

Short Communication
The prevalence of antimicrobial-producing Gram-positive bacteria in human gut: a preliminary study
Arif Luqman1*, Nimerta Kumari2, Jongkon Saising3, Aparna Viswanathan Ammanath4, Nur Hidayatul Alami1, Endry Nugroho Prasetyo1, Enny Zulaika1, Maya Shovitri1, Nengah Dwianita Kuswytasari1, Tri Haidar Munif1, Friedrich Götz4
Adv. life sci., vol. 10, no. 1, pp. 01-04, March 2023
*– Corresponding Author: Arif Luqman (Email: arif.luqman@its.ac.id)
Authors' Affiliations
2. Institute of Microbiology, University of Sindh, Jamshoro – Pakistan
3. School of Health Science and Biomedical Technology Research Group for Vulnerable Populations, Mae Fah Luang University, Chiang Rai – Thailand
4. Microbial Genetics, Interfaculty Institute of Microbiology and Infection Medicine Tübingen (IMIT), University of Tübingen, Tübingen – Germany
[Date Received: 07/11/2022; Date Revised: 09/02/2023; Date Published Online: 31/03/2023; Date Updated: 05/09/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Human gut microbiome is an excellent source for searching novel antimicrobials which is currently in need due to the rise of drug-resistance bacteria. Many Gram-positive bacteria isolated from human gut have been reported to produce antimicrobial compounds but still only a few studies investigate the prevalence of these bacteria in human gut.
Methods: We took stool samples from 19 adult participants (age: 20–70 years; ethnicity: European and Asian). Stool samples obtained from 7 females and 12 males. We screened for Gram-positive antimicrobial-producing bacteria from the stool samples and identified the positive ones using 16S rRNA sequencing.
Results: Here, we reported that antimicrobial-producing Gram-positive bacteria can be found in the stool samples of 6 out of 19 participants. By screening against Staphylococcus aureus USA300 and Pseudomonas aeruginosa PAO1, some isolates exhibited a different inhibition activity compared to the previously reported antimicrobial compounds.
Conclusion: Our findings showed that some strains isolated from human gut exhibit novel antimicrobial activity which implies that there could still be novel antimicrobial compounds in human gut produced by Gram-positive bacteria.
Keywords: Antimicrobial-producing bacteria; Gram-positive bacteria; Gut microbiota; Microorganism
Introduction![]()
Human gut microbiome, due to its complexity, its pivotal contribution to human physiology and development [1-3], and its unravelled mystery, is becoming a hotspot for researchers to mine resources for solving problems that human is facing. Particular problem that is arising is the need of novel antimicrobial to cope with the drug-resistance bacteria, which become a major threat. The human gut microbiome can be an excellent source for researchers to identify novel antimicrobial compounds as it also plays a crucial role in protection against pathogen invasion [4,5], particularly in gastrointestinal tract [6-8].
Many Gram-positive bacteria isolated from the human gut have been reported to produce antimicrobial compounds, namely Bacillus, Ruminococcus, Enterococcus, Lactobacillus, etc. [9]. However, there are still few studies investigating the prevalence of antimicrobial-producing Gram-positive bacteria in human gut. In this study, we investigated the prevalence of these bacteria and identified them using 16S rRNA.
Methods![]()
Stool Sample Collection
The use of human stool samples was approved by the Ethics Commission of the University of Tübingen (approval no. 320/2017BO2). Stool samples were obtained from 19 adult participants (age: 20–70 years; ethnicity: European and Asian; location: Tuebingen and Stuttgart, Germany). Stool samples were obtained from 7 female and 12 male participants.
Screening for Antimicrobial Activity
The obtained stool samples were diluted and spread on modified SK medium with lower sodium azide (0.25%) [10]. After incubation at 37°C for 48 h, the obtained colonies were randomly picked and cultured in TSB medium in 96-well plates at 37°C for 24 h. The supernatants were harvested, and 100 µL of each was mixed with 100 µL of freshly inoculated TSB medium (OD600 = 0.01) containing Staphylococcus aureus USA300 or Pseudomonas aeruginosa PAO1, which had been grown as precultures the previous day. The mixtures were incubated in 96-well plates for 24 h at 37 °C. OD600 of each mixture was then measured to observe the growth of S. aureus USA300 or P. aeruginosa PAO1.
Bacterial Isolates Identification
The isolates that showed inhibition against S. aureus USA300 or P. aeruginosa PAO1 were identified by sequencing the 16S rRNA gene using primers 27F and 1392R [11-13]. The corresponding sequences were then analysed using BLASTN (NCBI).
Results![]()
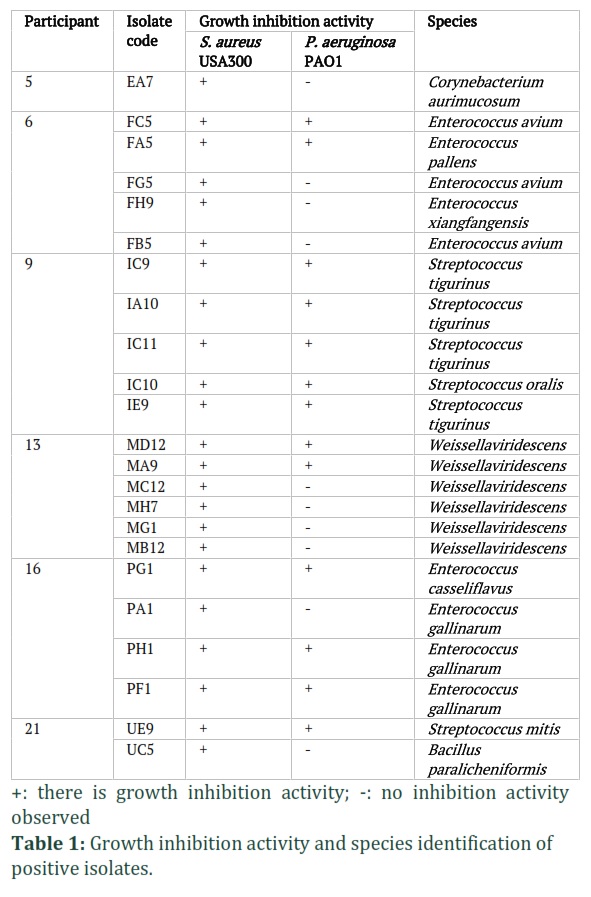
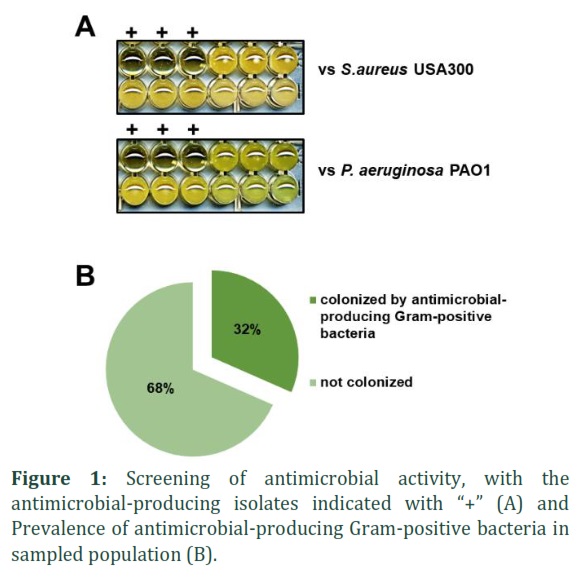
We have examined the stool samples collected from 19 participants by performing the screening of the antimicrobial activity of the grown isolates in modified SK agar medium. The SK agar medium was purposely formulated as a staphylococcal selective medium. We lowered the sodium azide concentration and successfully used it to grow Gram-positive bacteria on the agar. We picked 100 colonies from each participant and carried out the screening (Fig. 1A). We observed there were 23 isolates that showed the growth inhibition activity against S. aureus USA300 and only 13 of them showed the growth inhibition activity towards P. aeruginosa PAO1 as well. We did not find any isolate that showed growth inhibition activity towards P. aeruginosa PAO1 only (Table 1).
Out of the 19 participants, only 6 of them had bacteria that produced antimicrobial substances, and these bacteria were isolated from those individuals. This means that less than half of the participants were colonized by antimicrobial-producing Gram-positive bacteria, as shown in Figure 1B. Among 6 of the antimicrobial-producing Gram-positive bacteria-harboring participants, only 5 of them were colonized by isolates that showed growth inhibition activity towards P. aeruginosa PAO1 (26%). Species identification of the 23 isolates revealed that those showing antimicrobial activity belonged to genera previously reported to produce antibacterial compounds, including Streptococcus, Bacillus, Corynebacterium, Enterococcus, and Weissella (Table 1).
Figures & Tables
The fact that there are bacteria that are able to produce antimicrobial compounds as a gut commensal supports the findings that one of the main functions of commensal bacteria in the gut microbiome is to prevent the pathogenic bacteria to colonize the human gut and control their population [14-16]. With the antimicrobial compounds produced, these bacteria play a key role in maintaining the gut microbiome stability, resistance, and resilience against the pathogenic bacteria invasion. This implies that these bacteria crucially participate in maintaining gut health [17-19].
In this study, we identified some antimicrobial-producing bacteria from various genera. In genera Streptococcus, we identified 3 species: S. tigurinus, S. mitis, and S. oralis. All streptococcal isolates exhibited growth inhibition activity against both S. aureus USA300 and P. aeruginosa PAO1. The possible antimicrobial compounds produced by these isolates is viridins. Viridins were reported to be produced by streptococci, particularly Streptococcus sanguis and S. mitis. This compound is active against Gram-negative and Gram-positive bacteria [20], which is in agreement with our results that streptococcal isolates showed inhibition activity against both S. aureus USA300 and P. aeruginosa PAO1.
For the genera Bacillus, Corynebacterium, and Weissella, we only detected 1 species for each: B. paralicheniformis, C. aurimucosum, and W. viridescens.B. paralicheniformis has been reported to be able to produce formicin, a lantibiotic that is active against Gram-positive bacteria[21], a similar phenotype was shown by the UC5 isolate. C. aurimucosum has not been specifically reported to produce any antimicrobial compound. However, some studies reported Corynebacterium hydrocarboclastusproduces corynecin, a chloramphenicol analog that is active against Gram-positive and Gram-negative bacteria[22,23].Weissella, a lactic acid bacterium, particularly species W. confusa is known to produce some bacteriocins called weissellicin and reported to inhibit the growth of namely Bacillus cereus, Escherichia coli, P. aeruginosa and Micrococcus luteus [24]. However, a novel bacteriocin may have been produced, since the spectra of isolates MC12, MH7, MG1, and MB12 were active only against S. aureus USA300.
Enterococcal bacteria have been known to produce bacteriocins, such as enterocins, that are active against Gram-positive bacteria [25-28]. We observed similar activity for isolate FG5, FH9, and FB5 but not for FC5 and FA5, which might indicate a different and unreported antimicrobial compounds could be produced.
The discovery of isolates with inhibition activity distinct from known antimicrobial compounds such as viridin, corynecin, weissellicin, and enterocin suggests the presence of yet undiscovered and uncharacterized antimicrobial compounds. This opens up the possibility for researchers to uncover new antimicrobial substances from human gut commensal bacteria and develop novel therapeutic approaches for infections by utilizing these bacteria as probiotics.
The authors declare that there is no conflict of interest.
Author Contributions
AL and FG conceived the idea. AL, NK, JS, NHA, ENP, EZ, MS, and NDK designed the experiments. AL, AVA, and THM performed all the experiments. All authors contributed to the article and approved the submitted version.
![]()
Acknowledgement
This research is funded by Institut Teknologi Sepuluh Nopember Directorate of Research and Community Service with the contract number 1676/PKS/ITS/2022.
![]()
References
- Umu OC, Oostindjer M, Pope PB, Svihus B, Egelandsdal B, et al. Potential applications of gut microbiota to control human physiology. Antonie Van Leeuwenhoek, (2013); 104(5): 609-618.
- Walsh CJ, Guinane CM, O'Toole PW, Cotter PD. Beneficial modulation of the gut microbiota. FEBS Letters, (2014); 588(22): 4120-4130.
- Sommer F, Backhed F. The gut microbiota–masters of host development and physiology. Nature Reviews Microbiology, (2013); 11(4): 227-238.
- Ubeda C, Djukovic A, Isaac S. Roles of the intestinal microbiota in pathogen protection. Clinical and Translational Immunology, (2017); 6(2): e128.
- Reinoso Webb C, Koboziev I, Furr KL, Grisham MB. Protective and pro-inflammatory roles of intestinal bacteria. Pathophysiology, (2016); 23(2): 67-80.
- Campana R, van Hemert S, Baffone W. Strain-specific probiotic properties of lactic acid bacteria and their interference with human intestinal pathogens invasion. Gut Pathogens, (2017); 912.
- Iacob S, Iacob DG, Luminos LM. Intestinal Microbiota as a Host Defense Mechanism to Infectious Threats. Frontiers in Microbiology, (2018); 93328.
- Wang L, Zhu L, Qin S. Gut Microbiota Modulation on Intestinal Mucosal Adaptive Immunity. Journal of Immunology Research, (2019); 20194735040.
- Garcia-Gutierrez E, Mayer MJ, Cotter PD, Narbad A. Gut microbiota as a source of novel antimicrobials. Gut Microbes, (2019); 10(1): 1-21.
- Schleifer K-H, Krämer E. Selective Medium for Isolating Staphylococci. Zentralblatt für Bakteriologie: I Abt Originale C: Allgemeine, angewandte und ökologische Mikrobiologie, (1980); 1(3): 270-280.
- Marchesi JR, Sato T, Weightman AJ, Martin TA, Fry JC, et al. Design and evaluation of useful bacterium-specific PCR primers that amplify genes coding for bacterial 16S rRNA. Applied and Environmental Microbiology, (1998); 64(2): 795-799.
- Luqman A, Nega M, Nguyen MT, Ebner P, Gotz F. SadA-Expressing Staphylococci in the Human Gut Show Increased Cell Adherence and Internalization. Cell Reports, (2018); 22(2): 535-545.
- Luqman A, Muttaqin MZ, Yulaipi S, Ebner P, Matsuo M, et al. Trace amines produced by skin bacteria accelerate wound healing in mice. Communications Biology, (2020); 3(1): 277.
- Kamada N, Chen GY, Inohara N, Nunez G. Control of pathogens and pathobionts by the gut microbiota. Nature Immunology, (2013); 14(7): 685-690.
- Kamada N, Seo SU, Chen GY, Nunez G. Role of the gut microbiota in immunity and inflammatory disease. Nature Reviews Immunology, (2013); 13(5): 321-335.
- Pickard JM, Zeng MY, Caruso R, Nunez G. Gut microbiota: Role in pathogen colonization, immune responses, and inflammatory disease. Immunological Reviews, (2017); 279(1): 70-89.
- Jandhyala SM, Talukdar R, Subramanyam C, Vuyyuru H, Sasikala M, et al. Role of the normal gut microbiota. World Journal of Gastroenterology, (2015); 21(29): 8787-8803.
- Scott KP, Antoine JM, Midtvedt T, van Hemert S. Manipulating the gut microbiota to maintain health and treat disease. Microbial Ecology in Health and Disease (2015); 2625877.
- Marchesi JR, Adams DH, Fava F, Hermes GD, Hirschfield GM, et al. The gut microbiota and host health: a new clinical frontier. Gut, (2016); 65(2): 330-339.
- Dajani AS, Tom MC, Law DJ. Viridins, bacteriocins of alpha-hemolytic streptococci: isolation, characterization, and partial purification. Antimicrobial Agents and Chemotherapy, (1976); 9(1): 81-88.
- Collins FWJ, O'Connor PM, O'Sullivan O, Rea MC, Hill C, et al. Formicin – a novel broad-spectrum two-component lantibiotic produced by Bacillus paralicheniformis APC 1576. Microbiology, (2016); 162(9): 1662-1671.
- Nakano H, Tomita F, Yamaguchi K, Nagashima M, Suzuki T. Corynecin (chloramphenicol analogs) fermentation studies: selective production of Corynecin I by Corynebacterium hydrocarboclastus grown on acetate. Biotechnology and Bioengineering, (1977); 19(7): 1009-1018.
- Suzuki T, Honda H, Katsumata R. Production of Antibacterial Compounds Analogous to Chloramphenicol by a n-Paraffin-grown Bacterium. Microbiology & Fermentation Industry, (1972); 36(12): 2223-2228.
- Goh HF, Philip K. Purification and Characterization of Bacteriocin Produced by Weissella confusa A3 of Dairy Origin. PLoS One, (2015); 10(10): e0140434.
- Valenzuela AS, Benomar N, Abriouel H, Canamero MM, Galvez A. Isolation and identification of Enterococcus faecium from seafoods: antimicrobial resistance and production of bacteriocin-like substances. Food Microbiology, (2010); 27(7): 955-961.
- Ness IF, Diep DB, Ike Y. Enterococcal Bacteriocins and Antimicrobial Proteins that Contribute to Niche Control. 2014 Feb 16. In: Gilmore MS, Clewell DB, Ike Y, Shankar N, editors. Enterococci: From Commensals to Leading Causes of Drug Resistant Infection [Internet]. Boston: Massachusetts Eye and Ear Infirmary; 2014–. PMID: 24649514.
- Martin-Platero AM, Valdivia E, Ruiz-Rodriguez M, Soler JJ, Martin-Vivaldi M, et al. Characterization of antimicrobial substances produced by Enterococcus faecalis MRR 10-3, isolated from the uropygial gland of the hoopoe (Upupa epops). Applied and Environmental Microbiology, (2006); 72(6): 4245-4249.
- Balla E, Dicks LM, Du Toit M, Van Der Merwe MJ, Holzapfel WH. Characterization and cloning of the genes encoding enterocin 1071A and enterocin 1071B, two antimicrobial peptides produced by Enterococcus faecalis BFE 1071. Applied and Environmental Microbiology, (2000); 66(4): 1298-1304.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0