

Review Article
Optimizing Blood-Stage Vaccines: Essential Tools in Malaria Eradication Strategies
Musa Ahmad Aminu1, Aneeq Ur Rehman1, Atif Amin Baig2, Ahmed M. Salman3, Ahmad Syibli Othman1,3*
Adv. life sci., vol. 12, no. 2, pp. 425-434, May 2025
*– Corresponding Author: Ahmad Syibli Othman (Email: syibliothman@unisza.edu.my)
Authors' Affiliations 1. Faculty of Health Sciences, Universiti Sultan Zainal Abidin, 21300 Kuala Nerus – Malaysia
2. International Medical School, Management and Science University, 40100 Shah Alam – Malaysia
3. The Jenner Institute, University of Oxford, Oxford – United Kingdom
2. International Medical School, Management and Science University, 40100 Shah Alam – Malaysia
3. The Jenner Institute, University of Oxford, Oxford – United Kingdom
[Date Received: 20/10/2024; Date Revised: 05/01/2025; Date Available Online: 31/08/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Malaria continues to be a major threat to global health, taking a significant toll on mortality rates. Although numerous strategies for combating malaria are actively evolving, none has successfully eradicated the disease. Vaccines against malaria present a reliable approach; licensed liver-stage vaccines have opened a new era in malaria control but because they are not fully effective, a complementary blood-stage vaccine is needed. This review explores the pivotal role of blood-stage vaccines as formidable weapons in the ongoing battle against malaria by elucidating their profound impact on malaria-linked morbidity and mortality. The immunological mechanisms through which blood-stage vaccines mediate significant reductions in disease burden and interrupt transmission dynamics are crucial in vaccine design. Thus, they have been examined in this review. In the context of blood-stage malaria, specific antigens are critical to the survival and replication of the parasite, playing essential roles in the invasion of erythrocytes. As a result, these antigens present highly promising vaccine candidates that can stop the invasion in various ways, such as the binding of sialic acid residues by EBA175 or the hydrolysing effect of MSP1. Polymorphism challenges hinder blood-stage vaccine development but enhancing current candidates and identifying novel erythrocytic antigens offer a promising path to a malaria-free world. This multifaceted approach aims to fortify the immune response against the Plasmodium parasite, thereby advancing global eradication efforts.
Keywords: Malaria; Plasmodium,;Vaccine; Blood-stage
Introduction![]()
Malaria, a parasitic disease caused by an anopheline mosquito, remains a global public health concern. In 2022, an estimated 249 million malaria cases and 608,000 deaths were recorded. The majority of which were in tropical Africa [1]. Five predominant Plasmodium species can cause malaria in humans: Plasmodium malariae, Plasmodium falciparum, Plasmodium vivax, Plasmodium ovale, and Plasmodium knowlesi. Numerous efforts, such as preventative measures and treatment prescriptions, have been continuously developed to combat malaria [2]. However, despite relatively limiting the incidence of the disease, none of these efforts have met the satisfactory level of “malaria eradication” due to the emergence of insecticide-resistant mosquitoes and drug-resistant parasites [3]. Therefore, the development of more effective and sustainable control strategies is imperative.
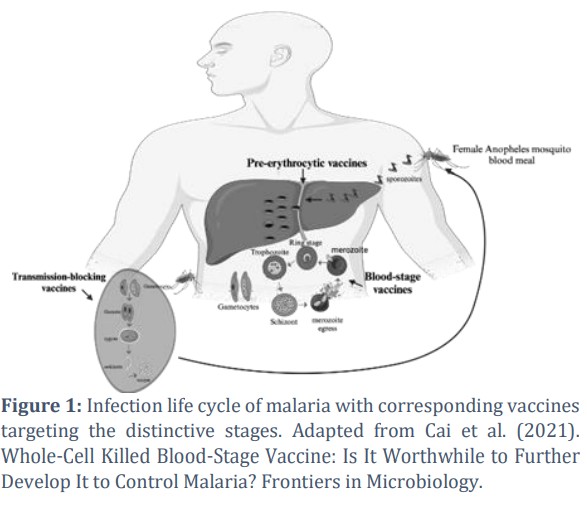
The multi-stage life cycle of malaria allows for a tailored approach in vaccine design, with most research targeting the asymptomatic hepatocytic stage to prevent the disease. Another approach is vaccines targeting the erythrocytes-invading asexual stage to offer protection against clinical symptoms, while others target the mosquito sexual stage to block transmission. Research that aims to produce a placental malaria vaccine to prevent severe sequelae of placental malaria in pregnant mothers and their foetuses is also ongoing.
There is substantial evidence in place that denotes the indispensability of a subunit malaria vaccine [4]. The approval of the RTS, S (Mosquirix) vaccine in 2019 marks the beginning of a promising era. However, Mosquirix only offers protection below the WHO’s objective of at least 75% efficacy. This led to improved efforts in malaria vaccine research, with the recommendation of sporozoites-targeting R21/Matrix- M vaccine at the forefront. The R21/Matrix-M offered up to 80% high-dose adjuvant efficacy in clinical trials [5]. Interestingly, for any pre-erythrocytic vaccine to provide sterile protection on a global scale, it must be 100% effective because even a single surviving sporozoite can lead to an invasion of erythrocytes. Until now, not even the newly licensed R21/Matrix-M meets this demand, elucidating the significance of a complementary blood-stage vaccine.
On the other hand, there is a growing interest in erythrocytic (blood-stage) vaccines to address the issues surrounding pre-erythrocytic interventions. This review discusses the role of blood-stage malaria vaccines (BSVs) in the race to the much-anticipated malaria “elimination” and eventual “eradication” eras. Challenges faced in the development of BSVs and practical ways through which they can be overcome are also critically reviewed.
![]() Methods
Methods
Literature Search Strategy and Selection Criteria
The most relevant and current publications and clinical trial reports on BSVs were included in the review. Research articles and reports of interest were collected from Google Scholar, the Centre for Disease Control and Prevention (CDC), the World Health Organization (WHO), PubMed, Science Direct, and the clinicaltrials.gov databases. Keywords: malaria vaccine, blood-stage malaria vaccine(s), erythrocytic malaria vaccine(s), vaccine efficacy against malaria, therapeutic approach, and Plasmodium (falciparum, vivax, ovale, malariae, knowlesi) were used. Of the 598 papers reviewed, 81 duplicates were excluded. The titles and abstracts of the remaining papers were analysed, and 443 were further excluded because of irrelevance. The remaining 74 and evolving data from relevant websites were included in the study.
![]() Discussion
Discussion
Overview of the Malaria Life Cycle
Although malaria is caused by various Plasmodium spp., the pattern of the infection life cycle is relatively similar. When a malaria-containing female Anopheles mosquito inoculates sporozoites into a primary host, the sporozoites infect liver cells and burst. At this period, mature sporozoites release numerous daughter cells called merozoites into the blood. In P. vivax and P. ovale, dormant hypnozoites can persist in the liver, occasionally causing malaria relapses in the host. Liver-stage vaccines exert their effect by attempting to eliminate the propagation of sporozoites. Without this intervention, sporozoites release merozoites into the bloodstream, which fasten themselves to and invade erythrocytes via asexual reproduction (Figure 1). This process involves three distinct stages with different morphological appearances: the ring stage, the trophozoite, and the schizont. After the schizonts have infected erythrocytes, they rupture and egress, releasing 16 to 32 fresh merozoites into the bloodstream, which then infect new erythrocytes. The asexual erythrocytic invasion repeats every 1 to 3 days, depending on spp. BSVs target this phase to prevent merozoites from invading erythrocytes, significantly reducing disease-linked mortality and morbidity. Without vaccine intervention, the merozoites that infect erythrocytes sporadically proceed to separate sexual states known as microgametocytes and macrogametocytes. The formed gametocytes stay in the blood and are later ingested during the next blood meal. Upon ingestion, the two gametocytes mature into gametes that fuse, forming a zygote in the Anopheles’ gut. The zygote develops and becomes an ookinete that generates an oocyst via movement in the gut. The generated oocyst then releases numerous sporozoites, which travel within the mosquito to reach its salivary glands. Vector-stage vaccines aim to stop malaria transmission by interfering with this sexual stage. The whole process in the Anopheles is completed within 12 to 15 days [6].
Immunology of Blood-Stage Malaria
The versatile nature of the immune system makes it capable of responding against any of the three stages of malaria infection [7]. Naturally acquired immunity against malaria has long been studied in endemic regions and is understood to be short-lived. This can be explained by the waning pattern of malaria-linked high morbidity and mortality in children. The risk of life-threatening episodes declines by early maturity, while the risk of infection remains throughout the lifespan [8]. High antibody titres and T-cell-based responses have been reported in animal models against sporozoites in the liver and schizonts (blood-stage), where they play a significant role in protection [9]. However, in human malaria infection, there is very little evidence regarding the role of liver-stage responses in naturally acquired immunity. Additionally, naturally produced anti-gametocyte antibodies rarely influence malaria transmission in humans living in endemic areas [7]. These two factors pose significant challenges to the liver stage and transmission-blocking interventions [10]. In contrast, there is a clear pool of evidence on the roles of blood- stage immune responses in reducing parasitaemia in humans [11]. Gamma globulins generated by immune adults conferred massive protection in children, elucidating that naturally acquired immunity is antibody-based and mainly against blood-stage malaria. The mode of action can be either against pre- invasion merozoites or against infected erythrocytes. Against merozoites, the response can stop them from invading erythrocytes, or it can lead to self-destruction by neutrophil-mediated parasite killing, phagocytic engulfment, or antibody-dependent cellular inhibition (ADCI) [7]. The humoral immune system can recognize infected red blood cells due to the conformational changes conferred by several merozoites antigens, and once recognized, infected erythrocytes are eliminated through several mechanisms, including obstruction of cytoadhesion to enable spleen clearance and cell- mediated killing [12]. It is well understood that a comprehensive system of antibodies raised against both pre-invasion merozoites and infected red blood cells results in potent protection from clinical symptoms and severe malaria [13].
Historical Perspective of BSVs
Protection against blood-stage malaria was first scientifically studied in the early 1900s when simian blood infected with P. knowlesi elicited effective immune responses in naïve Rhesus macaques [14]. Several experimental studies confirmed that vaccinated macaques performed much better than naïve groups when challenged with parasitized erythrocytes [15]. Intensive research in the mid-20th century demonstrated that immune serum obtained from previously infected macaques conferred passive immunity on the naïve group after immunization, further demonstrating the possibility for an erythrocytic malaria vaccine and paving a long research route that continues to intrigue [16].
Momentum was garnered when a clinical trial that transferred IgG obtained from naturally malaria- immune adults treated infected vulnerable children in malaria-endemic regions by significantly reducing parasitemia [17]. Two key studies conducted 30 years apart in The Gambia and Thailand were instrumental in advancing the novel idea of using serum immunoglobulin as a potential vaccine against erythrocytic malaria [18,19]. A few years later, updated safety-related protocols meant that direct passage of blood components was no longer considered fit for human vaccination. However, these early discoveries remain instrumental in present- day malaria vaccine research. Crucial milestones in the blood-stage malaria vaccine came into fruition when Trager and Jensen (1976) demonstrated the possibility of continuously culturing the P. falciparum parasite [17]. A succeeding study by Mitchell et al. (1977) showed how monkeys could similarly be immunized with the same human parasite, paving the way for whole parasite malaria vaccines (WPV) based on the highly observed initial protection [20,21]. WPVs recorded profound results in eliciting potent immune responses in early trials but were fundamentally impeded by the requirement of FCA as an adjuvant and the complicated immune mechanism associated with the immunization [17]. Confirmatory mice and human studies provided more insights when cultured CD4+ T cells offered protection in naïve experimental groups and a potent non-antibody mediated cellular immune response [22-24].
Overall, these initial findings highlighted the essential roles of antibodies and CD4+ T cells in controlling parasitaemia, but the vaccines involved had not been clinically tested for actual efficacy. Nonetheless, even at present, most effective malaria vaccines rely on antibody production, so the early assumption that a humoral immune response would suffice might not be a mere theory after all.
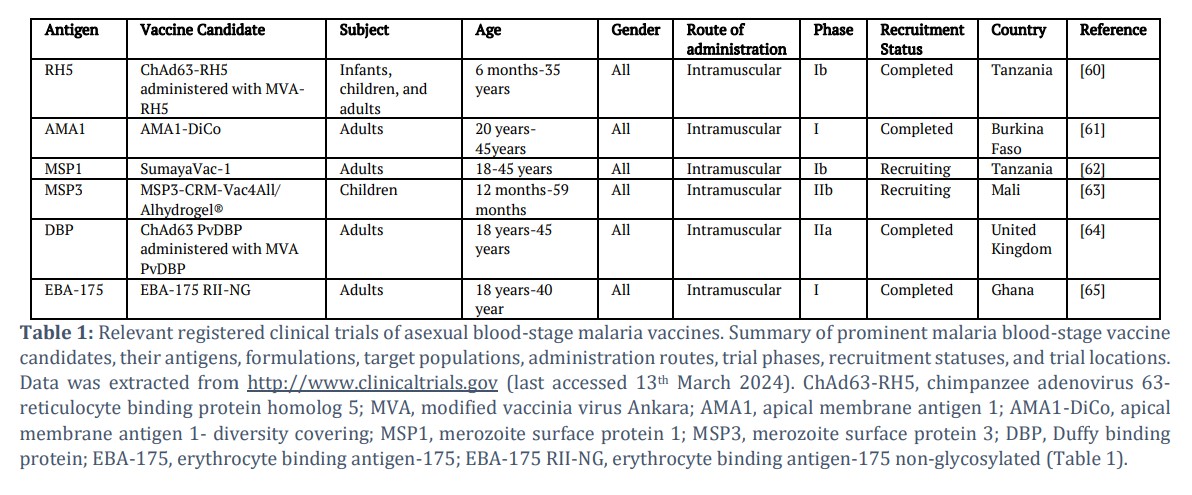
Key Erythrocytic Antigens in Blood-Stage Vaccine Development
Interest in BSVs has been developing since the African IgG experiment in the 1960s [25]. However, as stringent ethical protocols keep evolving, the feasibility of a direct human-to-human antibody transfer waned, giving rise to a more scientific approach to blood-stage malaria control through various merozoites antigens (Table 1). Largely aided by the sequencing of the P. falciparum genome, numerous blood-stage antigens that demonstrate strong immunological potential have been discovered [26]. Different antigens offer different immunological mechanisms, leading to myriad pathways to formulating a highly effective and safe vaccine against blood-stage malaria. At the same time, this denotes the necessity of critical evaluation in choosing suitable vaccine candidates [27].
Research targeting blood-stage antigens has been challenging over the last three decades. No candidate has made it through phase III clinical trials. Polymorphism and non-heterologous efficacy are the most renowned obstacles BSVs must overcome. The widely targeted MSP1 and AMA1 hold a lot of promise in their ability to produce high antibody titres and T- cell responses if the impending genomic challenges are addressed efficiently [25]. There is thus a feasible way for not only MSP1 and AMA1 but several other BSV candidates to futuristically overcome these drawbacks and finally make it beyond CHMI and the final phase of clinical trials.
Reticulocyte-Binding Protein Homolog 5 (RH5)
RH5 is an essential blood-stage antigen that binds basigin on the surface of red blood cells and mediates invasion via the nonredundant sialic acid-independent pathway. It is in the subtelomeric region of chromosome 4, where it harbours species-specific genes involved in host-parasite interactions [28]. Basigin is a multifunctional, highly conserved integral receptor essential for erythrocyte invasion of all Plasmodium spp. [29]. PfRH5 has demonstrated potent immune responses, with multiple and elevated levels of humoral immunity generated in both human and experimental animal studies [30-33]. The main challenges affecting RH5 vaccine research are the very low anti-RH5 antibodies generated by naturally acquired immunity in CHMI studies conducted in Kenya, Papua New Guinea, Mali, and Peru [34-37], gaps in understanding the actual mechanism of action by which anti-RH5 antibody prevents parasite invasion, and the amount of antibodies required for effective protection [28]. Because these obstacles are repairable, RH5 remains one of the most promising blood-stage vaccine candidates in modern-day malaria research.
Apical Membrane Antigen 1 (AMA1)
AMA1 was originally studied in P. falciparum, where it has been cloned. The apical membrane antigen 1 (AMA1) appears as a merozoite surface antigen, vital for identifying and infecting red blood cells [38]. AMA1 belongs to the microneme proteins, usually existing among apicomplexan parasites. It includes a hydrophobic domain similar to the integral membrane protein and is named due to its appearance by the apical complex. AMA1 is understood to migrate to the surface of the merozoite close to the period in which schizonts burst, eliciting a high immunogenic response in tested hosts. It has also been demonstrated to be safe and highly tolerable. AMA1 has experienced mixed results as a sole vaccine, with the majority of clinical trials stopping at phase 1, but yielded some promise when developed in multiepitope vaccines, with an adjuvant, or in combination with other protein components such as MSP1 [39,40]. Low and short-lived efficacy in non-endemic regional individuals, polymorphism, and long-term storage of vaccine formulations are the most pressing factors that deter AMA1’s progress in clinical trials [41]. The issue of polymorphism is being addressed by the increased interest in the highly conserved domain III of AMA1, whereas the combined BSV candidate AMA1-RON2 has thus far shown great promise in terms of efficacy [42]. With the continuous emergence of AMA1-based vaccines, especially in zoonotic malaria P. knowlesi, there is substantial hope in the surge for a successfully approved PkAMA1 vaccine in the upcoming years.
AMA1-Rhoptry Neck Protein-2 (AMA1-RON2)
Even though AMA1 vaccines continue to address the challenges of polymorphism and efficacy, the essential role that this antigen plays in erythrocytic invasion cannot be overemphasized. Thus, researchers continue to explore ways through which a highly efficacious AMA1-based vaccine can be developed. One robust formulation that has shown great promise is the combined immunogenic response elicited by AMA1 and one part of its complex tyrosine kinase receptor, the rhoptry neck protein-2 (RON2) [43]. This formulation generates a potent immune response with long-lived anti-invasion antibodies much higher than sole AMA1 [44]. Additionally, AMA-RON2 offered sterile protection after heterologous challenges in monkeys, along with long-lived immunity [45]. Although experimental in vivo studies may not necessarily translate exactly in humans, vaccines that show promise in pre-clinical trials often perform better than those with relatively inferior impact. One major constraint surrounding the AMA1-RON2 vaccine is the recent discovery that suggests that despite the observable high protection traits, the complex may not be essential for erythrocytic invasion in P. falciparum and P. vivax as opposed to a monomeric AMA1 antigen [46]. Therefore, more knowledge is needed in terms of cross and species-specific conserved interactions of AMA1 and RON2.
Merozoite Surface Protein 1 (MSP1)
MSP1 is designed to block the invasion of red blood cells. In P. falciparum, it differentiates into three variants PfMSP128, PfMSP138, PfMSP142, and PfMSP183 before invasion [41]. Upon invasion, PfMSP1₄₂ is processed into PfMSP1₃₃ and the C-terminal PfMSP1₁₉ fragment, with the latter being directly involved in erythrocyte attack through its interaction with the erythrocyte surface receptor band 3 protein [41]. PfMSP1 vaccines are highly immunogenic and have no safety concerns in CHMI clinical trials [47]. However, in terms of efficacy, both full-length PfMSP1 and PfMSP19 have performed poorly in CHMIs, with multiple immunization dosages required for any significant response [48]. In some reports, PfMSP19 recorded no parasitemia reduction effect in both homologous CHMI and natural infections [49,50]. Aiding MSP1 vaccines with certain adjuvants, receptors, and antigens, as in the Combination B vaccine, has elicited better responses and may thus be the way forward to an effective merozoite surface protein 1 vaccine [40,51].
Duffy-Binding Protein (DPB)
DBP is a prominent candidate for a subunit P. vivax vaccine and is the main parasite protein that invades red blood cells. The role of DBP in the viability of the asexual stage of P. vivax malaria justifies the persistent interest in vaccines targeting this antigen in spite of the many challenges surrounding its perceived licensure [52]. Immunological performances of the most known DBP vaccines against blood-stage P. vivax have been consistently disappointing with little to no robust immunity generated in conducted trials. Most in vivo tested recombinant DBP-II vaccines elicited insufficient mAbs even when multiple dosages were administered in mice [52]. However, akin to AMA1 vaccines, DBP has recently received a revived interest after demonstrating great promise when structural vaccinology tremendously improved anti-DBP antibody titers and showed minimal immunodominant reaction to non-neutralizing polymorphic epitopes [53]. With this new positive outcome, next-generation DBP vaccines could thus be the solution to this increasingly concerning parasite.
Merozoite Surface Protein 3 (MSP3)
The European Malaria Vaccine Initiative (EMVI) and the Pasteur Institute in Paris created a long synthetic peptide MSP3 in clinical trials conducted in Tanzania and Burkina Faso [54]. P. falciparum MSP3 has demonstrated a valuable impact on safety and in eliciting a robust immune response after several investigations [55]. The most developed and clinically tested prospective formulations of this antigen are MSP3-LSP, which is a combination of MSP3 and LSP1 chemically synthesized from the highly conserved region of the 95aa long MSP3, and GMZ2, a blood-stage vaccine candidate designed as a fusion of the less understood recombinant GLURP protein and MSP3, which is presently undergoing clinical trials [55]. High IgG titers were observed in phase I clinical trials of MSP3-LSP involving CHMI individuals in Switzerland, where progression ensued to more diverse trials involving children. However, interest in MSP3-LSP waned after a poor performance in phase II clinical trials [56]. In contrast, GMZ2 is a subject of keen research interest backed by the positive results being obtained from several murine and primate studies with and without adjuvants, indicating that the excellent T- cell and humoral responses it elicits will outperform the other MSP3 vaccine candidates in clinical trials [55]. The major drawbacks of these vaccines are the lack of an efficient method to trigger immune responses in humans like those observed in vivo and the right concentration of MSP3 antigen for potential immunization [57].
Erythrocyte-Binding Antigen 175 (EBA175)
This antigen belongs to the erythrocyte binding proteins (EBP) family, where interaction between erythrocytes and merozoites and the action of substitute invasion pathways is regulated [58]. Emphasis is on the EBA-175 RII NG Vaccine due to its binding and sequence specificity. During invasion, the sialic acid residues of erythrocyte glycophorin A are mainly bound by region II of EBA175’s 175 kDa merozoite surface receptor [57]. This feature makes the protein a potential BSV candidate. A phase 1a clinical trial using EBA175 RII-NG prepared with an aluminium phosphate adjuvant and expressed without glycosylation in Pichia pastoris was carried out in the United States in non-predisposed adults.
The results showed an immune response with a well- tolerated and safely sustained GIA activity [57]. In a subsequent study in semi-immune Ghanaians, the vaccine’s safety and immunogenicity were further studied in a phase 1b trial, and the same profile of the formulation was maintained, providing a positive route to succeeding requisite approval phases [58]. An important consideration for the EBA-175 RII NG vaccine is the varying concentration of the EBA-175 protein in Plasmodium spp. Because of the existing redundant merozoite invasion pathways, the insufficiency of EBA175 as a monomeric vaccine is illustrated, calling for a multivalent, adjuvanted formulation if any reputable milestone is to be achieved [59].
The safety and immunogenicity of ChAd63 RH5 and MVA RH5 were set to be investigated in infants, young children, and adults living in Tanzania in a dose- escalation, age de-escalation randomized double-blind controlled Phase Ib trial. Humoral and cellular immune responses to RH5 were proposed to be evaluated.
The results of the responses are yet to be disclosed. For AMA1-DiCo, the primary objective was to evaluate the vaccine’s safety and the impact of GLASE and Alhydrogel® adjuvants in three doses of 50 µg AMA1-DiCo regimen. In the case of SumayaVac-1, healthy adults living in endemic Tanzania are included in the phase I trial to evaluate the safety and reactogenicity of the MSP1-based vaccine. Children are the targeted group of the MSP3-CRM-Vac4All/Alhydrogel® Vaccine whose phase II clinical trial will reveal information regarding reactogenicity that is linked with the phase I trial. The efficacy of the ChAd63/MVA PvDBP vaccines was assessed in an interventional Phase IIa, CHMI- based trial. The results of this study are not yet disclosed to the public. Semi-immune adults were intramuscularly immunized with EBA-175 RII-NG to reveal information on the optimum dosage level that is safe for humans.
BSVs Versus Pre-Erythrocytic (PEVs) and Transmission-Blocking Vaccines (TBVs)
The morbidity and mortality associated with blood- stage malaria are the leading grounds for the ongoing adventure for an effective erythrocytic vaccine. Developing highly immunogenic, efficacious, and heterologous vaccines presents a promising strategy for malaria control. A fully functional blood-stage vaccine will offer a lot by decreasing the severity of illness. Because they target the symptomatic phase of infection, BSVs have a direct impact on disease manifestation, lowering parasitaemia and thus preventing the infected host from severe consequences. Since the first BSV (Combination B) was tested in endemic regions, reported results have continued to prove that non-immune volunteers can develop immunity when they undergo repeated blood-stage challenges [54]. Additionally, the neutralizing effects of these vaccines can significantly reduce morbidity and mortality, thereby alleviating the burden on healthcare systems and improving patient outcomes.
BSVs can induce broad immune responses by stimulating both antibody-mediated and cell-mediated immunity [41]. Elevated levels of anti-merozoites antibodies have been reported in experimental, in vivo, and clinical studies involving blood-stage malaria antigens, proffering a promising ground for a foreseeable vaccine licensure. During the blood stage, merozoite surface protein often remains in the red blood cell membranes, where its exposure to the immune system triggers a wave of humoral and cellular immune responses, thus enabling a targeted antigen approach for the development of highly immunogenic blood-stage vaccine candidates [41].
BSVs targeting nonredundant invasion pathways (such as the highly conserved PfRH5) have recorded highly neutralizing antibodies in preclinical studies [25]. At the same time, blood-stage vaccine candidates have demonstrated T-cell responses in in vivo studies, leading to potentially much-welcomed diverse immune responses [40].
By reducing and preventing parasite multiplication in the blood, gametocyte development can be inhibited, making the mosquito stage non-progressive. This has been observed in P. vivax Duffy binding protein II (PvDBPII/Matrix-MTM), where a 51% reduction in parasitemia in CHMI was reported [25]. When combined with a transmission-blocking vaccine, any surviving gametocytes will be eliminated either by the anti-multiplication ability of BSVs or by rendering them non-infectious through the immunological effects of TBVs. BSVs also offer reduced complexity of vaccine manufacture and delivery, as demonstrated by the culture of P. falciparum’s blood-stage malaria, where the procedure has been fully defined and established, as opposed to the complex sourcing of sporozoites, thus making the production, genetic manipulation, and attenuation easier [66]. Systemic administration of BSVs proposes a more direct mechanism in comparison to PEVs that require targeting the liver. These vaccines can easily be absorbed and acted upon by the immune components readily available in the blood. Furthermore, BSVs provide a direct impact on clinical disease and render immediate benefits to symptomatic malaria patients by unswervingly addressing the most difficult part of the disease. When malaria gets eliminated at this stage, all potentially devastating consequences, such as cerebral malaria, tissue and organ damage, as well as communal transmission, will all be avoided. In contrast to PEVs and TBVs, which need to be 100% effective for complete malaria control, BSVs provide a more feasible strategy that can make the preceding liver and succeeding sexual stages non-consequential. Prominent BSV candidates are also more understood and easier to express in heterologous systems than TBVs such as Pfs48/45, whose advancement has only recently begun [67]. This is why BSVs are considered essential if the sought malaria eradication goal is to be realized.
Advances In Malaria Vaccine Technology
Until now, the R21/Matrix-M is the only efficacious malaria vaccine approved for the public. Having met the WHO’s 75% efficacy goal, the impact that this vaccine will have in the coming years is being optimistically anticipated. However, to realize malaria elimination and eradication goals, a liver-stage vaccine must clear 100% of sporozoites to prevent progression into the blood stage. Due to the compounding difficulties that deter the production of an anti- sporozoites vaccine that will provide sterile protection, BSVs are inevitably vital in the malaria eradication pathway. To achieve this, efficient assessment of prevalence and enhancement of current approaches to BSVs are undoubtedly needed [68]. Advances in the discovery of novel human-tolerant adjuvants play a crucial role in enhancing the immunogenicity of blood- stage malaria vaccines by modulating immune responses to achieve optimal protection against the parasite while minimizing adverse reactions [69]. A practical example is the enhanced squalene-based oil- in-water emulsions such as MF59, where efficacy is improved by promoting the formation of a depot at the injection site, facilitating antigen presentation, and inducing robust antibody and cellular immune responses [70]. Moreover, the utilization of immune regulators such as OX40, CD40 and CD28 have been shown to accelerate cellular and humoral immunity [67,71]. Another area of relevance in improving BSVs is advanced nanotechnology, where there exists significant promise in vaccine delivery. Sophisticated nanoparticles like VLPs, alumina nanoparticles, magnesium oxide nanoparticles, and nano lipids enable precise antigen targeting, controlled release, and enhanced immunogenicity [72-76]. They can encapsulate vaccine antigens, protecting them from degradation and facilitating antigen presentation to immune cells [77].
Additionally, surface modification techniques can tailor nanoparticles for specific immune cell interactions, maximizing vaccine efficacy [78]. When these various fields integrate effectively, BSV antigens will experience a boom in immunogenicity, sequence variation, safety, efficacy, and rapid development.
Prospective Directions and Emerging Trends
Advancing technologies buoy future perspectives for blood-stage malaria vaccines. Approaches that aim to overcome current challenges and knowledge gaps hold immense potential for global health impact. Novel vaccine platforms, such as viral vectors, virus-like particles, and nucleic acid-based vaccines, offer unprecedented opportunities for antigen delivery, immune stimulation, and easy logistics [79]. Although predominant in cancer research, advances in adjuvant design, such as Toll-like receptor agonists and nanoparticle-based formulations, provide promising pathways to enhance blood-stage vaccine immunogenicity and efficacy [80]. Systems biology and immunoinformatics can rationalize antigen combinations to induce broad and long-lasting immunity while significantly limiting antigen sequence variation among Plasmodium spp. [81]. Similarly, multi-allele vaccine formulations can address the polymorphism of essential BSV candidates and strengthen their performance in late-phase clinical trials [82]. These approaches have led to a resurgence of interest in whole-killed BSVs where most of the challenges associated with subunit BSVs are seemingly overcome [83]. However, it remains to be seen whether the safety concern, which is the main constraint of whole-killed BSVs, can be resolved. The approval of RTS,S and R21/Matrix-M sets a path for more anti-sporozoites vaccines. However, in the case of this overwhelming disease, more may not necessarily mean best if a 100% effective pre- erythrocytic vaccine remains out of reach. To ascertain the ultimate “malaria eradication” goal, the development of a highly immunogenic, protective, safe, and efficacious blood-stage vaccine will be an equally-if-not-more-essential contributor. Achieving this goal is challenging, but with the continuing promise of modern innovations discussed in this review, a fully recommended and efficacious blood-stage vaccine is possible.
Tables and Figures
Acknowledgement
This work was supported by the Ministry of Higher Education, Malaysia (MOHE) under the Fundamental Research Grant Scheme [FRGS/1/2021/SKK0/UNISZA/02/5]. We would like to thank the Ministry of Higher Education Malaysia (MOHE) for supporting the A.S.O postdoctoral fellowship.
Authors Contributions
A.S.O. developed the literature search strategy, selection criteria, and scope of the manuscript. Aminu, M.A. performed and analysed all included literature and wrote the manuscript with Rehman, A.U. A.M.S. made a significant contribution to manuscript revision during the preparation of the manuscript. The critical trial reports were reviewed and written by A.A.B.
![]() References
References
- Anteneh M, Asres MS, Legese GL, Alemayehu MA, Woldesenbet D, et al. Treatment outcomes and associated factors in severe malaria patients at University of Gondar Hospital, Northwest Ethiopia: A retrospective study (2020–2023). Plos one, (2024); 19(12): e0309681.
- Alkurbi MO. Plausible inhibitors of malaria parasite Plasmodium falciparum 3D7 ATP-dependent DNA helicase. Advancements in Life Sciences, (2024); 10(4): 644-650.
- Pirahmadi S, Afzali S, Zargar M, Zakeri S, Mehrizi AA. How can we develop an effective subunit vaccine to achieve successful malaria eradication? Microbial Pathogenesis, (2021); 160(1): 105203.
- Etefia E, Inyang-Etoh P. Malaria Vaccine Development: Challenges and Prospects. Medical and Pharmaceutical Journal, (2023); 2(1): 28-42.
- Datoo MS, Natama HM, Some A, Bellamy D, Traore O, et al. Efficacy and immunogenicity of R21/Matrix-M vaccine against clinical malaria after 2 years' follow-up in children in Burkina Faso: a phase 1/2b randomised controlled trial. The Lancet Infectious Diseases, (2022); 22(12): 1728-1736.
- Aminu MA, Ur Rehman A, Azizan NA, Othman AS. Malaria and Cardiometabolic Health: A Review of Its Impact on Metabolic and Cardiovascular Disorders. Malaysian Journal of Medicine & Health Sciences, (2024); 20(10): 242-251.
- Cowman AF, Healer J, Marapana D, Marsh K. Malaria: Biology and Disease. Cell, (2016); 167(3): 610-624.
- Barry A, Hansen D. Naturally acquired immunity to malaria. Parasitology, (2016); 143(2): 125-128.
- Othman AS, Marin-Mogollon C, Salman AM, Franke-Fayard BM, Janse CJ, et al. The use of transgenic parasites in malaria vaccine research. Expert review of vaccines, (2017); 16(7): 685-697.
- Wu Y, Sinden RE, Churcher TS, Tsuboi T, Yusibov V. Development of malaria transmission-blocking vaccines: from concept to product. Adv Parasitol, (2015); 89109-152.
- Khoury DS, Aogo R, Randriafanomezantsoa-Radohery G, McCaw JM, Simpson JA, et al. Within-host modeling of blood-stage malaria. Immunol Rev, (2018); 285(1): 168-193.
- Chan J-A, Fowkes FJ, Beeson JG. Surface antigens of Plasmodium falciparum-infected erythrocytes as immune targets and malaria vaccine candidates. Cellular and Molecular Life Sciences, (2014); 713633-3657.
- Langhorne J, Ndungu FM, Sponaas A-M, Marsh K. Immunity to malaria: more questions than answers. Nature immunology, (2008); 9(7): 725-732.
- Freund J, Thomson KJ, et al. Immunization of monkeys against malaria by means of killed parasites with adjuvants. American Journal of Tropical Medicine, (1948); 28(1): 1-22.
- McCarthy JS, Good MF. Whole parasite blood stage malaria vaccines: a convergence of evidence. Human Vaccines, (2010); 6(1): 114-123.
- Coggeshall LT, Kumm HW. Demonstration of Passive Immunity in Experimental Monkey Malaria. The Journal of Experimental Medicine, (1937); 66(2): 177-190.
- Good MF, Miller LH. Interpreting challenge data from early phase malaria blood stage vaccine trials. Expert Rev Vaccines, (2018); 17(3): 189-196.
- Cohen S, Mc GI, Carrington S. Gamma-globulin and acquired immunity to human malaria. Nature, (1961); 192733-737.
- Bouharoun-Tayoun H, Attanath P, Sabchareon A, Chongsuphajaisiddhi T, Druilhe P. Antibodies that protect humans against Plasmodium falciparum blood stages do not on their own inhibit parasite growth and invasion in vitro, but act in cooperation with monocytes. The Journal of experimental medicine, (1990); 172(6): 1633-1641.
- Mitchell GH, Richards WH, Butcher GA, Cohen S. Merozoite vaccination of douroucouli monkeys against falciparum malaria. Lancet, (1977); 1(8026): 1335-1338.
- Trager W, Jensen JB. Human malaria parasites in continuous culture. Science, (1976); 193(4254): 673-675.
- Brake DA, Weidanz WP, Long CA. Antigen-specific, interleukin 2-propagated T lymphocytes confer resistance to a murine malarial parasite, Plasmodium chabaudi adami. Journal of Immunology, (1986); 137(1): 347-352.
- Amante FH, Good MF. Prolonged Th1‐like response generated by a Plasmodium yoeli‐specific T cell clone allows complete clearance of infection in reconstituted mice. Parasite immunology, (1997); 19(3): 111-126.
- Pombo DJ, Lawrence G, Hirunpetcharat C, Rzepczyk C, Bryden M, et al. Immunity to malaria after administration of ultra-low doses of red cells infected with Plasmodium falciparum. Lancet, (2002); 360(9333): 610-617.
- Duffy PE. Current approaches to malaria vaccines. Current Opinion in Microbiology, (2022); 70102227.
- Muh F, Lee SK, Hoque MR, Han JH, Park JH, et al. In vitro invasion inhibition assay using antibodies against Plasmodium knowlesi Duffy binding protein alpha and apical membrane antigen protein 1 in human erythrocyte-adapted P. knowlesi A1-H.1 strain. Malaria Journal, (2018); 17(1): 272.
- Healer J, Chiu CY, Hansen DS. Mechanisms of naturally acquired immunity to P. falciparum and approaches to identify merozoite antigen targets. Parasitology, (2018); 145(7): 839-847.
- Ord RL, Rodriguez M, Lobo CA. Malaria invasion ligand RH5 and its prime candidacy in blood-stage malaria vaccine design. Human Vaccines & Immunotherapeutics, (2015); 11(6): 1465-1473.
- Crosnier C, Bustamante LY, Bartholdson SJ, Bei AK, Theron M, et al. Basigin is a receptor essential for erythrocyte invasion by Plasmodium falciparum. Nature, (2011); 480(7378): 534-537.
- Douglas AD, Baldeviano GC, Lucas CM, Lugo-Roman LA, Crosnier C, et al. A PfRH5-based vaccine is efficacious against heterologous strain blood-stage Plasmodium falciparum infection in aotus monkeys. Cell host & microbe, (2015); 17(1): 130-139.
- Oboh MA, Asmorom N, Falade C, Ojurongbe O, Thomas BN. High genetic and haplotype diversity in vaccine candidate Pfceltos but not Pfrh5 among malaria-infected children in Ibadan, Nigeria. PeerJ, (2023); 11e16519.
- Jamwal A, Constantin CF, Hirschi S, Henrich S, Bildl W, et al. Erythrocyte invasion-neutralising antibodies prevent Plasmodium falciparum RH5 from binding to basigin-containing membrane protein complexes. Elife, (2023); 12e83681.
- Miura K, Diouf A, Fay MP, Barrett JR, Payne RO, et al. Assessment of precision in growth inhibition assay (GIA) using human anti-PfRH5 antibodies. Malaria Journal, (2023); 22(1): 159.
- Villasis E, Lopez-Perez M, Torres K, Gamboa D, Neyra V, et al. Anti-Plasmodium falciparum invasion ligand antibodies in a low malaria transmission region, Loreto, Peru. Malaria Journal, (2012); 11(1): 361.
- Douglas AD, Williams AR, Illingworth JJ, Kamuyu G, Biswas S, et al. The blood-stage malaria antigen PfRH5 is susceptible to vaccine-inducible cross-strain neutralizing antibody. Nature communications, (2011); 2(1): 601.
- Tran TM, Ongoiba A, Coursen J, Crosnier C, Diouf A, et al. Naturally acquired antibodies specific for Plasmodium falciparum reticulocyte-binding protein homologue 5 inhibit parasite growth and predict protection from malaria. The Journal of Infectious Diseases, (2014); 209(5): 789-798.
- Chiu CY, Healer J, Thompson JK, Chen L, Kaul A, et al. Association of antibodies to Plasmodium falciparum reticulocyte binding protein homolog 5 with protection from clinical malaria. Frontiers in Microbiology, (2014); 5314.
- Chua CY, Lee PC, Lau TY. Analysis of polymorphisms and selective pressures on ama1 gene in Plasmodium knowlesi isolates from Sabah, Malaysia. Journal of Genetics, (2017); 96(4): 653-663.
- Ouattara A, Niangaly A, Adams M, Coulibaly D, Kone AK, et al. Epitope-based sieve analysis of Plasmodium falciparum sequences from a FMP2. 1/AS02A vaccine trial is consistent with differential vaccine efficacy against immunologically relevant AMA1 variants. Vaccine, (2020); 38(35): 5700-5706.
- Milán-Noris EM, Monreal-Escalante E, Rosales-Mendoza S, Soria-Guerra RE, Radwan O, et al. An AMA1/MSP1 19 Adjuvanted Malaria Transplastomic Plant-Based Vaccine Induces Immune Responses in Test Animals. Molecular Biotechnology, (2020); 62534-545.
- Zheng J, Pan H, Gu Y, Zuo X, Ran N, et al. Prospects for Malaria Vaccines: Pre-Erythrocytic Stages, Blood Stages, and Transmission-Blocking Stages. Biomed Research International, (2019); 20199751471.
- Venkatesh V (2023) DESIGNING mRNA BASED MULTISTAGE VACCINE FOR MALARIA: Johns Hopkins University.
- Haron FN, Azazi A, Chua KH, Lim YAL, Lee PC, et al. Small-scale optimization of recombinant Plasmodium knowlesi apical membrane antigen 1 (rPkAMA1) expression in Pichia pastoris. Asian Journal of Medicine and Biomedicine, (2020); 4(SI1): 79-90.
- Duffy PE, Patrick Gorres J. Malaria vaccines since 2000: progress, priorities, products. npj Vaccines, (2020); 5(1): 48.
- Srinivasan P, Baldeviano GC, Miura K, Diouf A, Ventocilla JA, et al. A malaria vaccine protects Aotus monkeys against virulent Plasmodium falciparum infection. NPJ Vaccines, (2017); 2(1): 14.
- Drew DR, Wilson DW, Weiss GE, Yeoh LM, I GH, et al. Defining species-specific and conserved interactions of apical membrane protein 1 during erythrocyte invasion in malaria to inform multi-species vaccines. Cellular and Molecular Life Sciences, (2023); 80(3): 74.
- Thomson-Luque R, Stabler TC, Fürle K, Silva JC, Daubenberger C. Plasmodium falciparum merozoite surface protein 1 as asexual blood stage malaria vaccine candidate. Expert Review of Vaccines, (2024); 23(1): 160-173.
- Rathay V, Furle K, Kiehl V, Ulmer A, Lanzer M, et al. IgG Subclass Switch in Volunteers Repeatedly Immunized with the Full-Length Plasmodium falciparum Merozoite Surface Protein 1 (MSP1). Vaccines (Basel), (2024); 12(2): 208.
- Xu Q, Liu S, Kassegne K, Yang B, Lu J, et al. Genetic diversity and immunogenicity of the merozoite surface protein 1 C-terminal 19-kDa fragment (MSP119) of Plasmodium ovale imported from Africa to China. Parasites & Vectors, (2021); 14(1): 583.
- Chitnis CE, Mukherjee P, Mehta S, Yazdani SS, Dhawan S, et al. Correction: Phase I Clinical Trial of a Recombinant Blood Stage Vaccine Candidate for Plasmodium falciparum Malaria Based on MSP1 and EBA175. PLoS One, (2015); 10(9): e0137816.
- Rosenkranz M, Furle K, Hibbert J, Ulmer A, Ali A, et al. Multifunctional IgG/IgM antibodies and cellular cytotoxicity are elicited by the full-length MSP1 SumayaVac-1 malaria vaccine. NPJ Vaccines, (2023); 8(1): 112.
- Dickey TH, Tolia NH. Designing an effective malaria vaccine targeting Plasmodium vivax Duffy-binding protein. Trends in Parasitology, (2023); 39(10): 850-858.
- Chen E, Salinas ND, Ntumngia FB, Adams JH, Tolia NH. Structural analysis of the synthetic Duffy Binding Protein (DBP) antigen DEKnull relevant for Plasmodium vivax malaria vaccine design. PLoS Neglected Tropical Diseases, (2015); 9(3): e0003644.
- Genton B, Reed ZH. Asexual blood-stage malaria vaccine development: facing the challenges. Current Opinion in Infectectious Diseases, (2007); 20(5): 467-475.
- Alves KCS, Guimarães JM, Almeida MEMd, Mariúba LAM. Plasmodium falciparum merozoite surface protein 3 as a vaccine candidate: a brief review. Revista do Instituto de Medicina Tropical de São Paulo, (2022); 64e23.
- 56. Salamanca DR, Gomez M, Camargo A, Cuy-Chaparro L, Molina-Franky J, et al. Plasmodium falciparum Blood Stage Antimalarial Vaccines: An Analysis of Ongoing Clinical Trials and New Perspectives Related to Synthetic Vaccines. Frontiers in Microbiology, (2019); 102712.
- Takashima E, Otsuki H, Morita M, Ito D, Nagaoka H, et al. The Need for Novel Asexual Blood-Stage Malaria Vaccine Candidates for Plasmodium falciparum. Biomolecules, (2024); 14(1): 100.
- Yagi M, Palacpac NM, Ito K, Oishi Y, Itagaki S, et al. Antibody titres and boosting after natural malaria infection in BK-SE36 vaccine responders during a follow-up study in Uganda. Scientific Reports, (2016); 6(1): 34363.
- Bei AK, Membi CD, Rayner JC, Mubi M, Ngasala B, et al. Variant merozoite protein expression is associated with erythrocyte invasion phenotypes in Plasmodium falciparum isolates from Tanzania. Molecular and biochemical parasitology, (2007); 153(1): 66.
- Minassian AM (2018) Safety and Immunogenicity of ChAd63 RH5 and MVA RH5 in Adults, Young Children and Infants Living in Tanzania. University of Oxford. NCT03435874 NCT03435874.
- Launay O, Sirima S (2013) Safety and Immunogenicity of Recombinant Pichia Pastoris AMA1-DiCo Candidate Malaria Vaccine With GLA-SE and Alhydrogel ® as Adjuvant in Healthy Malaria Non-Exposed European and Malaria Exposed African Adults:a Staggered Phase Ia/Ib, Randomised, Double-blind, Multi-Centre Trial. NCT02014727 NCT02014727.
- Daubenberger C, Olotu A. A Randomised, Controlled, Double-blind, Parallel Group, Single Center Phase Ib Trial to Assess Safety, Reactogenicity, Immunogenicity and Parasite Growth Rates After Controlled Human Malaria Infection of a Candidate Dual-stage Malaria Vaccine, SumayaVac-1 (MSP-1 With GLA-SE as Adjuvant) in Healthy Malaria Exposed Adults of African Origin Aged 18-45 Years. (2024,May10);https://www.cms.ihi.or.tz/grants_portifolio/A%20randomised,%20controlled,%20double-blind,%20parallel%20group,%20single%20center%20Phase%20Ib%20trial%20to%20assess%20safety,%20reactogenicity,%20immunogenicity%20and%20parasite%20growth%20rates%20after%20Controlled%20Human%20Malaria%20Infection%20of%20a%20candidate%20dual-stage%20malaria%20vaccine,%20SumayaVac-1%20(MSP-1%20with%20GLA-SE%20as%20adjuvant)%20in%20healthy%20malaria%20exposed%20adults%20of%20African%20origin%20aged%2018-45%20years.
- Thera M (2023) A Phase 1b/2b Double Blind, Randomized, Controlled Study of the Safety, Immunogenicity, and Efficacy of Malaria Vaccine Candidate MSP3-CRM-Vac4All/ Alhydrogel® in Young Children in Mali. NCT05776017 NCT05776017.
- Minassian AM (2019) A Phase IIa Challenge Study to Assess Efficacy of the Plasmodium Vivax Malaria Vaccine Candidates ChAd63 PvDBP and MVA PvDBP in Healthy Adults Living in the UK. NCT04009096 NCT04009096.
- Koram KA, Adu B, Ocran J, Karikari YS, Adu-Amankwah S, et al. (2009) Phase I, Double-Blinded, Placebo-Controlled Dosage-Escalation Study of the Safety and Immunogenicity of EBA-175 RII-NG Malaria Vaccine Administered Intramuscularly in Semi-immune Adults. NCT01026246 NCT01026246.
- Cai J, Chen S, Zhu F, Lu X, Liu T, et al. Whole-Killed Blood-Stage Vaccine: Is It Worthwhile to Further Develop It to Control Malaria? Frontiers in Microbiology, (2021); 12670775.
- Othman AS, Franke-Fayard BM, Imai T, van der Gracht ETI, Redeker A, et al. OX40 Stimulation Enhances Protective Immune Responses Induced After Vaccination With Attenuated Malaria Parasites. Frontiers in Cellular and Infection Microbiology, (2018); 8247.
- Khan SA, Ahmad I, Khan W, Khan H, Akbar S. The contagious nature of SARS-CoV-2 omicron variant and vaccine efficacy. Advancements in Life Sciences, (2023); 9(4): 429-436.
- Bonam SR, Renia L, Tadepalli G, Bayry J, Kumar HMS. Plasmodium falciparum Malaria Vaccines and Vaccine Adjuvants. Vaccines (Basel), (2021); 9(10): 1072.
- Leyba AL, Razmjoo M, Francian A, Bierle A, Roberts B, et al. Overcoming Barriers of Vaccination Against Malaria Using Microneedles and TRIO. University of New Mexico (2023).
- Yao X, Wu J, Lin M, Sun W, He X, et al. Increased CD40 Expression Enhances Early STING-Mediated Type I Interferon Response and Host Survival in a Rodent Malaria Model. PLOS Pathogens, (2016); 12(10): e1005930.
- Lee SH, Chu KB, Kang HJ, Basak S, Kim MJ, et al. Virus-like particles expressing Plasmodium berghei MSP-8 induce protection against P. berghei infection. Parasite Immunology, (2020); 42(11): e12781.
- Paliwal H, Prajapati BG, Parihar A, Ganugula S, Patel JK, et al. (2023) Solid Lipid Nanoparticles in Malaria. Malarial Drug Delivery Systems: Advances in Treatment of Infectious Diseases: Springer. pp. 113-137.
- Hamad TG, Salman TA. Investigation of the Impact of Environmentally Friendly prepared Alumina Nanoparticles on Bacterial Activity. Advancements in Life Sciences, (2023); 1095-100.
- Imran M, Alsayeqh AF. Antiparasitic Activity of Chemically Synthesized Magnesium Oxide Nanoparticles against Small Ruminant Haemonchosis. Advancements in Life Sciences, (2022); 9(3): 356-362.
- Ur Rehman A, Aminu MA, Nasir MH, Othman AS. Engineered Virus-Like Particles (VLPs): A Modern Approach to Infectious Diseases and Metabolic Disorders. Malaysian Journal of Medicine & Health Sciences, (2024); 20.
- King LD, Pulido D, Barrett JR, Davies H, Quinkert D, et al. Preclinical Development of a Stabilized RH5 Virus-Like Particle Vaccine that Induces Improved Anti-Malarial Antibodies. BioRxiv, (2024); 2024.2001. 2004.574181.
- Xu Y, Fourniols T, Labrak Y, Preat V, Beloqui A, et al. Surface Modification of Lipid-Based Nanoparticles. ACS Nano, (2022); 16(5): 7168-7196.
- Mascola JR, Fauci AS. Novel vaccine technologies for the 21st century. Nature Reviews Immunology, (2020); 20(2): 87-88.
- Hu Y, Smith D, Frazier E, Zhao Z, Zhang C. Toll-like receptor 9 agonists as adjuvants for nanoparticle-based nicotine vaccine. Molecular Pharmaceutics, (2021); 18(3): 1293-1304.
- Ilyas H, Afzal A, Abbas Z, Noor S, Ullah I, et al. Commands of Synthetic Biology to Modernize and Re-design the Biological Systems. Advancements in Life Sciences, (2024); 11(3): 549-557.
- Terheggen U, Drew DR, Hodder AN, Cross NJ, Mugyenyi CK, et al. Limited antigenic diversity of Plasmodium falciparum apical membrane antigen 1 supports the development of effective multi-allele vaccines. BMC Medicine, (2014); 12183.
- Good MF, Stanisic DI. Whole parasite vaccines for the asexual blood stages of Plasmodium. Immunological Reviews, (2020); 293(1): 270-282.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0