 respectively. The contribution of lines (females) was for dry root weight was 1087.4512 greater as compared to testers (males) and line x tester interactions (table 3) then the 2.77.9648 for fresh root weight. the development of hybrid seed. The figure 2 & 3 indicated that higher fresh and dry shoot weight was recorded for B-336 and B-316 from parents while from F1 hybrids EV-197Q x Pop/209, EV-1097Q x B-316, Raka-poshi x Pop/209 and Sh-139 x Pop/209 respectively. The higher GCA for fresh and dry shoot weight was recorded for EV-1097Q (0.0279, 0.0108) and Sh-139 (0.0684, 0.0275) respectively. The higher SCA effects were recorded for EV-1097Q x Pop/209 (0.6043, 0.0525) and Sh-139 x B-316 (0.5821, 0.0481) followed by EV-1097Q x B-316 (0.5419, 0.0425) and Sh-139 x Pop/209 (0.5257, 0.0481) respectively. The additive effects were found to be as 0.1407 while very higher dominance effects (5.0253) were found for fresh shoot weight as compared to dry shoot weight 0.0009 and 0.0343 respectively. The contribution of lines (females) was for fresh shoot weight was 1625.0848 greater as compared to testers (males) and line x tester interactions (table 3) then the 299.9012 for dry shoot weight. The figure 2 & 4 indicated that higher total fresh weight was recorded for B-336 and B-316 and total dry weight B-336 and B-11 from parents while from F1 hybrids Sh-139 x Pop/209 and EV-1097Q x B-316 for total fresh weightwhile B-11 x B-316 and Sh-139 x B-316 for total dry weight respectively.
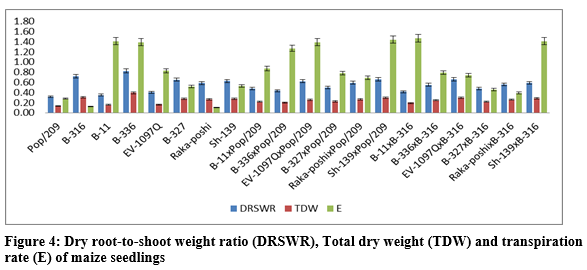
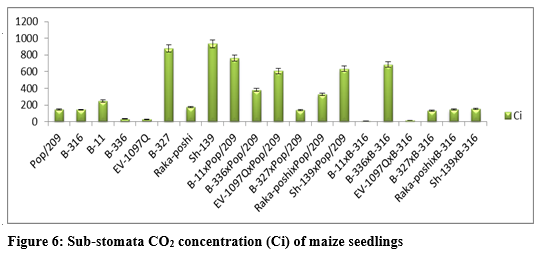
The higher GCA for fresh and dry shoot weight was recorded for EV-1097Q (0.2237, 0.0172) and Sh-139 (0.0374, 0.0456) respectively. The higher SCA effects were recorded for EV-1097Q x Pop/209 (1.0444, 0.1344) and Sh-139 x B-316 (1.0170, 0.1333) followed by EV-1097Q x B-316 (0.8863, 0.1228) and Sh-139 x Pop/209 (0.9338, 0.1400) respectively. The additive effects were found to be as 0.5890 while very higher dominance effects (17.3292) were found for total fresh weight as compared to total dry weight 0.0067 and 0.2594 respectively. The contribution of lines (females) was for total fresh weight was 1411.4902 greater as compared to testers (males) and line x tester interactions (table 3) then the 620.4837 for dry shoot weight. It is indicated in figure 3 that higher chlorophyll contents and stomata conductance was recorded for B-316 and B-11 from parents while from F1 hybrids Sh-139 x Pop/209 respectively. The higher GCA was recorded for EV-1097Q (0.1871, 0.0061) and Raka-poshi (0.2124, 0.0094) for chlorophyll contents and stomata conductance respectively. The higher SCA effects were recorded for EV-1097Q x Pop/209 (0.3711,0.0272) and Sh-139 x B-316 (0.2231, 0.0233) followed by EV-1097Q x B-316 (0.2055, 0.0206) and Sh-139 x Pop/209 (0.3849, 0.0206) respectively. The additive effects were found to be as 0.0565 while very higher dominance effects (2.2012) were found for chlorophyll contents as compared to stomata conductance 0.0002 and 0.0085 respectively. The contribution of lines (females) was for chlorophyll contents was 8757.9728 greater as compared to testers (males) and line x Sh-139 from parents while from F1 hybrids B-11 x B-316 and B-316 respectively. The higher GCA was recorded for EV-1097Q (0.1014, 147.6111), B-327 (0.1431, 15.2778), B-336 (0.1131, 39.6111) and B-11 (0.0881, 239.2778) for transpiration rate and sub-stomata CO2 concentration respectively. The higher SCA effects were recorded for EV-1097Q x Pop/209 (0.6164, 142.500) and Sh-139 x B-316 (0.5981, 217.1667) followed by EV-1097Q x B-316 (0.6119, 173.0556) and Sh-139 x Pop/209 (0.6081, 209.0556) respectively. The additive effects were found to be as 0.2464 while lower dominance effects (7.9776) were found for transpiration
rate as compared to sub-stomata CO2 concentration -0.3545 and 723213.5974 respectively. The contribution of lines (females) was for transpiration rate was 379082.3442 greater as compared to testers (males) and line x tester interactions (table 3) then the 1514.2892 for sub-stomata CO2 concentration. The figure 5 indicated that higher leaf temperature, water use efficiency and photosynthetic rate was recorded for Raka-poshi, Sh-139 and B-336 from parents while from F1 hybrids B-11 x B-316 for leaf temperature and Sh-139 x B-316 for water use efficiency and photosynthetic.
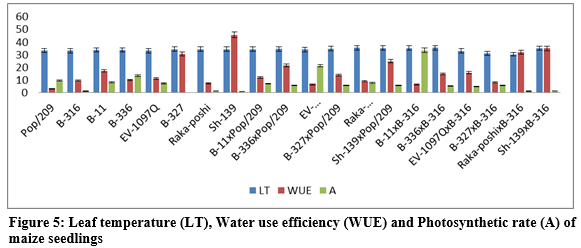
The higher GCA was recorded for B-327 (10.6858, 1.3750) for leaf temperature and photosynthetic rate respectively and Sh-139 (42.1195) for water use efficiency. The higher SCA effects were recorded for EV-1097Q x Pop/209 (6.6231, 22.6417, 15.9668) and Sh-139 x B-316 (2.2897, 22.3250, 18.2983) followed by EV-1097Q x B-316 (6.5347, 22.3972, 15.8755) and Sh-139 x Pop/209 (4.2764, 22.6694, 18.2040) respectively. The negative additive effects were found leaf temperature, water use efficiency and photosynthetic rate while higher dominance effects (5226.9638) were found for leaf temperature. The contribution of lines (females) was for photosynthetic rate was 100440.0943 greater as compared to testers (males) and line x tester interactions (table 3).
Discussion
The highest value of heritability was fond for fresh root length, fresh root and shoot weight, total fresh weight, photosynthetic rate and sub-stomata CO2 concentration. It was suggested that selection for drought tolerance maize genotypes may be helpful to improve grain yield [10-13]. The highest value of genetic advance was found for fresh root length, photosynthetic rate, transpiration rate, water use efficiency, and chlorophyll contents and sub-stomata CO2 concentration. The higher values of heritability and genetic advance indicated that selection of higher yielding maize genotypes on the basis of fresh root length, photosynthetic rate, transpiration rate, water use efficiency, chlorophyll contents and sub-stomata CO2 concentration may be helpful to improve maize yield [4,5,14-16]. GCA reflects the ability of genotypes to transmit traits in a general of series of crosses which indicated the additive gene effects while specific combining ability showed ability of genotypes to transmit traits in specific crosses.
Higher GCA of B-336 was recorded for fresh root length, fresh root and shoot weight, total fresh weight, photosynthetic rate and sub-stomata CO2 concentration. Higher SCA of EV-1097Q × Pop/209, Sh-139 × Pop/209, EV-1097Q × B-316 and Sh-139 × B-316 was recorded for fresh root and shoot length, fresh root and shoot weight, total fresh weight, photosynthetic rate and sub-stomata CO2 concentration, water use efficiency, leaf temperature, transpiration rate, stomata conductance, fresh and dry root-to-shoot weight ratio and chlorophyll contents. The higher GCA indicated the genotypes are good general combiner and may be used for the development of hybrid seed. The higher value of SCA and dominance effects indicated that the hybrids may be used for better yield [1,10,14,17].
References
- Ali Q, Ahsan M, Tahir MHN, Basra SMA. Genetic evaluation of maize (Zea mays L.) accessions for growth related seedling traits. International Journal for Agro Veterinary and Medical Sciences, (2012); 6(3): 164-172.
- Ali Q, Ashraf M, Anwar F. Seed composition and seed oil antioxidant activity of maize under water stress. Journal of the American Oil Chemists' Society, (2010); 87(10): 1179-1187.
- Ahsan M, Hussain M, Farooq A, Khaliq I, Farooq J, et al. Physio-genetic behavior of maize seedlings at water deficit conditions. Cercetari agronomice in Moldova (Romania), (2011).
- Ahsan M, Farooq A, Khaliq I, Ali Q, Aslam M, et al. Inheritance of various yield contributing traits in maize (Zea mays L.) at low moisture condition. African Journal of Agricultural Research, (2013); 8(4): 413-420.
- Ali Q, Elahi M, Ahsan M, Tahir MHN, Basra SMA. Genetic evaluation of maize (Zea mays L.) genotypes at seedling stage under moisture stress. International Journal for Agro Veterinary and Medical Sciences, (2011); 5(2): 184-193.
- Moulin S, Baret F, Bruguier N, Bataille C. Assessing the vertical distribution of leaf chlorophyll content in a maize crop; 2003. IEEE. pp. 3284-3286.
- Welcker C, Andréau B, De Leon C, Parentoni S, Bernal J, et al. Heterosis and combining ability for maize adaptation to tropical acid soils. Crop science, (2005); 45(6): 2405-2413.
- Kempthorne O. An introduction to genetic statistics. (1957).
- Atchley WR, Hall BK. A model for development and evolution of complex morphological structures. Biological Reviews, (1991); 66(2): 101-157.
- Ali Q, Ahsan M, Mustafa HSB, Hasan E. Studies of genetic variability and correlation among morphological traits of maize (Zea mays L.) at seedling stage. Albanian Journal of Agricultural Sciences, (2013); 12(3): 405-410.
- Mehmood A, Saleem M, Khabir A. Evaluation of heterosis for grain yield and some agronomic characters in maize hybrids between parents of diverse genetic origin. Sarhad Journal of Agriculture (Pakistan), (2003).
- Asrar-ur-Rehman Saleem US, Subhani GM, Ahmad N, Rahim M, Ali MA, et al. Correlation and path coefficient analysis in maize (Zea mays L.). Journal of Agriculture Research, (2007); 45(3).
- Xiang K, Yang K, Pan G, Reid L, Li W, et al. Genetic diversity and classification of maize landraces from China's Sichuan Basin based on agronomic traits, quality traits, combining ability and SSR markers. Maydica, (2010); 55(1): 85.
- Ali Q, Ahsan M, Khan NH, Ali F, Elahi M, et al. Genetic analysis for various quantitative traits of chickpea (Cicer arietinum L.). International Journal for Agro Veterinary and Medical Sciences, (2012); 6(1): 51-57.
- Ali Q, Ahsan M, Ali F, Aslam M, Khan NH, et al. Heritability, heterosis and heterobeltiosis studies for morphological traits of maize (Zea mays L.) seedlings. Advancements in Life Sciences, (2013); 1(1).
- Muraya M, Ndirangu C, Omolo E. Heterosis and combining ability in diallel crosses involving maize (Zea mays) S1 lines. Animal Production Science, (2006); 46(3): 387-394.
- Ali Q, Ahsan M, Ali F. Genetic advance, heritability, correlation, heterosis and heterobeltiosis for morphological traits of maize (Zea mays L). Albanian Journal of Agricultural Sciences, (2013); 12(4): 689-698.