

Full Length Research Article
Possible Approaches for the Origin of Life in Hot Mineral Water in Primary Hydrosphere
Ignat Ivanov Ignatov*1, Teodora Petrova Popova
Adv. life sci., vol. 12, no. 2, pp. 399-412, May 2025
*– Corresponding Author: Ignat Ivanov Ignatov (Email: mbioph@abv.bg)
Authors' Affiliations
2. Faculty of Veterinary Medicine, University of Forestry, Sofia 1756 – Bulgaria
[Date Received: 20/10/2024; Date Revised: 05/01/2025; Date Available Online: 31/08/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Protostructure emerges under corona discharge as ions and water molecules self-organize into stable formation. With 45.6 % density relative to living cells, this structure represents a primitive stage of biochemical evolution. Their formation follows Prigogine’s principle of entropy reduction, illustrating the potential role of early Earth’s reducing atmosphere and mineral surfaces in catalyzing the origin of complex biochemical systems.
Methods: Using hot mineral water from Rupite, Bulgaria, we conducted experiments involving corona electric discharge (1–30 kHz, 12 kV) to simulate prebiotic conditions. Fourier Transform Infrared (FT-IR) spectroscopy was employed to analyze the water's inorganic and organic composition. Additionally, we assessed the oxidation-reduction potential (ORP) and measured changes in physicochemical parameters under controlled conditions.
Results: The protostructure was created in laboratory conditions in hot mineral water under corona electric discharge. Self-organization estimation using Prigogine’s principle marks an early stage of biochemical evolution, emphasizing the role of the primordial atmosphere and hydrosphere in life's origin. The structure density was 45.6% of that of living cells. We quantitatively estimated the results of Miller’s and Wilson’s experiments. The biochemical reactions were described with the compounds as methane, ammonia, etc.
Conclusion: The findings support the hypothesis that hot mineral water in the primary hydrosphere, enriched with ions, exposed to electric discharge in the primary atmosphere, and with contact medium with the hydrosphere, providing optimal conditions for synthesizing organic compounds and forming protostructures. These results reinforce the concept of geothermal systems of life and highlight the significance of mineral water’s physicochemical properties in fostering prebiotic chemistry.
Keywords: Origin of life; Hot mineral water; Electric discharge; Protostructure
Introduction![]()
The origin of life on Earth most likely occurred in an aquatic environment. For the related processes, suitable building materials and energy are required. There are three possible sources of energy for living cells: Chemical, Solar, and Geothermal. Biogenic elements are basic components of living organisms [1]. These include: primary elements (O, C, H, N, Ca and P), involved in the composition of basic molecules of living matter, secondary elements (Na, K, Cl, S, Mg and F), which exist as salts, inorganic ions and parts of essential molecules and microelements or trace elements (I, Cu, Zn, Mo, Se, and Co), present in very scarce amounts, but key to the functioning of cells. Inorganic and organic biological compounds are formed from biogenic elements. The primary inorganic substance and solvent in the cell is water. Organic biological compounds include proteins, carbohydrates, lipids, and nucleic acids [2, 3].
Our planet's hydrosphere had been formed at a very early stage of its development, during accretion. Water was primarily found in the oceans, while CO2 was mainly located in sediments and the atmosphere, where N2 and CO2 predominated [4]. The atmosphere and hydrosphere most likely developed through the gradual release of H2O, CO2, HCl, N2, CO, and H2S from the Earth. The seawater composition with Cl, S, Na, K, Ca, and Mg content had been highly stable. The partial pressure of CO2 in the atmosphere did not change significantly due to its high solubility in seawater. The Na+, K+, and Mg2+ concentrations have been suggested to be controlled by exchange reactions between seawater and Al-silicates [5]. Even in the earliest stages of planetary evolution, liquid water, due to its specific chemical and physical properties, provided an environment where chemical reactions between atmospheric CO2 and minerals occurred [6].
Water is the primary substance in the living cell. Its content is 70-98% of the mass of both prokaryotic and eukaryotic cells. It is a medium where various chemical and biochemical processes occur, including the dissociation of electrolytes. Water is the source of hydrogen and hydroxyl ions, which are involved in numerous chemical reactions, including those that occur in living cells. Additionally, water serves as a dispersion medium for colloidal solutions of proteins and carbohydrates. It is a solvent of crystalloid substances in the cell [7].
In 1953, Miller conducted electrosynthesis experiments with compounds from the primordial atmosphere, forming organic molecules [8]. Different researchers repeated the experiments [9–12]. They reviewed recent advances in modeling plasmas and ionized gases with compositions similar to those of the primordial atmosphere. These authors analyzed the importance of plasma kinetics for processes in this atmosphere, which include elementary processes by which free electrons activate weakly reactive molecules, such as carbon dioxide or methane, thus potentially starting prebiotic reaction chains. Electron-molecular reactions and energy exchange between molecules are activated. The primordial atmosphere was highly ionized by electrical activity, photoionization, or meteor phenomena. Plasma kinetics are complex due to the non-equilibrium characteristics of the energy distribution. The authors draw attention to the electron velocity and vibrational distribution functions, which can provide insight into prebiotic processes initiated by collisions between electrons and molecules. Here, the degree of ionization of the gas, the energy distribution of free electrons, and the formation of the simplest reactive species are crucial, serving as the basis for the subsequent complex biochemical processes and their subsequent role in forming the first prebiotic species [12]. Gan et al., also highlighted the critical role of discharge plasmas in creating the chemical prerequisites for the emergence of life on early Earth, particularly with their potential to simulate prebiotic conditions. They have activated the synthesis of the building blocks of life, specifically the vital organic molecules–amino acids, nucleotides, carbohydrates, and lipids [13].
The investigation of the formation of amino acids in a neutral atmosphere by electric discharge was performed [14]. From 2013 to 2016, Ignatov and Mosin conducted corona discharge electrosynthesis experiments at the dielectric-water-air interface [15]. These experiments also show the possibility of an electric discharge from the atmosphere or a dielectric medium passing into the water. Hot mineral water from Rupite, Bulgaria, was studied, with a temperature of 74.6 °C at the source and 55.4 °C at one of the ponds. Formation of protostructures has also been detected [16]. Today, this hot mineral water is a habitat for some extremophiles. Derekova et al. isolated a new thermophilic, strictly aerobic, Gram-positive, spore-forming chemoorganotrophic bacterium from three hot springs in the Rupite Basin region, Bulgaria, named Anoxybacillus rupiensis [17].
The results of various researchers indicate that electrosynthesis most likely took place in the primary anoxic hydrosphere of the Earth, consisting of a mixture of water and gases – H2, CH4, NH3, and CO, subjected to the action of high-energy electric discharges (lightning). The experiments of Ignatov and Mosin also demonstrate that the most favorable conditions for the emergence of life and the maintenance of biochemical reactions are hot mineral water interacting with CaCO3, where processes of thermal polymerization can occur, leading to the formation of proto-aggregates [18]. Their studies of seawater near hot mineral water showed an increased likelihood of the first living organisms forming in hot, mineral-rich water [10].
Szostak points out that the accumulation of concentrated chemical compounds is essential for structuring protocells during the genesis of living matter [19]. It indicates that lakes in geothermally active areas have a variable temperature [20]. They can even be cooled on the surface and on land by melting ice.
In 2009, Mulkidjanian and Galperin analyzed the cell cytoplasm and highlighted the content of potassium, zinc, magnesium, and phosphate ions. They are not so common in sea and ocean water. This suggests that the cell membrane originated in freshwater pools [21]. Later, Damer and Deamer also linked the cell membrane composition to freshwater lakes with water from geothermal springs and rain in the primary hydrosphere [22].
Mineral water is of particular interest in terms of biogenesis from our point of view. The presence of several ions, such as calcium, sulfur, iron, magnesium, etc., is a prerequisite for forming water clusters. The clusters can form polymers with each other under certain conditions. Clusters and their polymers are likely a matrix for forming organic molecules with organogenic chemical elements. The main ones are carbon and nitrogen, but also hydrogen and oxygen. The available methane, ammonia, carbon dioxide, etc., in the primary atmosphere might be the source of these necessary elements. The energy source for these synthesis processes could be quanta of sunlight, geothermal or chemical energy from the breakdown of macromolecules. The water clusters formed around calcium or other ions may underlie the formation of organic molecules in the presence of methane and carbon dioxide from the primary atmosphere. They can also serve as the basis for forming organic molecules, biopolymers, and simple biological lipoprotein membranes (archaebacterial membranes are single-layered). Research indicates that water clusters form around positively charged metal ions. They are stable over time. Their sizes are from 1.3 to 1.5 nm. The primary layer consists of 6 water molecules, and the secondary layer comprises 24. The formation of clusters of water molecules around metal ions with the following general formulas has been demonstrated: calcium Ca+(H2O)n [23-25], magnesium Mg2+(H2O)n [26], and zinc Zn2+(H2O)n [27]. [H2O]n water clusters are also formed around negative ions such as CO32- [28], SO42- [29], and PO43- [30]. A layer of water molecules forms around the negative ions. Certain publications also show stability results over water clusters with anions and cations over time [23–30]. Data on a Gaussian distribution and a linear function for small clusters around ions have also been analyzed [31]. An analysis was also made with dissolved molecular hydrogen and hydrogen ions [32].
Various researchers have carried out experiments to structure proteinoids. They were defined in 1950 by Fox. Adamatzky et al., have reviewed studies on structuring proteinoids [33]. They described in detail the protostrutures received by Ignatov and Mosin in hot mineral water under coronal electric discharge conditions [33]. Adamatzky suggested that proteinoids with primary amino acid constituents could be computer-modeled [34]. His idea was inspired by Harada and Fox, who demonstrated that proteinoids oscillate electrically, similar to neuronal spiking activity [35]. Liquid water allows hydrocarbons and molecules that contain carbon, hydrogen, oxygen, nitrogen, and sulfur to be stable for a long time. Liposomes are also formed. These formations are thermally stable and resistant to degradative changes [36]. A team led by Sugawara, Kurihara, et al., has successfully structured protocells from fatty acids and nucleotides [37]. DNA replication had also been successful, as the medium temperature was reduced from 95 to 65 °C. Artificial DNA has been created using nano technological methods [38].
In this article, we examine processes related to the emergence of organic compounds and viable structures in open lakes with warm and hot mineral water. They have the following natural features that are important in this aspect: Open mineral lakes contact the primary atmosphere with large surfaces; The electrical gas discharge from the atmosphere interacts with the surface of the lakes; At the spring, there is an electrical gradient between the earth's layers; Geothermal activity is constant over time; Mineral water cools on contact with land.
The present study aims to analyze conditions in the primary hydrosphere, such as properties of water clusters, electric potentials at water wells, at the land boundary, as well as chemical reactions with the primary hydrosphere.
![]() Methods
Methods
Mineral water from the Rupite basin in Bulgaria was used in the research. The experiments were made with laboratory gaseous conditions of the Miller-Urey experiments [8].
Control Setup. Deionized water was used as a control to ensure baseline comparisons.
The corona color spectral analysis method is described by [39, 40], and the stable conditions for electric processes for the origin of life were studied with frequencies up to 1–30 kHz, U=12 kV.
Oxidation-reduction potential. The electric parameters of water, including ORP and pH, were measured using an advanced electrochemical meter equipped with precise sensors. The device was calibrated before each experiment to ensure measurement accuracy. The Range of the HANNA Instruments HI221 meter is pH (2.00–16.00 ±0.01) and ORP (±699.9±0.01 – ±2000±0.1) mV.
Fourier Transform Infrared (FTIR) spectroscopy. The samples were analyzed using a Fourier-transform infrared (FT-IR) spectrometer. The FT-IR spectra in our study were recorded with a “Thermo Nicolet Avatar 360 FT-IR” spectrometer (Waltham, MA, USA), equipped with a DTGS detector in the region 400–4000 cm-1, by accumulating 64 scans at a spectral resolution of 2 cm-1. Experiments were performed at 298.15 K. All measurements were conducted at the Institute of General and Inorganic Chemistry, Bulgarian Academy of Sciences (BAS), Sofia, Bulgaria [42].
Determination of Total Dissolved Solids (TDS)
A custom script was used to analyze the data, ensuring high precision and reproducibility in determining dissolved solid concentrations in the samples. Program processing was performed by Python program.
Primary atmosphere as a source of reduction compounds
The primordial atmosphere had been saturated with ammonia (NH3), methane (CH4), carbon dioxide (CO2), and hydrogen (H2) [5, 6]. Ammonia, methane, and carbon dioxide have reducing properties when dissolved in water, a prerequisite for chemical reactions forming organic molecules. The chemical reaction in prebiotic chemistry to produce formaldehyde (HCHO) from CO2 and H2 had been possible.
The formation of amino acids as glycine with reducing agents in water proceeds in principle as follows:
CH4 +NH3 + H2O + H2 → NH2CH2COOH + other products (1).
Formation of molecular hydrogen (H2) in the nature
According to the data of Klein al, (2020), the main non-biological processes in nature in which hydrogen is formed are the following: Oxidation of iron in minerals; Degassing of the magma at low pressure; Reaction of water with surface radicals during mechanical destruction of silicate rocks [43]. Hydrogen is the most abundant element in the universe. The nucleus of the hydrogen atom consists of one positively charged proton. A negatively charged electron revolves around it. Two hydrogen atoms bond in the atmosphere into a molecule (H2). The redox reaction, in which electrons pass from one reactant to another, is particularly characteristic. Donating electrons is called oxidation, and accepting them is called reduction. They proceed in parallel (oxidation-reduction). A hydrogen atom tends to transfer an electron to an acceptor. On the contrary, the oxygen atom tends to accept an electron. When an electron is transferred from each of the two hydrogen atoms to the oxygen, the hydrogen is oxidized, and the oxygen is reduced. The reaction releases a significant amount of free energy as heat [44].
Layers with electrode potentials
Water is filtered through rocks and saturated with beneficial minerals, determining its physicochemical composition. In the depths of the earth, near the mineral springs, there are layers with significant differences in the electrode potentials, which act as an anode and cathode in an underground electrolyzer [45, 46]. In this way, the earth’s crust naturally produces activated ingredients. Also, Fe-S layers generate a natural electric potential when mineral water passes through them [47]. The electrical potential of mineral water is affected by porous and fractured media [48].
Basis reactions in hot mineral water
In our opinion, the main chemical reaction responsible for the formation of biopolymers in hot mineral water, which may have played an essential role in the biogenesis of the ancient Earth, is the follows:
Synthesis of amines by the reductive amination of aldehydes can be represented as follows:
RCHO + NH3 + H2→ RCH2NH2 + H2O (2)
Analysis of Miller's experiments on the mass and number of atoms in molecules in electrosynthesis
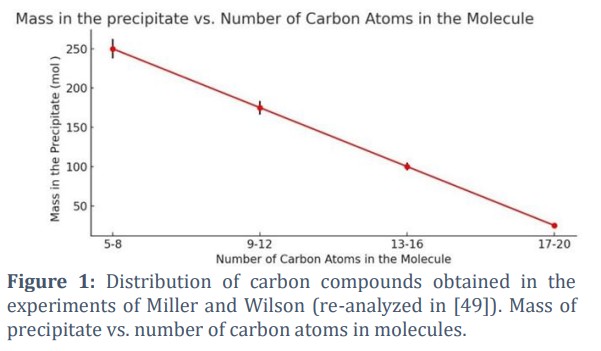
In 2015, Ignatov and Mosin [49] constructed a distribution of carbon compounds by performing a more comprehensive analysis of Miller and Urey’s experiments [8] and Wilson’s research [50] on the mass and number of atoms in molecules in electrosynthesis [50].
In 1953, Miller and Urey's experiments demonstrated that a mixture of CH4, NH3, H2, water vapor, amino acids, and other simple organic molecules could be synthesized under simulated early Earth conditions involving electric discharge and UV radiation. Among the identified products were 18 amino acids, including glycine, alanine, and aspartic acid, containing between 2 and 5 carbon atoms. These results suggest that the basic building blocks of life could naturally form in the primitive atmosphere and hydrosphere. In 1960, Wilson conducted experiments exploring water vapor, NH3, H2S, and yeast ash to electric spark discharges. The results showed the formation of larger organic polymer molecules containing 20 or more carbon atoms, representing a significant step toward synthesizing complex organic compounds. In 1964, Harada and Fox polymerized amino acids into short peptide chains by heating them to 95 °С, demonstrating a critical stage in the progression toward protein synthesis [35]. These experiments highlight that under various energy sources and chemical conditions, organic molecules can transition from simple compounds to more complex polymers, playing a crucial role in the evolution of life on Earth. The graph in Fig. 1 shows that the formation of molecules with different carbon chain lengths is diverse. In Fig. 1, the achieved linear relationship between the mass of the sediment and the number of carbon atoms in the molecule in Miller’s and Wilson’s experiments is presented. It is described in our previous research of Ignatov and Mosin [49].
Fig. 1 illustrates a linear regression with Residual Sum of Squares (RSS), and the formula is:
y=–12.5x+312.5
y – the mass of the precipitate (mol.%);
x – the average number of carbon atoms in each category
It shows how simpler molecules combine to form more complex ones under prebiotic conditions. The molecules with the highest percentage are the most stable and can be formed more easily under the given conditions. They have the smallest number of carbon atoms, from 5 to 8. Complex molecules of 17 to 20 carbon atoms are also formed, and these are the smallest percentage. A major conclusion is that more complex molecules can be formed in significant quantities. This supports the hypothesis that simple organic molecules could have evolved into more complicated compounds, which are decisive for the origin of life.
The results from Fig. 1 support the hypothesis that essential organic compounds could have formed under the natural conditions of early Earth. The observed distribution of carbon chain lengths suggests a balance between simplicity and complexity, potentially reflected by prebiotic chemical processes. According to Prigogine’s principle, self-organization and the decrease in entropy in open systems can explain the emergence of longer carbon chains. The linear model is shown in the graph, with a high R2 value, indicates that the variability in the data is well-explained, supporting the idea of organized chemical progression. While the Shannon entropy of 1.69 bits highlights moderate diversity, further analysis is required to connect this diversity to the formation of amphiphilic molecules, such as fatty acids and phospholipids, which are critical for prebiotic membranes. The graph provides a foundation for understanding how self-organization and polymerization processes might lead to complex prebiotic systems.
Physicochemical composition of the water from Rupite
Physicochemical composition of the water from Rupite is: Ca2+ at 32.8±1.7; Na+ at 531 ±53; Mg2+ at 15.6±0.7; HCO3− at 1533±76; SО42- at 81.3 ±4.1; Hydrosulfide HS– at 42±2 mg/ L.
The parameter of pH is 7.14±0.11, and the electrical conductivity is 2140 µS/cm.
Fourier Transform Infrared Spectroscopy (FT-IR)
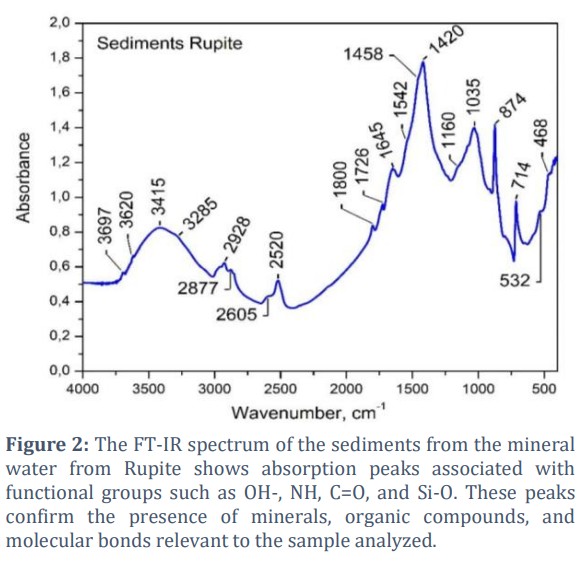
The analysis of Rupite, Bulgaria sediments using the Fourier Transform Infrared (FT-IR) spectroscopy is presented in Fig. 2.
The FT-IR spectroscopy reveals the presence of OH groups at 3620 cm-1 and 3697 cm-1, corresponding to water molecules or hydroxyl groups in minerals, which are essential for the adsorption of organic molecules on mineral surfaces. The 3415 and 3285 cm-1 peaks indicate stretching vibrations of NH, OH, and C-H bonds. At 1645 cm-1, the peak represents the presence of methyl and methylene groups, as well as lipids, suggesting the presence of organic material in the sample. At 1542 cm-1, C=O stretching is present in carbonyl groups commonly found in amide groups. In the water from Rupite, there are amide groups (C=O, N-H), proved at 1542 cm-1, 1458 cm-1, and 1420 cm-1. C-H stretches are observed in the spectrum's methyl and methylene groups, as well as in lipids. At 874, 714 cm-1, the C-O stretch in alcohols, esters, and hydrocarbons is demonstrated. Si-O bonds are also proven. The results show inorganic and organic compounds.
Water clusters with metal ions
Water clusters around anions and cations are structured in water, which has been described earlier [23–31, 51]. The primary layer comprises six water molecules, and the secondary layer comprises 24. We believe these clusters are the most likely matrix for biopolymer synthesis under primordial Earth conditions. We consider the following reaction, in which a ferrous ion participates in a redox reaction and accepts electrons, which are important in these processes: Fe3+ + e-→ Fe2+. [Fe (H2O)6]2+(H2O)24 is a theoretical model. Miliordos et al, (2015) found that 12.4-13.4 water molecules constituted the second solvation shell [52]. For Ca2+, the first shell should be n=1-8, and the second shell should be n=6–8. For Na+, the first shell is n=1–6 [53]. The data for HCO3− is (H2O)1−10 [54].
Water's hydrogen bonds give it unique physical and chemical properties of water. Liquid water has an anomalously high molar heat capacity (75 J/mol K). This means it can absorb relatively large amounts of solar energy during the day and release it at night without much temperature change. Due to highly polar water molecules and their mobility, water is an excellent and almost universal solvent for ionic compounds and polar organic molecules [55]. Electrostatic forces and the formation of electromagnetic interactions between the first and second layers influence the formation of clusters containing metal ions. In the earth’s bowels near mineral springs, there are layers with a significant difference in electrode potentials, which act as anode and cathode in an underground electrolyzer. Such are zinc and copper or potassium and nickel. In this case, zinc loses electrons, and copper gains them. Thus, the earth’s crust naturally produces activated ingredients [56]. The most unstable water clusters are formed around Ca2+, then around Zn2+, and are most stable with Mg2+ ions. The highest charge density is reported for magnesium due to the negligible density of magnesium ions.
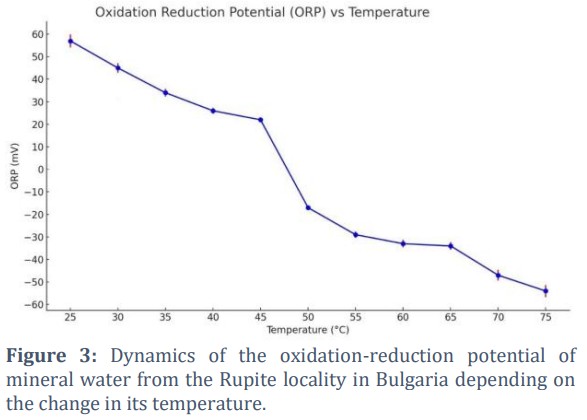
The results of our experiment with mineral water from Rupite, Bulgaria, which was cooled, demonstrated changes in its oxidation-reduction potential (ORP) in response to temperature fluctuations [57]. The atmosphere was similar to that in laboratory conditions, as observed in the Miller-Urey experiments, which involved methane, ammonia, hydrogen, and water vapor [8]. Fig. 3 shows the temperature and oxidation-reduction potential of the water being studied. With 10 measured samples, the result of the Student's t-test was p<0.05. The mineral water from Rupite shows a decrease in ORP with increasing temperature. This trend can be attributed to changes in the chemical activity of oxidizing and reducing ions. At temperatures above 50 °С, the ORP becomes negative, indicating predominantly reducing conditions. The presence of ions such as hydrosulfides (HS–), hydrogen carbonates (HCO3−), and possibly molecular hydrogen (H2) in the mineral water and the primordial hydrosphere is a key factor contributing to these reducing properties.
The results can be approximated with a cubic polynomial function of the kind:
forp(t) = 0.001226 t3 – 0.1696 t2 + 4.8579 t + 21.416
Water with ORP above zero mV is considered an oxidizing medium, while water with ORP below 0 mV is a reducing medium. In our measurements, a precise inversion of ORP from an oxidizing potential to a reducing potential was observed around 50 °С, marking the transition point where the medium changes from oxidative to reductive as temperature increases.
Key reductants from the primordial atmosphere – including hydrogen (H₂), methane (CH₄), ammonia (NH₃), and carbon monoxide (CO) played a significant role in early redox reactions in the gaseous phase. These compounds retained their reducing properties upon dissolving in water, fostering a chemically reducing environment in the primordial hydrosphere.
Such conditions were essential for prebiotic chemistry and the formation of organic molecules to affect entropy [58, 59].
These atmospheric compounds (H2, CH4, NH3, and CO) acted as reductants, driving essential redox reactions in the gaseous phase and aqueous solutions, thereby contributing to the reducing environment necessary for prebiotic chemical processes. As the temperature increases, ionic reactions in water are accelerated, enhancing the rate of redox reactions. This results in the faster depletion of available oxidants, further reinforcing the dominance of reductants.
Basic compound prerequisites for the formation of biopolymers
Kaolinite is a clay material with a surface that can adsorb water and other molecules. It was most likely also present in the primordial Earth. Glycine is an amino acid that can be adsorbed on kaolin, and polymerization can occur.
Glycine (NH2CH2CO2H) and methylamine (CH3NH2) are simple, structurally analogous water-soluble organic compounds related to each other by the presence or absence of the acid moiety (carboxyl group, −CO2H) in the Earth's biosphere. They have been found in numerous meteorites and comets, indicating that they were formed by processes before, during, and after the formation of the Solar System [60].
Glycine is the most straightforward non-essential amino acid in a living organism. Its molar weight is 75.1, and the R-group is a single hydrogen atom, making it the smallest amino acid. Simple substances found in living organisms are involved in the biosynthesis of many complex molecules, and glycine occupies a central position in metabolism as a precursor of nitrogenous compounds and several key metabolites of low molecular weight, such as glutathione, creatine, purines, porphyrins, etc. [61]. Additionally, glycine has surfactant properties [62]. The magnetic properties of the C-centered glycyl [NH2CHCOOH] point radical, one of the radiation products of glycine in solution, have also been studied [63].
Interestingly, glycine and methylamine have been produced experimentally by synthesis in gas-phase reactions in UV-irradiated interstellar ice analogs and in Miller-Urey-type experiments (potential abiotic syntheses in various environments). Glycine synthesis from adding meteoric CO2 or HCN to methylamine can occur by photochemical processes. It has been proposed that glycine and methylamine can be formed from the hydrogenation of CO to formaldehyde and the subsequent Strecker reaction and nucleophilic substitution, as well as from the molecular photodissociation of acetic acid and hydroxylamine to give highly reactive radical/ionic species [60]. The geometry and vibrational frequencies of neutral and protonated glycine have been investigated, finding that the preferred protonation site is the amino group [64]. Studies have shown that amino acids, primarily glycine, and serine, comprise a significant fraction of the total organic material in the atmosphere and can serve as cloud condensation nuclei by forming water clusters [65]. Glycine is related to biological membranes. It causes morphological changes even in the outer layer of bacteria. Adding 1.0% glycine to the culture medium was shown to facilitate the formation of membrane vesicles in a non-pathogenic probiotic strain of Escherichia coli and to alter the protein profile by increasing the inner membrane and cytoplasmic proteins [66].
In this aspect, the reaction between Ca2+ ions, kaolinite Al2Si2O5(OH)4, and glycine C2H5NO2. The reactions are as follows:
1. A cluster is formed with a first layer of 6 water molecules. A second layer of up to 24 water molecules can also be formed.
2. Adsorption of glycine on kaolinite. Glycine interacts with the hydroxyl groups of kaolinite and clusters around the calcium ion in principle:
Al2SiO5(OH)4 + 2NH2CH2COOH + [Ca(H2O)6]2+→Al2SiO5(OH)4(NH2CH2COOH)Ca2+ + 6H2O (3)
3. Condensation and polymerization of glycine with principle reaction:
2NH2CH2COOH t, Ca2+, kaolinite → NH2CH2CONHCH2COOH (4)
Glycine (NH2CH2COOH) is adsorbed on the kaolinite surface (Al2Si2O5(OH)4) as the carboxyl groups of glycine interact with the hydroxyl groups of kaolinite. Calcium ions from cluster structures [Ca(H2O)6]2+ contribute to stabilizing adsorbed molecules by coordinating with glycine carboxyl groups and water molecules in the hydration cluster in the principal reaction.
The polymerization of amino acids on mineral surfaces such as kaolinite can lead to the formation of peptide bonds. The polymerization of glycine on the surface of kaolinite requires specific conditions, including high temperature or the presence of catalysts such as transition metals. The pH of the medium is also essential here. According to Calvin, every condensation-dehydration reaction of amino acids and nucleotides in individual blocks of peptides and nucleic acids occurred in the primary hydrosphere at pH=9-11 [67].
Protostructures under corona discharge conditions
We experimented with a coronal electric discharge with water from Rupite, Bulgaria. The water is placed in a dielectric container and is completely isolated from the air. The discharge parameters are 1–30 kHz, 12 kV [39].
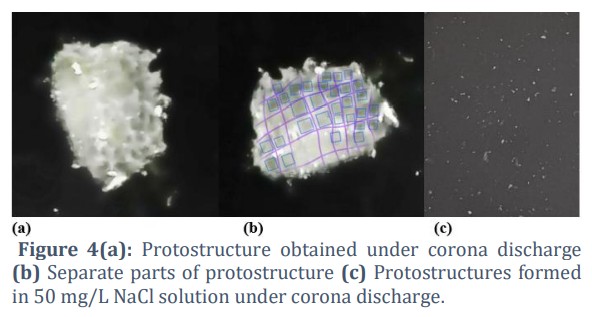
In Fig. 4 a, we show a protostructure resembling we obtained experimentally, with a size of 14.2/12.1/12.4 mm and a volume of 1100 mm3 =1.1 cm3. The corona discharge electrode was filled with water from Rupite, Bulgaria. The impact experiments were performed with a corona electric discharge with parameters specified in [36, 37]. Small structures with a volume of 3.8 mm3 were formed in the beginning. They are documented and marked in Fig. 4 b.
At the beginning of the corona discharge experiment, small protostructures form due to the interaction of electrons and ions with water molecules, containing inorganic and organic compounds, with a volume of 3.8 mm³. These small structures are stable under initial conditions and are organized locally through electrostatic and capillary interactions. Through self-organization and fusion, they gradually combine to form larger proto-structures, reducing a system’s free energy. Favorable conditions, such as high temperature, the presence of ions, and the electric dynamics of the corona discharge, contribute to this process. A larger structure with a volume of 1,100 mm³ is formed. This demonstrates a mechanism of self-organization characteristics of the early stages of biochemical evolution.
The results of corona electric discharge experiments with protostructure formation were repeated under the same conditions. Under these conditions, we also examined a sample with a 50 mg/L NaCl solution in deionized water. Protostructures were formed in the presence of an electric discharge. The structures that were obtained are shown in Figure 4c. No structures were observed in the control sample of the same solution without applying electric discharge. The size of the structures is 1.1/0.8/0.9 mm, and the volume is 0.4 mm3. This structure is 9.5 times smaller than small protostructures with a volume of 3.8 mm3. There was no creation of a big protostructure of 1100 mm3.
The protostructure, formed through corona discharge in mineral water, contains 77% of the ions, and the remaining 23% is present in the water surrounding the electrode. It has a density of 0.479 g/cm3. This is approximately 45.6% of the density of living cells, estimated at around 1.1 g/cm3.
This difference reflects the lower concentration of substances and less organization of the protostructure compared to a living cell. Living cells contain high concentrations of organic and inorganic molecules, including proteins, lipids, and minerals, contributing to their greater compactness and complexity.
In contrast, the protostructure has a higher water content and lacks the intricate molecular organization characteristic of living cells. This comparison highlights that while the protostructure exhibits self-organization and stability, it represents an early stage of biochemical evolution. The analysis supports the idea that the primordial atmosphere and hydrosphere with electric discharge and hot mineral water may be critical in forming fundamental biological structures.
The physicochemical composition of the structure from Fig. 4
The following results are valid for the physicochemical composition:
- The physicochemical composition of the water in the electrode with volume 115 cm3 is: Ca2+ at 3.8; Na+ at 61.1; Mg2+ at 1.8; HCO3− at 176.3; SO42- at 9.3; HS– at 4.8 in mg in 115 cm3
- The physicochemical composition of the water in the protostructure with volume 1.1 cm3 is: Ca2+ at 0.03; Na+ at 0.45; Mg2+ at 0.01; HCO3− at 1.3; SO42- at 0.07; HS– at 0.04mg in 1.1 cm3
- These ions have the properties of ionic centers around which water molecules arrange themselves. Metal ions are hydrophilic because they can form ionic or polar bonds with water. Hydrocarbon structures are hydrophobic because of the hydrocarbon chains. Air is a dielectric, and the corona gas discharge occurs in it. Water is a conductor, and the discharge mainly flows along the surface.
The smallest structure under these conditions was evaluated, with a volume 3.8 mm3. In Fig, 4 b the smaller structures appear darker than the surrounding areas. This means a more significant component concentration exists to preserve the structure formed.
Self-organization in the artificial protostructure is a process in which interacting components, such as ions in solution, spontaneously form organized and stable shapes. These structures evolve through interactions that minimize the system's free energy while enhancing structural complexity and organization. Electrons are released during the electrical corona discharge, contributing to the formation of the structures.
Membrane formation
The self-organization of lipid molecules in aqueous media leads to the formation of bilayers that encapsulate organic molecules. Lipids are a heterogeneous group of substances that are poorly soluble in water but soluble in organic solvents [2]. Animal lipids contain fatty and monocarboxylic organic acids with a linear carbon chain of 4-26 C atoms. They can be saturated, unsaturated, and mono- or polyethylene [2]. Characteristic properties are their low solubility in water, their acidic nature, which decreases with increasing carbon atoms in the chain, and their reactivity with alcohols (they form esters). Unsaturated fatty acids can easily include halogens, such as iodine. Lipids are classified as simple lipids (acyl glycerols – esters of glycerol, fatty acids, and waxes) and complex ones (phospholipids, glycolipids, and lipoproteins). Lipids form the basic structure of all biological membranes [2].
Lipids are essential for the existence and functioning of living cells. They form the basis of cell membranes, which consist of polar lipids composed of a hydrophobic and a hydrophilic part. Lipids dissolve readily in organic solvents but have a region that is attracted to and soluble in water. This "amphiphilic" property is fundamental to their role as building blocks of cell membranes. Their molecules have a head (often of glycerol) attached to two long fatty acid chains (tails). These tails repel water and dissolve readily in organic solvents, giving the molecule its lipid character. In water, under laboratory conditions, they spontaneously form spherical structures called liposomes, which have a double-layered wall. The water-soluble heads of the lipids face outward in water. The two layers form a fluid, sandwich-like structure, with the fatty acid chains in the middle due to their hydrophobicity. This type of lipid bilayer, formed by the self-assembly of lipid molecules, is the basic structure of the cell membrane. It is the most stable thermodynamic structure [44]. Possibilities for spontaneous membrane formation are described due to the property of the hydrophobic parts of lipids to self-associate in an aqueous environment and the tendency of their hydrophilic parts to interact with the aqueous environment and with each other [68]. This has allowed the primary cells to separate their interior from the outside environment and form the organelles within the cell. Microbodies are roughly spherical, 0.5 to 1 μm in diameter, surrounded by a single membrane [44]. Membrane organelles allow the separation of specific chemical reactions and the retention of their products within the cell. The hydrophobic film formed by the lipid bilayer (lipid asymmetry) limits the cell from the environment and forms it as an independent biological unit. Additionally, the biophysical properties of various lipids and their phase behavior significantly contribute to membrane function. In addition to being major structural components of membranes, lipids serve other functions, including acting as energy storage molecules [68, 69].
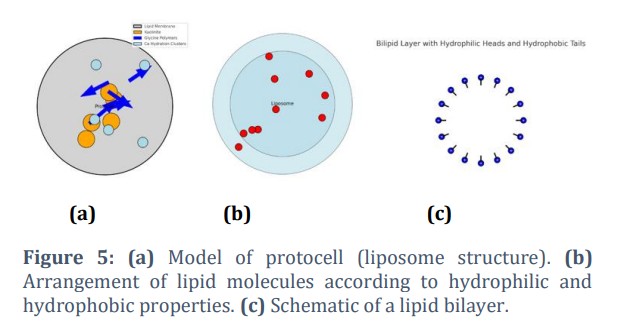
Fig. 5a illustrates a liposome, a structure formed by lipid bilayers encapsulated in water and other molecules. The membrane consists of phospholipids that self-assemble in an aqueous environment.
Various biomolecules, such as amino acids, nucleotides, small peptides, etc., can reside inside the liposome. The figure shows them represented by red circles.
Liposomes are essential structures in prebiotic biochemistry because they enable the isolation of chemical reactions in a confined space, a process crucial for the development of complex biochemical systems [66]. Liposomes can exhibit protocell-like behavior by encapsulating organic molecules and facilitating interactions. As a result, more complex life forms are likely to have evolved.
The cell membrane is a liquid organization of amphiphilic molecules, a hundred times more viscous than water, similar to thin oil. This chemical structure makes it exceptionally flexible, the ideal boundary for rapidly growing and dividing cells. It is also a reliable barrier, allowing some solutes to pass while blocking others. The lipid bilayer gives the membrane its characteristic permeability (the rate at which a substance in solution passes through it). Lipid-soluble compounds and some small molecules can permeate the membrane. Still, the lipid bilayer effectively repels the many large, water-soluble molecules and electrically charged ions the cell must import or export to live [41].
Today's smallest cell is 150-200 nm or 0.15-0.2 µm, comparable to the size of viruses, among the simplest viable biological objects. Viroids (consisting only of RNA) and prions, representing an infectious protein, are more straightforward than these [7]. The probability that they were the first to form on Earth is high, but the possibility is no less that these were the extremophile archaebacteria.
Fig. 5b shows an arrangement of lipid molecules depending on their parts' hydrophilic and hydrophobic properties. The basic structure of all biological membranes is the lipid bilayer, but in the cells, it contains phospholipids, glycolipids, and cholesterol. They form a liquid phase to which proteins bind. Carbohydrates are often attached to membrane lipids and proteins [3]. Fig. 5c schematically represents a lipid bilayer. The blue circles with (+) and (-) charges represent the polar hydrophilic heads of the lipid molecules. The symbols illustrate polarity, not necessarily to presence electric charge. They interact with water molecules. The lines represent nonpolar hydrophobic tails that avoid water molecules and line up against each other. The specific structure of the lipoprotein membranes of cells is due to this arrangement of lipid molecules.
Biological membranes enclose the cell's interior from the environment [68]. The various membranes performing these vital roles comprise roughly equal weight percentages of proteins and lipids, and carbohydrates are less than 10%. All lipids are amphipathic, with hydrophilic (polar) and hydrophobic (nonpolar) parts. As a result, the lipid components of the membranes are arranged in a continuous bimolecular bilayer less than 10 nm thick. The polar parts of its constituent molecules lie on both surfaces of the double layer, while the non-polar parts make up its interior. Thus, this structure serves as an impenetrable barrier to major water-soluble substances in the cell, providing the basis for the separation function of biological membranes [70]. Although the surfaces of the membranes contain polar units, they act as electrical insulators and can withstand several hundred thousand volts without failure. It has been found experimentally and theoretically that the structure and these unusual properties are imparted to biological membranes by the lipid bilayer [70].
Nucleic acids are other vital biopolymers. Polymerization can link nucleotides into polymers (e.g., RNA molecules). However, these processes in natural conditions require catalysts and suitable energy sources, such as UV radiation or chemical energy inputs. In laboratory experiments, such processes are facilitated by varying conditions, including pH and temperature, in the presence of ions.
The process is activated by catalysts or on surfaces, which improves the formation of bonds between nucleotides. Minerals such as kaolinite, clay, and pyrite serve as catalysts. They facilitate the linking of nucleotides into longer polymers. Water clusters with metal ions such as Mg2+ and Ca2+ can stabilize the RNA structure by neutralizing the negative charge on the phosphate chain so the molecule folds appropriately.
As a result of the reactions of calcium and magnesium, [71] water produces molecular hydrogen:
Magnesium ions stabilize RNA molecules, and calcium ions stabilize the phospholipids of cell membranes.
Transport of compounds through cell membrane
The cell membrane is a barrier with selective permeability. The transport across membranes occurs by various mechanisms: simple diffusion (driven by concentration or electrical gradients of the transported solute), passive transport or facilitated diffusion (also follows concentration or electrical gradients but mediated by carriers and channels), active transport (against the electrochemical gradient of substances, with energy expenditure), through secondary active transporters (use the electrochemical gradient created by primary active transport systems such as the sodium pump), endocytosis (phagocytosis and pinocytosis) and exocytosis [3].
The movement of dissolved substances through the cell membrane occurs according to the principle of diffusion. According to this principle, the solute diffuses along the concentration gradient, that is, from a place of higher to a place of lower concentration, until equal concentration is reached at both places and equal, random diffusion in both directions. A solute with a high concentration has a high free energy; that is, it is able to do more diffusion than a solute with a low concentration. In diffusion, the solute loses free energy, so that when it reaches equilibrium at a lower concentration, it is unable to return spontaneously (under the action of its own energy) to its previous high concentration [44]. Water and alcohols penetrate most membranes in less than a second. Water is insoluble in organic solvents, but penetrates cell membranes due to the small size of its molecules. The ability of water to easily pass through membranes is particularly important for establishing equilibrium [44].
Interestingly, molecules in biological membranes are constantly in motion. In fact, the membrane can be considered a two-dimensional fluid in which the protein components move like boats. The arrangement of the lipid molecules in the bilayer, with their polar ends toward the surface and the nonpolar parts toward the central region, gives this bilayer structure the molecular orientation of a crystal and the fluidity of a liquid. In this liquid crystalline state, thermal energy causes lipid and protein molecules to diffuse laterally and rotate about an axis perpendicular to the plane of the membrane. In addition, lipids occasionally flip from one surface of the bilayer membrane to the other and attach and detach from the surface of the bilayer at very slow but measurable rates. Lipids and proteins can exhibit restricted rocking motions. The membrane is a "fluid mosaic" of complex lipids and proteins molecular associations. In this way, the plane of the biological membrane is divided into parts by domain structures, similar to how the three-dimensional space of the cell is divided into parts by the membranes [71]. Some authors suppose extraterrestrial life is connected with proteins and a nucleus [72].
Through the membrane of the protocell, there should be a movement of water molecules, ions, and salts from a water lake located on both sides of it. To explain this process, let's apply the Nernst equation for an osmotic process:
Е=RT/zFln |C|out/|C|in
E is the equilibrium potential (V)
R is a universal gas constant (8.314 J.mol-1.K-1)
T is the absolute temperature (K)
z is the valency of the ion
F e Faraday constant (96485).
At the amount of calcium ions in water 0.01 mol/L or 400.8 mg/L, the amount of calcium ions in the protostructure is assumed to be 0.1 mol/L or 4008 mg/L.
According to Nernst's law, at 20 °C, the electric potential on both sides of the membrane is (-29 mV). At 75 °C the result is (-35 mV):
E75 °C = 8.314×348.15/2x96485xln(0.01/0.1)=(-34.5 mV).
In osmosis, water enters the protocell, but the electrical parameters are kept constant on both it sides. This means diffusion will begin from the protocell to the lake water to maintain the electrical potential.
![]() Discussion
Discussion
The origin of life on Earth remains one of the greatest scientific mysteries. The Earth, 4.54 billion years old, is situated in the Sun's habitable zone. Remarkably, the first evidence dates back 3.5 billion years ago, highlighting the favorable conditions of our planet. A team of Australian scientists has studied the Pilbara region of Western Australia [73, 74]. Well-preserved traces are found in ancient marine, lakeshore, and hot springs. Stromatolites primarily consist of calcium carbonate (CaCO3) [75]. Damer and Deamer propose a scenario for the origin of life in fluctuating volcanic hot spring pools [22]. In 2010, Ignatov and Mosin explored the possibility of life arising in hot mineral water lakes [10, 59]. The spectral analyses indicate that the model systems are similar to mineral water rich in calcium and bicarbonate ions. Szostak emphasizes that open mineral lakes provide more life opportunities due to the accumulation of essential minerals [19, 20]. In this paper, the authors explore the structuring of clusters around metal ions, primarily calcium, as a precursor to the formation of living cells. Possible reactions in the reducing atmosphere of ancient Earth are described. The primordial atmosphere, devoid of oxygen and composed mainly of methane, ammonia, and carbon dioxide, vastly differed from the current one. Biochemical processes are discussed for structuring glycine and other amino acids in forming living cells [76]. The uniqueness of glycine is highlighted by its identification in meteorite samples [77]. During the ancient Earth’s history, high-voltage electric discharges in the primordial atmosphere were prevalent. In 1953, Miller and Urey synthesized organic compounds under electric discharge conditions [8]. Numerous scientists replicated their experiments [9]. In 2013, Ignatov and Mosin demonstrated with experiments that complex organic molecules can arise spontaneously under extreme conditions in the primordial atmosphere and hydrosphere in Miller’s and Wilson’s experiments. The linear relationship between the mass of the sediment and the number of carbon atoms in the molecule is presented in Miller’s and Wilson’s experiments. The results were observed using entropy analyses primarily applied by Prigogine self-organization. According to Prigogine’s principles, systems transition from chaos to order, and the accumulation is complex. The decrease in entropy observed in the experiments indicates that the systems become increasingly organized, enabling self-organization and the evolution of more complicated structures. The calculated Shannon entropy demonstrates moderate compound diversity, balancing complexity and order. One of the co-authors, Ignatov, created an artificial structure in laboratory conditions using corona discharge at the air-water boundary. The air had a chemical composition resembling the ancient atmosphere, while the hot water, sourced from Rupite, Bulgaria, was rich in calcium and hydrogen carbonate ions. The resulting artificial structure comprised 45.6 % of the density of a living cell, with its chemical composition thoroughly analyzed. Adamatzky has also explored the possibility of creating a cell under laboratory conditions through computer modeling [33,34]. Osmotic processes between the cell membrane and the surrounding environment are described using the Nernst equations. The energy sources in water for the origin of living cells are chemical, solar, and geothermal. Geothermal energy is constant and closely associated with hot mineral water in springs, ponds, and lakes. The discovery of the thermophilic bacterium Anoxybacillus rupiensis sp. Nov. in Rupite by the scientists further supports the uniqueness of the evolutionary environment. The physicochemical composition of the tested mineral water from Rupite, Bulgaria, was studied. The main chemical reactions involved in the formation of biopolymers in hot mineral water, which played a crucial role in the biogenesis of the ancient Earth, are presented. In the water from Rupite, the Fourier Transform Infrared test showed inorganic and organic compounds. In an experiment with a coronal electric discharge using water from Rupite, Bulgaria, in a dielectric container completely isolated from the air at discharge parameters of 1-30 kHz and 12 kV, we observed the formation of an artificial protostructure with a volume of 1100 mm³. The protostructures formed under corona electric discharge in 50 mg/L NaCl solution with a volume 0.4 mm3, making them 9.5 times smaller than previously observed small protostructres of inorganic and organic compounds (3.8 mm3) and significantly smaller 2750 times than larger protostructure (1100 mm3). The protostructure in the mineral water from Rupite had a volume of 1.1 cm3. About 45.6% of the density of the living cell was structured in the protostructure. This density was 0.479 g/cm3. The analysis reveals that the conditions for the origin of life in waters with an ion content exceeding 2250 mg/L are optimal for this natural phenomenon. In conditions involving corona discharge and hot mineral water, an environment of energy density and dynamic processes is created, facilitating a localized entropy. The electrons and ions released during the corona discharge interact with the water's surface, forming structures. These processes enhance the self-organization and polymerization of organic molecules by increasing local concentration and energy. Hot mineral water, enriched with Ca2+, Mg2+, and HCО3- ions, stabilizes water clusters and provides a suitable matrix for chemical reactions. The temperature gradient and reducing environment further promote the formation of more complex molecules for self-organization and structural complexity – key steps toward the origin of life. These findings highlight the importance of energy input via corona discharge and the properties of hot mineral water as critical factors for overcoming chaos and fostering the formation of ordered biological structures in the primordial hydrosphere.
Tables and Figures
Author Contributions
I.I. conducted the tests, physical, biophysical, and chemical analyses, and prepared the figures. T.P.P. performed the biochemical and cell analyses and wrote the text. The article is in memory of Oleg Mosin (1966-2016).
Conflict of Interest
The authors declare that there is no conflict of interest.
![]() References
References
- Blanco A, Blanco G. Chapter 1 – Chemical composition of living beings. In: Medical Biochemistry. Editor(s): Antonio Blanco, Gustavo Blanco, Academic Press, (2017): 1-3.
- Blanco A, Blanco G. Chapter 5 – Lipids. In: Medical Biochemistry. Editor(s): Antonio Blanco, Gustavo Blanco, Academic Press, (2017): 99–119.
- Blanco A, Blanco G. Chapter 11 – Membranes. In: Medical Biochemistry. Editor(s): Antonio Blanco, Gustavo Blanco, Academic Press, (2017): 215–250.
- Cogley JG, Henderson-Sellers A. The origin and earliest state of the Earth's hydrosphere. Reviews Geophysics, (1984); 22(2):131–175.
- Shimazu Y, Urabe T. Some numerical experiments on the evolution of the terrestrial atmosphere and hydrosphere. Journal of Physics of the Earth, (1967);15:1–18.
- Hazen RM, Papineau D, Bleeker D, et al. Mineral evolution, American minerals, (2008); 93: 1693–1729.
- Popova TP. Microbiology. Academic Press of the University of Forestry, Bulgaria, (2016); 1–311 [in Bulgaria].
- Miller S, Urey HC. Organic compounds synthesis on the primitive Earth: Several questions about the origin of life have been answered, but much remains to be studied. Science to be studied. Science, (1959);130: 245–251.
- Bada JL, Lazcano A. Prebiotics soup – revisiting the Miller experiment. Science, (2003); 300: 745–746.
- Colon Santos SM. Exploring the untargeted synthesis of prebiotically-plausible molecules. PhD thesis. Glasgow University (2019).
- Sasselov D, Grotzinger JP, Sutherland JD. The origin of life as a planetary phenomenon. Science Advances, (2020);6(6).
- Longo MG, Vialetto L, Diomede P, Longo S, Laporta V. Plasma modeling and prebiotic chemistry: A review of the state-of-the-art and perspectives. Molecules, (2021);26(12):3663.
- Gan DW, Hong LF, Li TY, Zhang TQ, Wang XY et al. Discharge plasma for prebiotic chemistry: Pathways to life’s building blocks. Earth and Planetary Physics, (2024); 8(6): 1–13.
- Plankensteiner K, Reiner H, Schranz B, Rode BM. Prebiotic formation of amino acids in a neutral atmosphere by electric discharge. Angewandte Chemie, (2004); 43(14): 1886–1888.
- Ignatov I, Mosin O. Modeling of possible processes for the origin of life and living matter in hot mineral and seawater with deuterium. Journal of Environment and Earth Science, (2013); 3: 103–118.
- Ignatov I. Origin of life and living matter in hot mineral water and properties of polar molecules in the primary hydrosphere and hydrothermal ponds. Uttar Pradesh Journal of Zoology, (2021); 42: 37–52.
- Derekova A, Sjøholm C, Mandeva R, Kambourova M. Anoxybacillus rupiensis sp. Nov., isolated from Rupi basin (Bulgaria). Extremophiles, (2007); 11: 577–583.
- Mougkogiannis P, Adamatzky A. Proto-neurons from abiotic polypeptides. Encyclopedia, (2024); 4: 512–543.
- Szostak JW. An optimal degree of physical and chemical heterogeneity for the origin of life? Philosophical Transactions of the Royal Society, (2011); 366: 2894–901.
- Szostak JW. On the origin of life. Medicina (Buenos Aires), (2016); 76.
- Mulkidjanian AY, Galperin MY. On the origin of life in the Zinc world. Validation of the hypothesis on the photosynthesizing zinc sulfide as cradles of life on Earth. Biology Direct, (2009); 4: 26–28.
- Damer B, Deamer D. Coupled phases and combinatorial selection on fluctuating hydrothermal pools: A scenario to guide experimental approaches to the origin of cellular life. Life, (2015); 5(1): 872–887.
- Kochanski E, Constantin E. Theoretical and experimental studies of the system Ca+(H2O)n for n=1 to 10. The Journal of Chemical Physics, (1987); 87: 1661–65.
- Watanabe H, Iwata S. Molecular orbital studies of the structures and reactions of a singly charged calcium ion with water clusters, Ca+(H2O)n. The Journal of Physical Chemistry A, (1997); 101: 487–496.
- Ariyarathna IR, Miliordos E. Geometric and electronic structure analysis of calcium water complexes with one and two solvation shells. Physical Chemistry Chemical Physics (2020); 39.
- Delgado AA, Sethio D, Kraka E. Assessing the intrinsic strengths of ion–and solvent–solvent interactions for hydrated Mg2+ Clusters, Inorganics, (2021);9(5): 31.
- Wang P, Su Y, Shi R, Huang X, Zhao J. Structures and spectroscopic properties of hydrated Zinc(II) Ion Clusters [Zn2+(H2O)n (n = 1−8)] by Ab Initio Study. Journal of Cluster Science, (2023); 34: 1625–1632.
- Pathak AK, Mukherjee T, Maity DK. IR Spectra of carbonate‐water clusters, CO3−2(H2O)n: A theoretical study. Synthesis and reactivity in inorganic, metal-organic, and nano-metal chemistry, Synthesis and Reactivity in Inorganic, Metal-Organic, and Nano-Metal Chemistry, (2008); 38(1): 76–83.
- Custelcean R, Williams NJ, Seipp ChA. Aqueous sulfate separation by crystallization of sulfate–water clusters. Angewandte Chemie, (2015); 54(36): 10525–10529.
- Rudolph WW, Irmer G. Raman and infrared spectroscopic investigations on aqueous alkali metal phosphate solutions and Density Functional Theory calculations of phosphate–water clusters. Applied Spectroscopy, (2007); 61(12): 1312–1327.
- Ignatov I, Marinov Y, Huether F, Gluhchev G, Iliev MT. Modeling water clusters: Spectral analyses, Gaussian distribution, and linear function during time. Ukrainian Journal of Physics, (2024); 69(9): 632–642.
- Vassilev N, Ignatov I, Popova TP, Huether F, Ignatov AI et al. Nuclear Magnetic Resonance (NMR) and Density Functional Theory (DFT) study of water clusters of hydrogen-rich water (HRW). Water, (2024); 16(22): 3261.
- Sharma S, Mougoyannis P, Tarabella G, Adamatzky AA. Review on the protocols for the synthesis of proteinoids arXiv: (2022); 2212.02261.
- Adamatzky A. Towards proteinoid computers. Hypothesis paper. Biosystems, (2021); 208: 104480.
- Harada K, Fox SW. The thermal condensation of glutamic acid and glycine to linear peptides. Journal of American Chemical Society, (1958); 80: 2694–2697.
- Brack A. Liquid water and the origin of life. Origin of Life and Evolution of Biosphere, (1993); 23: 3–10.
- Kurihara K, Tamura M, Shohda K-I, Toyota T, Suzuki K, Sugawara T. Self-reproduction of supramolecular giant vesicles combined with the amplification of encapsulated DNA, Nature Chemistry, (2011); 3 (10): 775–781.
- Sato Y, Takinoue M. Pioneering artificial cell-like structures with DNA nanotechnology-based liquid-liquid phase separation. Biophys. Physicobiology, (2024); 21: e210010.
- Ignatov I, Drossinakis Ch, Ignatov AI. Color coronal spectral analysis: Results with water solution of Calcium Carbonate. Portugaliae Electrochimica Acta, (2025); 43(2): 113–126.
- Nevoit G, Bumblyte IA, Korpan A, Bumblyte IA, Potyazhenko M et al. The biophoton emission in biotechnological and chemical research: From meta-epistemology and meaning to experiment. Part 1. Ukrainian Journal of Physics (2024); 69(3): 190–206.
- Mehandjiev D, Neshev N, Vassileva P, Vassileva P, Gluhchev G et al. History-dependent hydrogen bonds energy distributions in NaCl aqueous solutions undergoing osmosis and diffusion through a ceramic barrier. Journal of Chemical Technology and Metallurgy, (2023); 58(2): 340–346.
- Ignatov I, Gluhchev G, Ignatov AI. Desalination of seawater. Osmotic process for “blue energy” and estimation for desalination, Ukrainian Journal of Physics, (2024); 69(12): 905–913.
- Klein F, Tarnas JD, Bach W. Abiotic sources of molecular Hydrogen on Earth. Elements, (2020); 16(1): 19–24.
- Schmidt-Rohr K. Oxygen is the high-energy molecule powering complex multicellular life: Fundamental corrections to traditional bioenergetics. ACS Omega (2020); 5(5): 2221–2233
- Miliordos E, Xantheas SS. Ground and excited states of the [Fe(H2O)6]2+ and [Fe(H2O)6]3+ Clusters: Insight into the electronic structure of the [Fe(H2O)6]2+–[Fe(H2O)6]3+ Complex. J. Chem. Theory Comput.(2015); 11(4): 1549–1563.
- Ignatov I. Review of different types of mountain springs and mineral waters from Bulgaria based on their natural origin and health benefits. Med. Perspekt, (2023); 51(4): 199–206.
- Picard A, Garcman A, Girquis PR. Interactions between iron sulfide minerals and organic carbon: Implications for biosignature preservation and detection. Astrobiology, (2021); 21(5).
- Jougnot D, Roubinet D, Guarracino L, Maineult A. Modeling streaming potential in porous and fractured media, Description and benefits of the effective excess charge density approach. In: Biswas, A., Sharma, S, (eds) Advances in Modeling and Interpretation in Near Surface Geophysics. Springer Geophysics. Springer, Cham, (2020).
- Ignatov I, Mosin OV. S. Miller’s experiments in modelling non-equilibrium conditions with gas electric discharge simulating primary atmosphere. European Journal of Molecular Biotechnology, (2015); 10(4): 197-209.
- Wilson AT. Synthesis of macromolecules. Nature, 1960; 188: 1007–1009
- Trachman M, Markham GD, Glusker J P et al. Interactions of metal ions with water: Ab initio molecular orbital studies of structure, vibrational frequencies, charge distributions, bonding enthalpies, and deprotonation enthalpies. 2. Monohydroxides, Inorganic Chemistry, (2001); 40 (17): 4230–4241.
- Miliordis E, Xantheas SS. Ground and excited states of the [Fe(H2O)6]2+ and [Fe(H2O)6]3+. Clusters: Insight into the electronic structure of the [Fe(H2O)6]2+– [Fe(H2O)6]3+ complex. Journal of Chemical Theory and Computation, 2015; 11(4): 1549–1563.
- Wang P, Shi R, Su Y, et al. Hydrated sodium ion clusters [Na+(H2O)n (n = 1–6)]: An ab initio study on structures and non-covalent interaction, (2019); 7.
- Garand E, Wende T, Goebbert D J et al. Infrared spectroscopy of hydrated bicarbonate anion clusters: HCO3−(H2O)1−10 J. Am. Chem. Soc, (2010); 132(2): 849–856.
- Pletnev IV, Smirnova SV, Sharov AV, Zolotov YA. New generation extraction solvents: from ionic liquids and aqueous biphasic systems to deep eutectic solvents. Russian Chemical Reviews, (2021);90(9):1109.
- Erfurt P. The geochemistry of hot Springs. In: The geoheritage of hot Springs. Geoheritage, geoparks and geotourism. Springer, Cham, (2021)51-90.
- Bychowski J. Understanding how ORP is used in cooling water treatment. Cmem-aqua, (2022).
- Kamenshchikov SA. External Prigogine theorem method for universal characterization of complex system evolution. Chaos and Complexity Letters, (2014); 8(1): 63–73.
- Ignatov I. Entropy and time in living organisms. ArchivEuromedica. 1st&2nd Edition, (2011); 74-75.
- Aponte JC, Jamie EE, Daniel PG, Milam SN, Charnley SB et al. Pathways to meteoritic glycine and methylamine. ACS Earth Space Chemistry, (2017);1(1): 3−13.
- Razak M A, Begum P S., Viswanath. B., Rajagopal, S. Multifarious beneficial effect of nonessential amino acid, glycine: A review. Oxidative Medicine and Cellular Longevity, (2022); 9857645
- Qiao W, Qiao Y. The relationship between the structure and properties of amino acid surfactants based on glycine and serine. Journal of Surfactants and Detergents, (2013); 16: 821–828.
- Simon S, Gil A, Sodupe M, Bertrán J. Structure and fragmentation of glycine, alanine, serine and cysteine radical cations. A theoretical study. Journal of Molecular Structure: THEOCHEM. (2005); 727 (1-3): 191–197.
- Noguera M, Rodrı́guez-Santiago L, Sodupe M, Bertran J. Protonation of glycine, serine and cysteine. Conformations, proton affinities and intrinsic basicities. Journal of Molecular Structure: THEOCHEM, (2001); 537: 307–318.
- Ball BT, Vanovac S, Tuguldur T, Shields GC. Monomers of glycine and serine have a limited ability to hydrate in the atmosphere. The Journal of Physical Chemistry A, (2021); 125 (38): 8454–8467.
- Hirayama S, Nakao R. Glycine significantly enhances bacterial membrane vesicle production: a powerful approach for isolation of LPS-reduced membrane vesicles of probiotic Escherichia coli. Microbial Biotechnology, (2020); 13(4): 1162–1178.
- Calvin M. Chemical Evolution, Oxford: Clarendon, (1969);1- 278
- Van Meer GDR, Voelker GW. Feigenson. membrane lipids: where they are and how they behave. Nature Review Molecular Cell Biology, (2008); 9: 112–124.
- Luisi PL, Walde P, Oberholzer Th. Lipid vesicles as possible intermediates in the origin of life. Interface Science, (1999); 4: 33–39.
- Thompson TE. Lipids in biological membranes. Encyclopedia Britannica, (2024)
- Toshkova R, Neshev N, Ignatov I, Huether F, Ignatov AI, Angushev I. Effects of hydrogen-rich water on hamsters with experimental myeloid tumor. Libri Oncologici, (2023);51: 85–96.
- Tagami Sh. Why we are made of proteins and nucleic acids: Structural biology views on extraterrestrial life. Biophys. Physicobiology, (2023); 20: e200026
- Van Kranendonk MJ, Deamer DW, Djokic T. Life springs. Scientific American, (2017); 317(2): 28–35.
- Baumgartner RJ, Van Kranendonk MJ, Fiorentini ML, Pages A, Wacey D et al. Formation of micro-spherulitic barite in association with organic matter within sulfidized stromatolites of the 3.48 billion-year-old Dresser Formation, Pilbara Craton. Gebiology, (2020); 18(4): 415–425.
- Kaźmierczak J, Fenchel T, Kühl M, Kempe S, Kremer B et al. CaCO3 Precipitation in multilayered cyanobacterial mats: Clues to explain the alternation of micrite and sparite layers in calcareous stromatolites. Life, (2015); 5 (1): 744–769.
- Emeline AV, Otroshchenko VA, Ryabchuk VK, Serpone N. Abiogenesis and photostimulated heterogeneous reactions in the interstellar medium and on primitive earth: Relevance to the genesis of life. Journal of Photochemistry and Photobiology C: Photochemistry Reviews, 2003; 3(3): 203–224.
- Aponte JC, Elsila JE, Glovin DP, Milam SM, Charnley S et al. Pathways to meteoritic glycine and methylamine. ACS Earth and Space Chemistry, (2017); 1(1): 3–13.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0