

Full Length Research Article
Genetic Variants of B4GALNT2 are Associated with Twinning in Sheep with Potential to Boost Small Ruminant Populations and Mutton Production
Fouzia Razzaq1, Muhammad Bilal bin Majeed1, Shagufta Naz2, Asad Ali1, Afzal Rashid3, Haiba Kaul1*
Adv. life sci., vol. 12, no. 2, pp. 324-330, May 2025
*– Corresponding Author: Haiba Kaul (Email:haiba.kaul@uvas.edu.pk)
Authors' Affiliations
2. Department of Zoology, Lahore College for Women University, Lahore, 54600 – Pakistan
3. Department of Animal Nutrition, University of Veterinary and Animal Sciences, Ravi Campus, Pattoki, 55300 – Pakistan
[Date Received: 20/10/2024; Date Revised: 05/01/2025; Available Online:22/03/2025; Date Updated: 31/08/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Twinning in sheep is an economically important trait. Of the many genes that can affect litter size in sheep, B4GALNT2 is an important member. Genetic variants found in the B4GALNT2 effect the ovulation and can promote twinning in sheep. Keeping in view the importance of prolificacy in sheep, this research was planned to identify the association of B4GALNT2 gene variants with twining in Lohi sheep.
Methods: A total of 200 blood samples of female sheep (100 each who gave birth to twins and single sheep respectively) were collected from three sheep farms. DNA was extracted and genotyped for B4GALNT2 (rs410464838). For genotyping of SNPs, a PCR-based restriction fragment length polymorphism assay was used. Data were analyzed by using SPSS and R. Hardy-Weinberg equilibrium was evaluated for both groups of sheep. Association of litter size with B4GALNT2 genetic variants was tested through chi square test.
Results: The results of the study showed that genotypes at the B4GALNT2 locus were at Hardy-Weinberg disequilibrium for twin birth. Association analysis revealed a strong association of the AA genotype with twinning. Animals with this genotype were found to be predisposed to having twins. Furthermore, Bayesian analysis reinforced these findings, demonstrating a robust association between the A allele and an increased likelihood of twin births.
Conclusion: Overall, the results of this study indicate that the B4GALNT gene is a good candidate for selection of litter size in sheep. If incorporated as part of a marker assisted selection program, it can result in a sustained expansion of small ruminants to increase mutton production and uplift the socioeconomic condition of small ruminant farmers.
Keywords: Twinning; Prolificacy; B4GALNT2; Sheep; Association
Introduction![]()
Twinning, or the birth of twin lambs, is a highly desirable trait in sheep (Ovis aries) farming due to its numerous benefits. Ewes that have twins rather than single lambs are more likely to be profitable for farmers. This is because twin births allow for increased lamb production from the same number of ewes [1]. The sheep is a household or domesticated species that is raised in agricultural surroundings for meat, wool, hair, leather and also a source of employment in the rural sector. Demand for mutton is rapidly increasing in developing countries, with estimates suggesting total meat consumption will reach 326 Mt per year by 2050 in these regions [2]. However, production is struggling to keep pace with this growing demand, leading to high prices that make mutton inaccessible for many low-income consumers. One key strategy for boosting mutton supply is improving reproductive traits like litter size. Increasing average litter size could significantly improve profitability for sheep farmers [3].
Twinning has a genetic component, and studies have identified several genetic factors associated with twinning in sheep, focusing particularly on genes related to fertility and reproductive traits. Research indicates that the ovulation rate and litter size in sheep are genetically controlled by a combination of genes with minor effects (polygenic component) and their interaction with environment [4].
Notably, the growth differentiation factor 9 (GDF9) gene has been linked to ovarian function and fertility, with specific polymorphisms correlating with higher lambing rates in certain breeds, particularly among Egyptian sheep [5]. The Booroola fecundity gene (FecB) is another significant contributor, associated with increased ovulation rates and prolificacy in breeds like Garole and Booroola [6]. Recent genome-wide association studies (GWAS) have uncovered multiple single nucleotide polymorphisms (SNPs) related to reproductive traits, indicating that genetic selection could effectively enhance twinning rates while considering the interplay of environmental factors such as maternal age and health [7]. However, monogenic inheritance has also been reported, particularly with the expression of the fecundity (Fec) gene, which has a large effect on litter size [8-9]. Major genes influencing litter size include FecB, FecX, bone morphogenetic protein-15 (BMP15), GDF9, and BMP receptor-1B (BMPR1B) [7-9].
A promising candidate gene, Beta-1, 4-N-acetyl-galactosaminyltransferase 2 known as B4GALNT2 for fecundity in sheep was identified within the FecL locus in French Lacaune sheep [10]. The ectopic expression of the B4GALNT2 gene plays a key role in regulating ovulation rates and twinning in sheep. Specific genetic variants like those in the B4GALNT2 gene, as well as more general factors like heritability and major genes for prolificacy, all contribute to the genetic basis of twinning in sheep. Selective breeding targeting these genetic factors can be an effective way to increase twinning rates in sheep flocks.
Selecting ewes and rams from prolific lines can increase the likelihood of twinning in future generations. Some breeds of sheep are more prolific than the others. However, through marker assisted selection in prolific genes, litter size can be increased in any sheep breed like indigenous sheep breeds of Pakistan like Lohi. Lohi is the most popular breed and is economically important that is used for mutton and wool [11]. It is also sacrificed during Eid. Twinning has been observed in sheep, and a twinning rate of 33% has been reported in Lohi sheep [11].
Therefore, this study was designed to identify molecular markers that influence fecundity in our local sheep breed such as Lohi, Thus, marker assisted selection in sheep will enable sheep breeder to choose the most productive animals to mate, based on evidence of genetic markers related to reproductive traits. It is expected to result in an increase in the number of sheep for local farmers.
Methods![]()
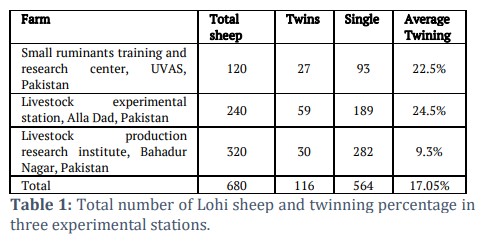
The study was conducted after ethical approval from the institution Animal Care and Use Committee (ACUC) of the University of Veterinary and Animal Sciences, Pakistan. Phenotypic data was collected from the lambing records of three sheep farms i.e., Small Ruminants Training and Research Centre (SRT & RC), UVAS, Livestock Experiment Station Allah Dad, Khanewal, and Livestock Production Research Institute, Bahadur Nagar, Okara, Pakistan. All the three flocks were on semi-intensive production system and homogeneous age and parity sample was selected. Across the farms housing 680 Lohi sheep, comprehensive data on lambing, ewe age at lambing, production systems, and birth type (single or multiple) were recorded. Out of this population, 200 ewes were specifically chosen for molecular analysis, with blood samples collected accordingly. These ewes were subsequently categorized into two distinct groups based on their lambing history: Group A comprised ewes that had previously given birth to singles (n=100), while Group B consisted of ewes that had previously delivered twins (n=100). The selection aimed for a homogeneous sample in terms of age and parity, which is easier to achieve within a single breed like Lohi.
The genetic variants in B4GALNT2 (rs410464838) gene were analyzed and were checked for their association with litter size. A total of 5 ml of blood sample was collected out of each ewe in 0.5 M EDTA (pH: 8) and were stored at -20°C. DNA was extracted using a modified phenol-chloroform protocol [13,14]. Primer pairs for the amplification of the genomic region were designed using Primer 3 software [14] and B4GALNT2 was genotyped by PCR-RFLP method. Forward primer 5’ – TCTTAAATTGCCCTCAGTCC -3’and reverse primer 5’- AAGAACTCTGCCAAGAGAGG -3’; were used for the study. The PCR was done in 25 μl reaction volume containing 50 ng DNA template, 10 pmol forward and reverse primers, 0.4 mM each dNTP, 2.5 mM MgCl2, 1X Taq buffer and 1 U of Taq DNA polymerase (Thermo Scientific). It was then subjected to thermocycling conditions consisting of 4 minutes of denaturing at 94°C followed by 30 cycles of amplification each containing three steps. The denaturation was carried out at 94°C for 30 seconds, annealing at 58°C for 30 seconds and an extension at 72°C for 45 seconds and lastly final extension of 5 minutes at 72°C. The amplicons formed as a result of PCR were analyzed on 1.5% agarose gel.
PCR-RFLP method was used for genotyping of B4GALNT2 SNPs. After amplification, the 240-base pair (bp) PCR product was digested with restriction enzyme AvaII (Thermo Scientific). Samples were then incubated at 37°C for 16 hours for complete digestion. Digestion of the B4GALNT2 allele yielded fragments of 240 bp for GG genotype, 240, 163 and 77 bp for AG genotype, and 163, and 77 bp for AA genotype. The digested products were separated by 4% agarose gel electrophoresis. Wild type GG and variant alleles were differentiated from each other based upon the different migration profiles of digested DNA fragments.
Selected DNA samples were sequenced to validate the results of RFLP. They were later used as controls in analyzing the RFLP data. DNA sequencing was done commercially by automated ABI PRISM® 3730 Genetic Analyzer. The sequencing results were later analyzed using Chromas software. The intronic variant was analyzed using Ensembl Variant Effect Predictor (VEP) and Global minor allele frequency (GMAF).
The data was entered and analyzed using SAS software. Genotypic and allelic frequencies were calculated. Having estimated the allele frequency in each group, the expected frequencies of the AA, AG and GG genotypes were calculated under the assumption of Hardy-Weinberg Equilibrium (HWE). The data were entered and analyzed using SAS software, where genotypic and allelic frequencies were calculated to assess the genetic diversity within the populations. After estimating the allele frequency in each group, the expected frequencies of the AA, AG, and GG genotypes were computed under the assumption of Hardy-Weinberg Equilibrium (HWE). This assumption is fundamental in population genetics as it provides a baseline to evaluate whether observed genotype frequencies deviate from expected distributions due to evolutionary forces.
Chi-square tests were used to find the association between genotypes and litter size, allowing for a clear determination of whether specific genotypes are linked to reproductive outcomes. Observed and expected frequencies were then compared using an exact goodness of fit test using the Xmulti function in the XNomial package in R. This approach is particularly useful for small sample sizes or when expected frequencies are low, ensuring robust results. This was done separately for twin and single lambing data, and this allowed assessment if each group was in HWE. In addition, observed genotype frequencies between twin and single samples were compared using a Fisher's exact test, through the Fisher test function in R. This method is advantageous for analyzing categorical data with small sample sizes, providing precise p-values for evaluating associations between genotype distribution and lambing outcomes.
To check for false positives in the association between the B4GALNT2 genotypes and lambing outcomes, a Bayesian logistic regression model was employed. This approach allowed for the incorporation of prior knowledge about the relationship between alleles and reproductive traits, enhancing the robustness of the results. The model utilized Markov Chain Monte Carlo (MCMC) methods to estimate posterior distributions for the parameters, providing a probabilistic framework to assess the likelihood of true associations while controlling for false positives. Together, these statistical methods offer a comprehensive framework for understanding the genetic influences on litter size in Lohi sheep.
Results![]()
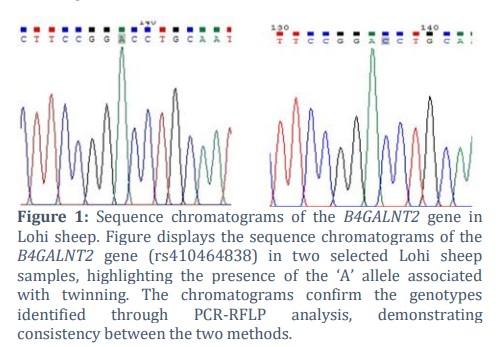
Out of 680 Lohi sheep in three experimental stations, 116 of them gave birth to twins and 564 produced single births. The average twinning rate in our study was 17.05%, as tabulated in Table 1. Of the 680 sheep, 200 samples were selected for further studies. All the selected sheep were of 2-4 years of age, disease free and in second or third parity. Semi-intensive production system was used in all three farms. Of the 116 sheep that gave birth to twins, 24 were themselves born as twins. Data about the 38 of the sheep that gave twin birth was not available. This data highlights the variability in twinning rates across different experimental stations, with the overall average twinning percentage at 17.05%. For molecular analysis, DNA was extracted, and primers were optimized at 58°C with 2.5mM MgCl2. After amplification, the 240-base pair (bp) PCR product was digested using restriction enzymes AvaII. Digestion of the B4GALNT2 ‘A’ allele yielded fragments of 163 and 77 bp while digestion of ‘G’ allele yielded a 240 bp fragment. Sequencing of the selected samples confirmed the genotypes detected by PCR-RFLP. About 10 samples were sequenced and the results showed the same genotype of B4GALNT2 as it was detected by PCR-RFLP. Sequencing results of the two samples are depicted in figure 1.
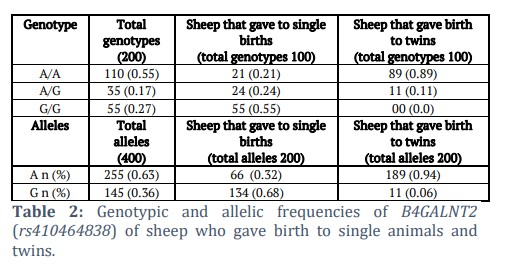
An analysis of B4GALNT2 genotypes in Lohi sheep revealed distinct allele frequencies associated with lambing outcomes. Ewes that gave single births exhibited a G allele frequency of 0.67, while the A allele was present at a frequency of 0.33. In contrast, ewes that gave birth to twins showed a significantly higher frequency of the A allele at 0.94, with the G allele being rare at 0.06.
The genotype frequencies for twin births were AA: 0.89, AG: 0.11, and GG: 0.00. Results are depicted in table 2.
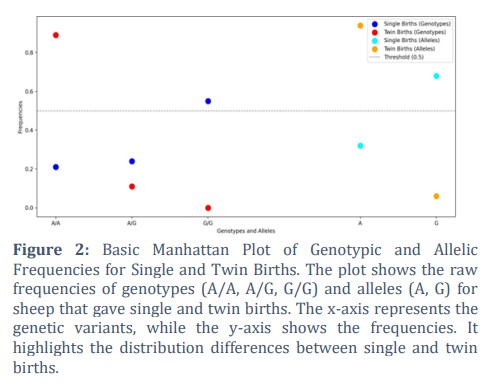
Hardy-Weinberg equilibrium (HWE) tests indicated that the single birth sample was not in HWE (χ² = 14.51, df = 2, P = 0.0007), whereas the twin birth sample conformed to HWE (χ² = 0.20, df = 2, P = 0.90). A Fisher's exact test revealed a significant difference in genotype distribution between the two groups (P = 5.7×10−14), confirming that the A allele frequency is significantly higher in ewes with twins compared to those with single births. The allelic and genotypic frequencies are depicted as Manhattan plot in figure 2.
The results of Bayesian analysis indicated a strong association between the A allele of the B4GALNT2 locus and an increased likelihood of twin births, with a posterior mean for the genotype coefficient of β1=3.45 (95% credible interval: 2.10 to 4.80).. This method demonstrated improved control of false positives compared to traditional approaches, as evidenced by lower rates of incorrect identification of non-significant variables. The results suggest that ewes carrying the A allele had approximately a 94% probability of giving birth to twins, reinforcing the potential of allele A as a genetic marker for improving litter size in Lohi sheep.
However, this study has limitations, including a relatively small sample size and potential biases due to population structure and environmental influences not accounted for in the analysis. Future research should aim to include larger and more diverse populations while controlling these factors to validate these preliminary findings and explore the underlying biological mechanisms further.
Figures & Tables
Ovulation rate is the major source of variation governing multiple births in sheep. Genetic variants in B4GALNT2 affect the ovulation rate [10] and identification of such markers can help in devising a successful selection program for litter size in sheep. Although there are genetic variations reported in different genes with major effects on reproductive performance traits, such as ovulation rate and litter size in different sheep breeds around the world [6,15-16], the effect of environmental factors like age, parity, nutrition, body weight and season cannot be denied. Adequate nutrition is essential for optimal reproductive health, as deficiencies can lead to poor body condition and reduced fertility, while a balanced diet enhances ovulation and litter sizes. Environmental factors, such as temperature and housing conditions, can also affect reproductive performance by causing stress that disrupts hormonal balance. Estimates of heritability regarding prolificacy in sheep varies across different sheep breeds but it is commonly regarded as a complex trait with many genetic loci and environmental factors influencing the litter size [17]. In studies where large heritability was found for the mean prolificacy, with high percentage of twins at lambing environmental variability was greatly reduced [18]. So, we reduced the non-genetic factors as possible to investigate the association of genetic variants in the B4GALNT2 gene with litter size in Lohi sheep. Lohi is a local mutton breed, as for a successful improvement program, it is essential to evaluate indigenous breeds as well [19].
Our study reports a mean twinning percentage of 17.05% while the highest twinning was 24.5% in one of the farms. Previously, the average twinning percentage in Lohi sheep was reported to be 33% by Ahmad and colleagues. This might be due to the genetic causes that remain to be validated. In order to correlate this phenotype with molecular cause, allelic and genotypic frequencies of B4GALNT2 were estimated and the association of the litter size with B4GALNT2 was determined.
A difference of genotype and allele frequencies was observed between ewes with twin births and those with single births. B4GALNT2 genetic variant (rs410464838) resides on chromosome 11 sheep and is biallelic (A/G). This variation is present in the flanking region of the last exon of B4GALNT2. This is a transcript variant occurring within an intron of a coding transcript near the splice site. As mutations in B4GALNT2 enhance gene expression that can result in multiple ovulation, this intronic variant can affect transcription levels, and transcript stability [20,21]. Moreover, intronic variants can also increase the efficiency of mRNA translation [22,23]. However, functional studies can assess its importance in gene regulation. Allele ‘A’ was markedly frequent in animals that gave birth to twins while ‘G’ allele was common in the other group. In animals that gave birth to single animals, GG genotype was the most frequent with a frequency of 0.55 and a corresponding G allele frequency of 0.68 (and A allele frequency of 0.32). In contrast, in the group of ewes that gave birth to twins, AA genotype was the most common (genotype frequency of 0.89) with a corresponding allele frequency of 0.94; there were no homozygous GG sheep observed. The results showed a potential association of the A allele with twinning in Lohi sheep. In addition to this, observed genotype frequencies for twin and single samples were compared using a Fisher exact test in R. This comprehensive analytical approach aimed to discern potential associations between the B4GALNT2 variant genotypes and litter size, ensuring a robust examination of genetic influences in both twin and single datasets. The chi-square value was 31.403 (P < .0001), which showed a significant association between genotypes and litter size.
In our study, we noticed that the single birth frequencies were not in Hardy-Weinberg equilibrium (HWE) that can be attributed to several potential factors. Firstly, HWE assumes a large, randomly mating population with no evolutionary influences such as natural selection, genetic drift, mutation, or gene flow. If any of these conditions are violated, deviations from HWE can occur. For example, if there is non-random mating—such as inbreeding or assortative mating, this can lead to an increase in homozygosity and affect allele frequencies. However, due to the difference of genotypic and allelic frequencies in both groups ‘A allele’ of B4GALNT2 gene can be considered as a potential marker for fecundity in Lohi sheep. However, functional studies and testing of this polymorphism in large samples are required to validate the results. Previously mutation in B4GALNT2 has been found to be strongly associated with litter size in ten breeds of sheep [24,26] However, this study was conducted on 200 animals (100 of each group) and therefore studies on larger groups of animals are required to validate the results of the study and investigate the effect across different parities. It is also important to validate this association on different populations of Lohi sheep before implementation in an MAS program. From an operational point of view, there are many challenges that need to be overcome to incorporate this marker as part of a MAS program. Likely, it would be part of a selection program that would select for more than one gene and more than one trait. Further, any selection would need to be made to ensure that the resultant ‘improved breed’ is appropriate for the environment in which it is going to be raised.
Future research should focus on conducting functional studies to confirm the specific biological mechanisms by which B4GALNT2 influences twinning rates in sheep. This could involve gene knockout or overexpression experiments to observe the effects on fertility. Additionally, expanding the investigation across diverse sheep populations will help assess the prevalence and impact of different alleles in various environmental contexts. Longitudinal studies tracking reproductive performance over multiple breeding seasons could further elucidate the long-term effects of B4GALNT2 variants on litter size, ultimately contributing to more effective genetic selection strategies.
Author Contributions
Haiba Kaul conceptualized and supervised the study, Asad Ali and Afzal Rashid were also involved in supervision, Fouzia Razaq collected data and performed experiments. Bilal Bin Majeed helped in data analysis and DNA sequencing and Shagufta Naz helped in statistical analysis and was involved in proofreading the manuscript.
Acknowledgement
Authors are thankful to the farm managers, all staff members of three experimental stations who provided access to data and samples (blood) of Lohi sheep. Authors especially thank Prof. Dr. Khalid Javed (retd.) for his guidance, support and valuable suggestions in conducting this research work.
The authors declare that there is no conflict of interest regarding the publication of this paper.![]()
References
- Haile A, Getachew T, Mirkena T, Duguma G, Gizaw S, et al. Community-based sheep breeding programs generated substantial genetic gains and socioeconomic benefits. Animal, (2020); 14(7):1362-1370.
- Thornton PK. Livestock production: recent trends, future prospects. Philosophical Transactions of the Royal Society B: Biological Sciences, (2010); 365(1554): 2853-2867.
- Zhou M, Pan Z, Cao X, Guo X, He X, et al. Single nucleotide polymorphisms in the HIRA gene affect litter size in Small Tail Han Sheep. Animals, (2018); 8(5): 71
- Khazaal NM, Alghetaa HF, Al-Shuhaib MB, Al-Thuwaini TM, Alkhammas AH. A novel deleterious oxytocin variant is associated with the lower twinning ratio in Awassi ewes. Animal Biotechnology, (2023); 34(8): 3404-3415.
- El Fiky ZA, Hassan GM, Nassar MI. Genetic polymorphism of growth differentiation factor 9 (GDF9) gene related to fecundity in two Egyptian sheep breeds. Journal of Assisted Reproduction and Genetics. (2017); 34(12): 1683-1690.
- Zongsheng ZH, Heng YA, Yaosheng YU, Yifan XI, Manjun ZH, Liang H. Identification of SNP within the sheep RXRG gene and its relationship with twinning trait in sheep. Kafkas Universitesi Veteriner Fakultesi Dergisi, (2018); 24(1): 39-43.
- Jia JL, Xie DJ, Zhang YY, Zhang HX, Zhang LP, et al. Association of BMPR-1B gene 3"-UTR region polymorphism with litter size in Tibetan sheep. Kafkas Universitesi Veteriner Fakultesi Dergisi, (2021); 27(3): 285-289.
- Chantepie L, Bodin L, Sarry J, Woloszyn F, Ruesche J, et al. Presence of causative mutations affecting prolificacy in the Noire du Velay and Mouton Vendéen sheep breeds. Livestock Science, (2018); 216: 44-50.
- Drouilhet L, Lecerf F, Bodin L, Fabre S, Mulsant P. Fine mapping of the FecL locus influencing prolificacy in Lacaune sheep. Animal Genetics, (2009); 40: 804–812.
- Drouilhet L, Mansanet C, Sarry J, Tabet K, Bardou P, et al. The highly prolific phenotype of Lacaune sheep is associated with an ectopic expression of the B4GALNT2 gene within the ovary. PLoS Genetics, (2013); 9(9): e1003809.
- Ahmad Z, Yaqoob M, Younas M. The Lohi sheep: a meat breed of Pakistan review. Pakistan Journal of Agricultural Sciences, (2001); 38(3-4): 69-72.
- Grimberg J, Nawoschik S, Belluscio L, McKee R, Turck A, Eisenberg A. A simple and efficient non-organic procedure for the isolation of genomic DNA from blood. Nucleic acids research, (1989); 17(20): 8390.
- Kaul H, Riazuddin SA, Shahid M, Kousar S, Butt NH, et al. Autosomal recessive congenital cataract linked to EPHA2 in a consanguineous Pakistani family. Molecular vision, (2010); 16(58-59): 511-517
- Untergasser A, Cutcutache I, Koressaar T, Ye J, Faircloth BC, et al. Primer3–new capabilities and interfaces. Nucleic Acids Res, (2012); 40(15): e115.
- Xu SS, Gao L, Xie XL, Ren YL, Shen ZQ, et al. Genome-wide association analyses highlight the potential for different genetic mechanisms for litter size among sheep breeds. Frontiers in genetics, (2018); 10(9): 118.
- Akhatayeva Z, Mao C, Jiang F, Pan C, Lin C, et al. Indel variants within the PRL and GHR genes associated with sheep litter size. Reproduction in Domestic Animals, (2020); 55(11): 1470-1478.
- Murphy TW, Keele JW, Freking BA. Genetic and nongenetic factors influencing ewe prolificacy and lamb body weight in a closed Romanov flock. Journal of Animal Science, (2020); 98(9): skaa283.
- SanCristobal-Gaudy M, Bodin L, Elsen JM, Chevalet C. Genetic components of litter size variability in sheep. Genetics Selection Evolution, (2001); 33(3): 249-271.
- Sharif N, Ali A, Dawood M, Khan MI, Do DN. Environmental effects and genetic parameters for growth traits of Lohi sheep. Animals, (2022); 12(24): 3590.
- Xu L, Chen Z, Chen S, Chen Y, Guo J, et al. An Identification of Functional Genetic Variants in B4GALNT2 Gene and Their Association with Growth Traits in Goats. Genes, (2024); 15(3): 330.
- Park JS, Woo SJ, Song CS, Han JY. Modification of surface glycan by expression of beta-1, 4-N-acetyl-galactosaminyltransferase (B4GALNT2) confers resistance to multiple viruses infection in chicken fibroblast cell. Frontiers in Veterinary Science, (2023);10: 1160600.
- Rogalska ME, Vivori C, Valcárcel J. Regulation of pre-mRNA splicing: roles in physiology and disease, and therapeutic prospects. Nature Reviews Genetics, (2023); 24(4): 251-269.
- Ji X, Cao Z, Hao Q, He M, Cang M, et al. Effects of New Mutations in BMPRIB, GDF9, BMP15, LEPR, and B4GALNT2 Genes on Litter Size in Sheep. Veterinary Sciences, (2023); 10(4): 258.
- Guo X, Wang X, Liang B, Di R, Liu Q, et al. Molecular cloning of the B4GALNT2 gene and its single nucleotide polymorphisms association with litter size in small tail han sheep. Animals, (2018); 8(10): 160.
- Nenova R, Dimitrova I, Stancheva N, Bozhilova-Sakova M, Tzonev T, Minkova T. Genetic markers associated to improving prolificacy of sheep. A review. Bulgarian Journal of Agricultural Science, (2023); 29(2): 371–377.
- Getaneh M, Taye M, Alemayehu K, Haile A, Getachew T, Ayalew W. A review on candidate genes associated with sheep fertility traits: Implications for genetic improvement of indigenous sheep breeds in developing countries. Ecological Genetics and Genomics, (2024); 14: 100243.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0