

Full Length Research Article
Purification, Characterization, and Functional Analysis of Isocitrate Lyase Isoforms from Corn Scutella using Ion-exchange Chromatography
Saba Hadi1*, Zahraa B. Mohammed2, Huda F. Ramadan2
Adv. life sci., vol. 12, no. 2, pp. 445-450, May 2025
*– Corresponding Author: Saba Hadi (Email: dr.sabahadi@uomustansiriyah.edu.iq)
Authors' Affiliations 1. Biology Department, College of Science, Mustansiriyah University, Baghdad 10001 – Iraq
2. Department of Biology\College of Education for Women, University of Kirkuk, Kirkuk – Iraq
2. Department of Biology\College of Education for Women, University of Kirkuk, Kirkuk – Iraq
[Date Received: 14/01/2024; Date Revised: 16/02/2025; Date Available Online: 31/08/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: The work described herein investigates the properties of the enzyme isocitrate lyase, which was purified from corn plants using advanced techniques such as sulfate precipitation and ion exchange chromatography. The results indicated the presence of two forms of the enzyme, ICL1 and ICL2, which differed in their molecular weights and enzymatic activities. The optimum pH for ICL1 was found to be 7.5, while for ICL2, it was 6. Additionally, the effects of glycine and glycolate on the enzyme's activity were studied, revealing elevated activities at the optimal concentrations of these substances.
Methods: The isocitrate lyase enzyme was purified from corn plants using ion exchange chromatography and sulfate precipitation techniques. Enzyme activity was assessed using spectrophotometric methods, and the molecular weight was determined through gel chromatography. Studies were conducted to investigate the influence of pH, glycine, and glycolate on enzyme activity.
Results: Two isoforms of isocitrate lyase, ICL1 and ICL2, were purified, exhibiting molecular masses of 164 kDa and 208 kDa, respectively. ICL1 demonstrated optimum activity at pH 7.5, while ICL2 exhibited optimum activity at pH 6. In this study, specific concentrations of glycine and glycolate were found to enhance the enzymatic activities of both isoforms.
Conclusion: This research provides significant insights into the characteristics of the isocitrate lyase enzyme in corn plants. The data indicate the presence of distinct enzyme forms with specific interactions in varying environmental conditions, which may be applicable in agricultural practices to increase crop yield and improve the metabolic turnover of organic acids.
Keywords: Isocitrate lyase isoenzymes; Corn scutella; Chromatography; Glycine; Glycolate
Introduction![]()
The isocitrate lyase enzyme, EC 4.1.3.1, catalyzes a reversible reaction of aldol-type fragmentation into glyoxylate and succinate. Along with malate synthase, isocitrate lyase ICL is considered the key enzyme in the glyoxylate cycle. It appears at early stages of oil plant seed germination and becomes less active as stored fat breaks down [1,2].
Isocitrate lyase is widespread in bacteria, mushrooms and higher plants. In metazoans, the enzyme is present in some nematodes during the adult stage, post-embryonic larvae and embryos and probably in some arthropods. The presence of ICL and the glyoxylate cycle in vertebrates remains debated [3,4]. The regulation of ICL activity during seed germination is one of the most important problems, as it provides the key to general patterns of enzymatic regulation during ontogenesis [5,6].
To date, ICL has been purified to homogeneity from Pseudomonas indigofera bacteria, mushrooms, higher plants, nematodes, acarines, insects, and rats [4,7]. Among plants, the most frequent occurrence of ICL was found in seeds accumulating fat. Nevertheless, there is also some information about its occurrence in other organs, for example, fruit and green leaves [6-8]. ICL purification from different sources generally involves ammonium sulfate fractionation, DEAE-Toyopearl ion-exchange chromatography, and gel filtration on different types of Sephadex. ICL purification (including dialysis, acetone fractionation, and chromatography on Sephadex G-200 and DEAE cellulose) from developing Hyalomma dromedarii acarine embryos has also been described [9,10].
Since the use of common sorbents normally does not provide an effective means of separating enzyme isoforms, the purpose of this study is to devise a method for separating isocitrate lyase isoenzymes from corn scutella by using ion-exchange chromatography.
![]() Methods
Methods
Corn Scutella of 4-day-old etiolated corn germs (Baghdad sort), hydroponically grown at t = 25 °C, were used as the target of analysis.
The activity of isocitrate lyase (EC 4.1.3.1) was determined spectrophotometrically by an SF-46 spectrophotometer, following light absorbance change at a wavelength of 324 nm. This absorbance change is due to the formation of phenyl hydrazine complex with glyoxylate [8]. The spectrophotometric assay mixture was prepared with 50 mM Tris-HCl buffer (pH 7.5), supplemented with 5 mM magnesium chloride (MgCl₂), 4 mM dithiothreitol (DTT), 2 mM sodium isocitrate, and 4 mM phenylhydrazine hydrochloride.
The amount of isocitrate lyase producing 1 μM glyoxylate in 1 minute at t = 25 °C was reserved as the enzyme activity unit. The total amount of protein was calculated using the Lowry method [9].
A special purification pattern was developed for obtaining highly-purified isocitrate lyase specimens, which included 4 stages. All the operations were carried out at 0-4°C temperature according to Scherbyna et al., [10]:
- Obtaining homogenate: weighed amount of plant material (m = 5.0g) was homogenized at the ratio of 1:5 with the following separation medium composition: 50 mM Tris-HCl buffer (рН 7.5), including 3 mM EDTA, 0.1М MgCl2 and 5 mM DTT. Centrifugation was carried out for 5 minutes at 5000 g.
- Proteins were fractionated with ammonium sulfate within 40% saturation limits. The obtained solution was centrifuged for 20 minutes at a weight of 12000 g.
- The protein fraction was separated using a Sephadex G-25 column (1.5 × 20 cm; Pharmacia, Sweden). Elution was carried out with 50 mM Tris-HCl buffer (pH 7.5) at a flow rate of 10–20 mL per hour to remove unwanted proteins.
- Ion exchange chromatography was performed using a DEAE cellulose column (1.5 × 15 cm; Whatman, UK) pre-equilibrated with 50 mM Tris-HCl buffer (pH 7.5). Enzyme elution was carried out using a linear KCl gradient (40-150 mM) in the equilibration buffer.
The gel chromatography technique on a 2 х 40 cm column with Sephadex G-200 was used to determine the quaternary structure and the molecular weight of native ICL [10]. The elution volume (Ve) was calculated. The void volume (Vo) of the column was calculated with blue dextran (Serva). The molecular weight of the analyzed enzyme was calculated using the formula obtained from the calibration curve according to Dajani et al. (1996) [1]:
Electrophoretic analysis of proteins was carried out in a 7.5% polyacrylamide gel [11]. Universal development of proteins was achieved with silver nitrate. Gels were stored in 7% acetic acid solution.
The specific identification of the enzyme was determined using modified Schiff reagent [12]. SDS-PAGE electrophoresis was performed with a 12.5% polyacrylamide gel concentration. Every specimen contained 3-5 µg protein.
Standard marker proteins (Sigma) were used to plot the calibration curve [13]. The testing was done in three replications. Analytical evaluation for each sample was carried out in two replications. Typical experiment data are presented in the table and the figures, where each value represents the arithmetic mean. The statistical Student’s criterion was used for mathematical treatment [14].
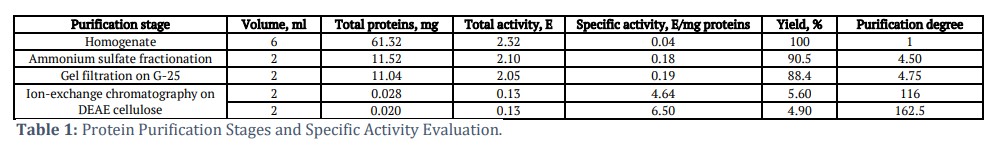
The study identified two distinct isocitrate lyase (ICL) isoforms in corn scutella, with different electrophoretic mobilities. Using electrophoresis, two enzyme isoforms with varying mobility were observed, corresponding to molecular forms of ICL1 and ICL2. The enzyme purification process consisted of four stages, resulting in specific activities of 4.64 E/mg for ICL1 and 6.50 E/mg for ICL2, with purification degrees of 116 times and 162.5 times, respectively. The enzyme activity elution profile after ion-exchange chromatography on DEAE cellulose showed maximum activity for the first and second isoforms at KCl concentrations of 60.2 mM and 94.2 mM, respectively.
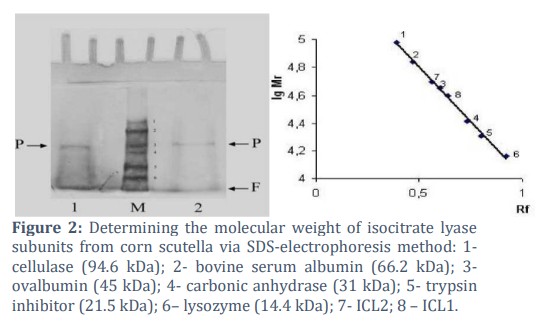
Electrophoretic analysis revealed that the isoforms had distinct Rf values: ICL1 (Rf-0.29) and ICL2 (Rf-0.25), with a single protein band observed for each isoform. Gel chromatography on Sephadex G-200 revealed two peaks corresponding to molecular weights of 164 kDa and 208 kDa. Electrophoresis with SDS provided molecular weights of 43±1.2 kDa for ICL1 and 48±0.5 kDa for ICL2. These results suggest that ICL is a tetrameric protein. Kinetic studies indicated that both isoforms followed Michaelis-Menten kinetics, with ICL1 showing a Michaelis constant (Km) value of 55.6 μM and ICL2 exhibiting a lower substrate affinity (Km = 83.3 μM). The enzyme activity was pH-dependent, with ICL1 showing peak activity at pH 7.5 and ICL2 at pH 6.0.
The effect of glycine and glycolate concentrations on ICL activity was also investigated. Glycine activated both isoforms, but with differing concentrations, with ICL1 showing increased activity at 5 μM and ICL2 at 10 μM. Glycolate had a significant effect on both isoforms, with ICL1 being activated at lower concentrations (5 μM) and ICL2 at higher concentrations (100-500 μM).
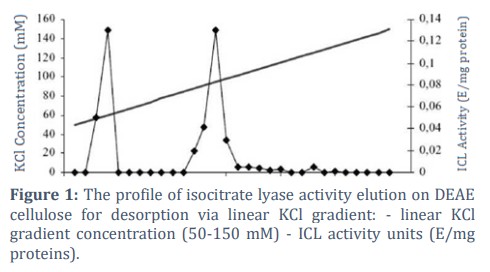
The key stage in obtaining highly-purified isoforms of isocitrate lyase was the application of ion-exchange chromatography on DEAE cellulose with a linear KCl gradient (50-150 mM). Two peak values of enzyme activity elution were obtained during the purification of ICL from the scutella. The 1st and 2nd isoforms exhibited enhanced elution at 60.2 mM and 94.2 mM KCl solution concentrations. The profile of isocitrate lyase activity elution after DEAE cellulose is illustrated in Figure 1.
Electrophoretic analysis of purified specimens revealed a single protein band in polyacrylamide gel during universal protein staining or specific development. Thus, it was found that a corn scutellum had 2 forms of enzyme with diverse electrophoretic migration rates: ICL1 with Rf-0.29 and ICL2 with Rf-0.25.
During the gel chromatography on Sephadex G-200, ICL from corn scutella was eluted as two peaks corresponding to molecular weights of 164 and 208 kDa. Electrophoretic analysis of proteins in the presence of SDS allowed determination of the molecular weight of each subunit, which was 43±1.2 kDa for ICL1 and 48±0.5 kDa for ICL2 in Figure 2.
The results obtained with gel chromatography on Sephadex G-200 and the denaturing electrophoresis data suggest that ICL represents a tetrameric protein.
During the analysis of kinetic properties for homogeneous isocitrate lyase specimens in corn scutella, it was established that the enzyme followed Michaelis–Menten kinetics. The isocitrate Km value was 55.6 μM for ICL1. Substrate affinity for ICL2 was lower compared to the first enzyme form (Km = 83.3 μM). The data indicate that the enzyme's isocitrate affinity varies significantly across different metabolite concentrations and is comparable to that of other substances.
The investigation of the effect of hydrogen ion concentration on ICL activity in scutella showed that such ICL activity relationship was bell-shaped for both isoforms. In these conditions, the highest activity level for ICL1 was observed at рН 7.5. Optimum рН was slightly offset into the acidic range of values and equalled 6 for the second isoform. The optimum pH for ICL from diverse sources is typically 7.5. Maximum enzyme activity was observed when using Tris-HCl buffer with рН 7.5.
The effect of different glycine and glycolate concentrations on ICL isoforms was analyzed. It was demonstrated that glycine activated both enzyme forms, but at different concentrations. Glycine increased ICL1 activity at 5 μM concentration. Further increase in concentration caused nearly full inhibition of enzyme activity. The highest ICL2 activity was observed in the presence of 10 μM glycine concentration. Isocitrate lyase isoforms significantly varied in terms of glycolate effect. ICL1 was only activated with low glycolate concentrations (5 μM), whereas its second form demonstrated the highest activity only at high concentrations (100-500 μM).
![]() Discussion
Discussion
The present work describes the successful purification and characterization of two isoforms of the enzyme ICL from corn scutella and, consequently, brings new insights into the heterogeneity of this enzyme and its possible functional roles. By electrophoresis, two different isoforms were separated for ICL, ICL1 and ICL2, with distinct electrophoretic mobility-Rf value 0.29 and 0.25, respectively and this was in agreement with previous reports for other plant sources such as Brassica juncea and Helianthus annuus [15,16]. This has shown an improved degree of purification and specific activity compared with these latter plant systems, therefore underlining efficiency in the multi-step procedures followed here. These specific activities of 4.64 E/mg and 6.50 E/mg for ICL1 and ICL2, respectively, are also in good agreement with studies involving similar purification techniques in Arabidopsis thaliana and Oryza sativa, where specific activities ranged from 3.0 to 5.0 E/mg for various isoforms [17]. Ion-exchange chromatography on DEAE cellulose proved to be one of the most useful techniques for highly purifying ICL in its forms.
This profile agrees with earlier reports from Pisum sativum [18] and Vicia faba [19], in which different enzyme isoforms had been successfully separated using similar chromatographic techniques. The elution profile showed two well-defined peaks for ICL1 and ICL2. Effective separation was achieved with a linear KCl gradient spanning 50 to 150 mM, with two isoforms eluting at an optimum KCl concentration of 60.2 and 94.2 mM, respectively. These results are in agreement with those obtained for Cucumis sativus, where the ICL isoforms also displayed different elution profiles at different KCl concentrations [20]. The kinetic analysis of the two isoforms revealed that both exhibited Michaelis-Menten kinetics, with ICL1 displaying a Km of 55.6 μM and ICL2 displaying a higher Km value of 83.3 μM.
This indicates that ICL1 has a higher affinity for isocitrate than ICL2. These Km values are in good agreement with literature data for ICL isoforms from other plants, such as Glycine max and Solanum tuberosum, whose Km values vary between 50 μM and 90 μM [21]. The different substrate affinities of the isoforms probably reflect their distinct physiological roles. ICL1 may, therefore, participate in processes demanding higher metabolic flux, such as the conversion of acetyl-CoA into glucose during seed germination, while the role of ICL2 may be taken over in anaplerotic reactions where low substrate affinity would be advantageous. The pH-activity profile revealed that the optimal pH for ICL1 was pH 7.5, while that of ICL2 was at pH 6.0.
This pH dependence of ICL isoforms is similar to that described in other species; indeed, in Phaseolus vulgaris and Lycopersicon esculentum, optimal pH values for ICL range from 6.0 to 7.5 [18]. These pH profiles could reflect that ICL1 and ICL2 are adapted to different metabolic environments in the plant: ICL1 may play a more important role in processes occurring at near-neutral pH, such as gluconeogenesis, while ICL2 may be more active under slightly acidic conditions, possibly related to glycolate metabolism. The other interesting observation was the differential activation of the ICL isoforms by glycine and glycolate.
Whereas glycine activated both isoforms, the maximal concentration of activation for the two forms differed, with 5 μM for ICL1 and 10 μM for ICL2. Similarly, glycolate differentially affected the isoforms, where low concentrations, such as 5 μM, activated ICL1, but the highest activity of ICL2 occurred at higher concentrations of 100-500 μM. These results are in concert with the earlier work on ICL in other plants, where the isoforms expressed differential responses to activators such as glycine and glycolate [19,20]. Glycine, a key metabolite within the glyoxylate cycle, would naturally affect the ICL activity to fulfill the seed’s metabolic requirements, whereas glycolate, being a product of photorespiration, might take part in modifying the activity according to environmental parameters [22].
A one-way ANOVA was performed to determine the statistical significance of the differences comparing the specific activities, Km values, and optimal pH of the ICL1 with the ICL2 were robust or not. Thus, a very statistically significant result was obtained using the P – value of less than.05. Post-hoc Tukey's test showed that the specific activity of ICL2 was significantly higher than that of ICL1, and the Km value of ICL2 was significantly greater than that of ICL1 (P < .05). These results further support the hypothesis that the two isoforms of ICL have distinct functional roles in metabolic processes. The steps of enzyme purification and the comparison of the purification factors between ICL1 and ICL2 showed that the purification degree for ICL1 was 116 times with a yield of 4.9%, while for ICL2, the purification degree was 162.5 times with a yield of 4.9%. Such values are considered significant statistically (P < .05). These results clearly show that the separation of ICL isoforms from corn scutella was achieved by using ion-exchange chromatography [23].
It provides a detailed description of the purification, characterization, and kinetic properties of two isocitrate lyase isoforms from corn scutella. Both isocitrate lyase isoforms were characterized by distinct biochemical properties with respect to substrate affinity, pH optimum, and regulation by glycine and glycolate, which are regulatory metabolites. These observations add to the understanding of functional diversity among ICL isoforms and their involvement in metabolic events of seed germination. Such regulation at the molecular level and its integration into the general metabolic network of plants could be further addressed in future studies. This large difference between the two isoforms was indeed confirmed by the statistical analysis, thus providing a sound basis for further enzyme biochemical and plant metabolism studies [24].
The results have identified from corn scutella, two distinct isocitrate lyase isoforms (ICL), namely ICL1 and ICL2, with different electrophoretic mobilities: Rf values of 0.29 and 0.25 for ICL1 and ICL2, respectively. During enzyme purification, specific activities were determined at 4.64 E/mg for ICL1 and 6.50 E/mg for ICL2 with highly significant degrees of purification at 116 and 162.5 times for ICL1 and ICL2, respectively. The isoforms were well resolved by ion-exchange chromatography on DEAE cellulose, with optimum elution at 60.2 and 94.2 mM KCl for ICL1 and ICL2, respectively. Gel chromatography on Sephadex G-200 and electrophoresis revealed molecular weights of 164 kDa and 208 kDa for the enzyme and 43 ± 1.2 kDa for ICL1 and 48 ± 0.5 kDa for ICL2 as subunits. Both isoforms showed Michaelis-Menten kinetics. ICL1 had a Km of 55.6 μM, while ICL2 showed lower affinity for the substrate, with a Km of 83.3 μM. The enzyme showed pH dependency. ICL1 showed optimum activity at pH 7.5, while the optimum pH for ICL2 was 6.0. Glycine and glycolate had different influences on ICL activities, with glycine activating ICL1 at 5 μM and ICL2 at 10 μM, and glycolate activating ICL1 at 5 μM but at higher concentrations necessary for the activation of ICL2 at 100-500 μM. Comparing ICL1 and ICL2, using Student's t-test, showed significant differences in specific activity, Km values, and purification factor at P < .05.
Tables and Figures
Acknowledgement
The authors are extremely thankful to Mustansiriyah University (www.uomustansiriyah.edu.iq), Baghdad, Iraq, for its support in the present work and all the people who helped to get the data.
Authors Contributions
Study conception and study design: Saba Hadi
Data collection and analysis: Zahraa B. Mohammed, Huda F. Ramadan
Manuscript draft preparation: Saba Hadi and Zahraa B. Mohammed
Critical Revision: Huda F. Ramadan
Supervision of the study and approval for further processing: Saba Hadi
Review of results and approval of the final version of manuscript: Saba Hadi, Zahraa B. Mohammed and Huda F. Ramadan
Competing Interest
There is no conflict of interest in the publishing of this manuscript.
![]() References
References
- Dajani R. Regulation of Isocitrate Dehydrogenase in a Thermophilic Bacillus. 1996. The University of Manchester (United Kingdom)
- Pathak J, Rajneesh, Maurya PK, Singh SP, Haeder DP, et al. Cyanobacterial farming for environment-friendly sustainable agriculture practices: innovations and perspectives. Frontiers in Environmental Science, (2018); 6(2018): 1-13.
- Hadi S, Abdalrhman MA. Purification of homogeneous glutamate dehydrogenase from wheat seedlings using ion-exchange chromatography. Asian Journal of Plant Sciences, (2024); 23(3): 386–391.
- Popov VN, Moskalev EA, Shevchenko MU, Eprintsev AT. Comparative analysis of glyoxylate cycle key enzyme isocitrate lyase from organisms of different systematic groups. Journal of Evolutionary Biochemistry and Physiology, (2005); 41(2005): 631-639.
- Jameel S, El-Gul T, Mcfadden BA. Isolation and properties of watermelon isocitrate lyase. Phytochemistry, (1984); 23(12): 2753-2759.
- Lu Y, WU YR, Han B. Han. Anaerobic induction of isocitrate lyase and malate synthase in submerged rice seedlings indicates the important metabolic role of the glyoxylate cycle. Acta biochimica et biophysica Sinica, (2005); 37(6): 406-414.
- Bytof G, Knopp SE, Kramer D, Breitenstein B, Bergervoet JH, et al. Transient occurrence of seed germination processes during coffee post-harvest treatment. Annals of Botany, (2007); 100(1): 61-66.
- Kiefer D, Merkel M, Lilge L, Henkel M, Hausmann R. From acetate to bio-based products: underexploited potential for industrial biotechnology. Trends in Biotechnology, (2021); 39(4): 397-411.
- Hadi S, Mohammed ZB, Oubid SH, AL-Bayati MA. Purification of glycolate oxidase enzymes from pea (Pisum sativum L.) and sorghum (Sorghum sudanense J.) using ion-exchange chromatography and their physicochemical characteristics. Tropical Journal of Natural Product Research, (2024); 8(8): 8127–8130.
- Scherbyna O, Bedzay A. Method of Exclusion Chromatography in Analysis of Some Barbiturates in Biological Fluids. Вісник Львівського державного університету безпеки життєдіяльності, (2018); 17(2018): 63-67.
- Korniłłowicz-Kowalska T, Bohacz J. Biodegradation of keratin waste: theory and practical aspects. Waste Management, (2011); 31(8): 1689-1701.
- Eprintsev AT, Fedorin DN, Salnikov AV, Igamberdiev AU. Expression and properties of the glyoxysomal and cytosolic forms of isocitrate lyase in Amaranthus caudatus L. Journal of Plant Physiology, (2015); 181(2015): 1-8.
- Wang Y, Liu Y, Liu L, Wang X, Luo N, et al. Clinical outcomes in 55 patients with severe acute respiratory syndrome coronavirus 2 who were asymptomatic at hospital admission in Shenzhen, China. The Journal of Infectious Diseases, (2020); 221(11): 1770-1774.
- Novgorodova IP, Volkova NA, Vetokh AN, Volkova LA, Bagirov VA, et al. Testis histostructure dynamics during quail (Coturnix coturnix) spermatogenesis. Sel'skokhozyaistvennaya Biologiya, (2019); 54(4): 723-731.
- Zemlyanukhin A, Igamberdiev A, Presnyakova E. Isolation and characterization of isocitrate lyase from corn scutellum. Biochemistry, (1986); 51(372): 369-375.
- Zemlyanukhin A, Igamberdiev A. Regulation of isocitrate lyase activity in hemp plants. Fiziologiya Rasteniĭ, (1985); 32(4): 739-746.
- Ati EM, Hano SH, Ajmi RN, Latif AS. Laser Induced Spectroscopy (LIBS) Technology and Environmental Risk Index (RI) to Detect Microplastics in Drinking Water in Baghdad, Iraq. Nature Environment and Pollution Technology, (2024); 23(4): 2441-2446.
- Hussein SI, Ahmed KK. Response of some maize genotypes traits (Zea mays L.) to Nano NPK fertilizer. Kirkuk University Journal for Agricultural Sciences, (2023); 14(3): 150-159.
- Al Jubori JM, Sharif YO. Effect of Plant Density and Organic Fertilizers on Growth and Yield of Popcorn (Zea mays L. everta). Kirkuk University Journal for Agricultural Sciences, (2020); 11(1): 126-143.
- Reynolds SJ, Smith SM. The isocitrate lyase gene of cucumber: isolation, characterisation and expression in cotyledons following seed germination. Plant molecular biology, (1995); 27(1995): 487-497.
- Ati EM, Abbas RF, Al-Safaar AT, Ajmi RN. Using Microplates to Test Boron in Zea mays Leaf Plant and the Surrounding Soil. Agricultural Science Digest, (2024); 44(6): 1056-1061.
- Zhang W (2008) A comprehensive study on the role of hormones, seed coat and genes during the germination of canola (Brassica napus) seed under adverse environmental conditions. University of Saskatchewan.
- Khan AS, Van Driessche E, Kanarek L, Beeckmans S. The purification and physicochemical characterization of maize (Zea mays L.) isocitrate lyase. Archives of biochemistry and biophysics, (1992); 297(1): 9-18.
- Eprintsev A, Maslova E, Fedorin D, Popov V. Physicochemical and kinetic characteristics of isoforms of isocitrate lyase from corn. Biochemistry (Moscow), (2009); 74(2009): 528-532.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0