

![]()
Analysis of growth response and tolerance index of Glycine max
(L.) Merr. under hexavalent chromium stress
Hira Amin1, Basir Ahmed Arain1, Farah Amin2, Muhammad Ali Surhio2
Adv. life sci., vol. 1, no. 4, pp. 231-241, August 2014
*- Corresponding Author: Hira Amin (Email: hira.amin00@gmail.com)
Author Affiliations
2- National Centre of Excellence in Analytical Chemistry, University of Sindh, Jamshoro -76080 – Pakistan
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Metal pollution has become one of the most serious environmental problems due to various human activities. It can damage or alter the biosphere reducing the agricultural productivity and can affect both animals and humans. Emission of various pollutants into the atmosphere has many harmful effects on plant growth. Rapid urbanization, unregulated industrialization, growing transport, metal plating and agricultural activities have created a problem of heavy metals contamination.
Methods: A greenhouse experiment was conducted to determine the toxicity of chromium on Glycine max. Chromium concentration applied to G. max was managed as 0.5, 2.5, 5, 10, 25, 50 and 100 mg kg-1 for experimental period of 90 days. The phytotoxic effect of chromium metal was analyzed by studying seed germination, seedling vigor index, root and shoot length, root and shoot fresh and dry weights, chlorophyll content tolerance index.
Results: The data presented in this study showed that chromium metal adversely affects the seedling vigor of G. max and significantly (p<0.05) reduces seed germination and growth. The toxic effect of chromium on the seeds increased with increasing the concentration of the metal. It was also found that high concentrations of chromium (50 and 100 mg kg-1) can completely inhibit the seed germination.
Conclusion: The chromium metal is extremely toxic for seeds and young seedling of G. max at high concentrations. Moreover, G. max has little potential to counteract the deleterious effect of chromium metal in soil at aforementioned treatments. The results of the present study may help in better understanding of the mechanisms involved in pytoextraction.
Key word: Glycine max (L.) Merr., Chromium, Germination, Tolerance index
Introduction
Chromium (Cr) is an environmental pollutant that ranks seventh in abundance within the earth crust [1]. Naturally occurring Cr in soil ranges from 10 to 50 mg kg-1 [2]. The chemistry of Cr is very complex, its solubility, mobility and bioavailability in soil strongly depend on the various oxidation states from 0 to +6. Depending on its oxidation state and concentration, Cr acts as a toxic or essential element for living organism. The two most common species of Cr are Cr (III) and Cr (VI) available in anionic form as chromate, dichromate and hydro chromate ions [3].
The sources of Cr in environment are both natural and anthropogenic. Cr is used on a large scale in many industries, including electroplating, metallurgy, pigments, tanning, wood preservation, production of paints and Cr chemicals production, pulp and paper production [4]. The leather industry is the major cause of high efflux of Cr to the biosphere, accounting for 40% of the total industrial use [5]. Sewage and fertilizers are also the main sources of Cr [6]. These anthropogenic activities may lead to the widespread contamination in the environment [7].
A high concentration of Cr was found to be harmful for plant life. It reduces the protein contents, inhibit the enzyme activity, and causes chlorosis and necrosis [8]. The Cr concentration in plant adversely affects several morphological and biochemical parameters [9]. Cr toxicity interfere with several metabolic processes in plant, causing reduced seed germination or early seedling growth, biomass, photosynthetic impairing [10,11,12]. Phytotoxicity of Cr is considered inhibitory for plant growth. Its presence in surplus amount inside the plant can cause stunted growth [13]. The presence of Cr in soil can disturb the pattern of nutrient uptake in plant because of nutrient metal interaction [14]. Phytotoxic effects of Cr metal on seed germination and seedling growth of various vegetable crops such as Hibiscus esculentus (L.) [15], Daucus carrota (L.), Raphanus sativus (L.), Beta vulgaris (L.), Lycopersium esculentum (L.), Solanum melongena (L.), Vigna radiata (L.), Vigna angularis (L.), Lablab purpureus (L.), Lathyrus ordoratus (L.), Triticum aestivum (L.) has been reported [16,17].
The present study was conducted to investigate the phytotoxic effect of Cr and growth response of Glycine max (L.) to illustrate the potential of this species toward Cr metal stress.
Methods
Test plant material
Certified seeds of Glycine max (L.) were procured from National Agricultural Research Centre (NARC), Islamabad. Seeds of uniform size, color and weight were selected for experimental work and sterilized with 0.1% mercuric chloride solution and then rinsed with deionized water [18].
Soil Collection and Pot Preparation
For pot experiment soil samples were collected from riverbed of Indus, Jamshoro. The soil (2 part sand and 1 part clay) was air dried for 2-3 days. The pots were then filled with 5 kg air dried soil. The physicochemical properties of soil are given in table 1.
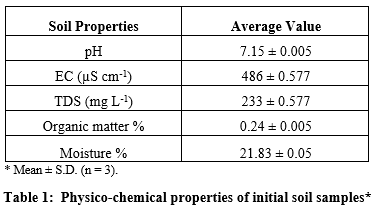
Chemicals and stock solution preparation
All chemicals used were of analytical grade; anhydrous potassium dichromate were procured from Fluka (Sigma-Aldrich, Spain) and used without any further purification. Stock solution of Cr (1000 mg L-1) was prepared by dissolving 2.28 g of potassium dichromate in 1000 ml of deionized water. The stock solution was then appropriately diluted to get the test solution of desired Cr concentration.
Experimental setup
A pot culture experiment was conducted at Green House of Institute of Plant Sciences, University of Sindh, Jamshoro. The experiment was arranged in complete randomized block design with three replicates for each treatment. To initiate the experiment under controlled condition, air dried soil artificially polluted with different concentration of potassium dichromate solution i.e. 0.5, 2.5, 5, 10, 25, 50 and 100 mg kg-1 respectively along with an untreated control. Cr solution was uniformly mixed with air dried soil and kept for 2 weeks to stabilize. Fifteen seeds were sown with uniform distance in each pot.
Germination and growth measurements
The seed germination was observed for every 24 hour until the germination percentage was constant. For the evaluation of seedling growth 10 germinated seedlings of similar morphology were allowed to grow with concentration of 0.5, 2.5, 5, 10, 25, 50 and 100 mg kg-1 Cr in soil, irrigated on alternate day with distilled water. The seedlings were harvested carefully after 90 days, washed with distilled water to remove soil particles and analyze for various growth responses. Growth response of G. max (L.) were studied in terms of root and shoot length (cm), root and shoot fresh weight (g), root and shoot dry weight (g). Seedlings were collected and cut at root-shoot junction and the length of their root and shoot were measured with a metric scale and expressed in centimeters [19]. The fresh weight of root and shoot samples were recorded on an analytical balance and expressed in gram per plant [20]. Later, plant parts were dried in an oven at 60 o C for 24 hour to get constant dry weight for root and shoot.
Germination percentage
The germination percentage is the proportion, expressed as percentage of germinated seeds to the total number of viable seeds that were tested by following formula: [21]
%G = (Number of germinated seeds / Total number of planted seeds) × 100 (1)
Seedling vigor index
Seedling vigor index are those properties of seed which determine the level of activity and performance of the seed during germination and seedling emergence. It is a single measurable property like germination describing several characteristics associated with various aspects of the performance of seed. Seedling vigor index is calculated by following formula: [22]
SVI = Germination percentage × Seedling length (2)
Stress tolerance index
Stress tolerance index is a useful tool for determining the high yield and stress tolerance potential of genotypes. Stress tolerance indices for different growth parameters were calculated using following formulae: [23]
RLSTI = (Root length of stress plant / Root length of control plant) × 100 (3)
SLSTI = (Shoot length of stress plant / Shoot length of control plant) × 100 (4)
RFSTI = (Root fresh weight of stress plant / Root fresh weight of control plant) × 100 (5)
SFSTI = (Shoot fresh weight of stress plant / Shoot fresh weight of control plant) × 100 (6)
RDSTI = (Root dry weight of stress plant / Root dry weight of control plant) × 100 (7)
SDSTI = (Shoot dry weight of stress plant / Shoot dry weight of control plant) × 100 (8) (8)
Chlorophyll measurement
Chlorophyll contents were determined spectrophotometrically. Leaves were cut into small pieces, mixed thoroughly and 0.25 g of leaves was taken into a mortar to grind them finely by pestle with 25 ml of 80% acetone for 5 minutes. The homogenate was filtered through filter paper (Whatman® No.42) and volume was made up to 25 ml with 80% acetone.
Extract monitoring by spectrophotometer
After the extraction, chlorophyll contents were monitored by UV-Visible spectrophotometer (Biochrom Libra S22). The absorbance of each solution was measured at 663 and 645 nm against 80% acetone blank in 1 cm quartz cuvette at room temperature. The amount of chlorophyll-a, chlorophyll-b and total chlorophyll (a+b) contents were calculated by using Arnon’s equation: [24]
Chl a (mg g-1) = [(12.7 × A663) − (2.69 × A645)] × ml acetone / mg leaf tissue (9)
Chl b (mg g-1) = [(22.9 × A645) − (4.68 × A663)] × ml acetone / mg leaf tissue (10)
Total Chl = Chl a + Chl b (11)
Statistical analysis
Data were statistically analyzed using one-way ANOVA on PASW® Statistics 18 (SPSS Inc., Chicago, IL, USA). The results are presented as means ± S.E. (standard errors) and data from the different treatments and control were compared by Duncan’s multiple-range test at p< 0.05.
Results
Seed germination under chromium stress
The result of present study reveals that higher concentration of Cr adversely influences the germination process of G. max seeds (Figure 1). The maximum germination percentage was noted in control by 86.67 ± 0.000. Cr treatments has significant (p<0.05) effect on seed germination by 77.78 ± 1.528, 71.11 ± 0.577, 68.89 ± 1.155, 60.00 ± 1.000, 53.33 ± 1.000 reduction in germination (Table 2). No data for seed germination was found in 50 and 100 mg Cr kg-1 soil treatment, indicating that higher level of Cr significantly (p<0.05)affected germination strength of G. max.
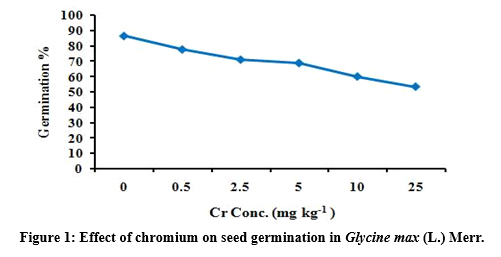

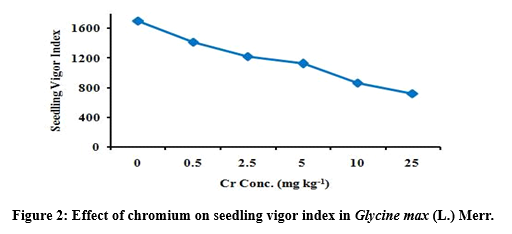
Seedling vigor index under chromium stress
Seedling vigor index of G. max was gradually decreased with increased Cr concentration at 0.5, 2.5, 5, 10 and 25 mg kg-1 (Table 2). The average seedling vigor index of G. max reduces from 1698.66, 1412.96, 1220.74, 1125.18, 866 and 720 respectively due to increase Cr level at aforesaid treatments as compared to control (Figure 2).
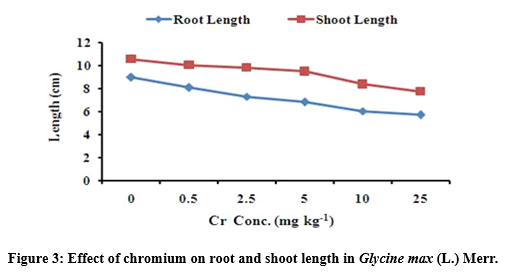
Root and shoot elongation under chromium stress
The results after 90th day exposure of Cr to G. max, showed considerable reduction in root and shoot elongation (Figure 3). The average values of root and shoot length were significantly (p<0.05) affected at 25 mg Cr kg-1. The maximum value of root length was 8.100 ± 1.153 followed by 7.333 ± 0.288 at 0.5 and 2.5 mg kg-1 and the minimum value of root length was 5.733 ± 0.680 at 25 mg kg-1 Cr treatment respectively. Similarly, the maximum value of shoot length was 10.067 ± 0.503 followed by 9.833 ± 1.154 at 0.5 and 2.5 mg kg-1 and the minimum values of shoot elongation was 7.767 ± 0.642 at 25 mg kg-1 treatment as compared to control plants respectively (Table 3).

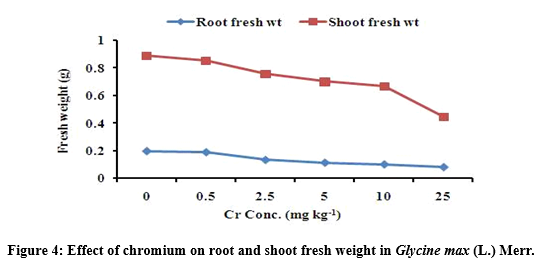
Root and shoot fresh weight under chromium stress
The root and shoot fresh weight of G. max was adversely affected due to increased Cr concentration in soil (Figure 4). Result shows that the root and shoot fresh weight was significantly (p<0.05) affected at 25 mg Cr kg-1. The maximum value of root fresh weight was 0.188 ± 0.033 followed by 0.133 ± 0.025 at 0.5 and 2.5 mg kg-1 and the minimum value of root fresh weight was 0.082 ± 0.010 at 25 mg Cr kg-1 treatments respectively. Similarly, the maximum value of shoot fresh weight was 0.854 ± 0.229 followed by 0.760 ± 0.329 at 0.5 and 2.5 mg kg-1 and the minimum values of average shoot fresh weight was 0.446 ± 0.051 at 25 mg Cr kg-1 respectively (Table 3).
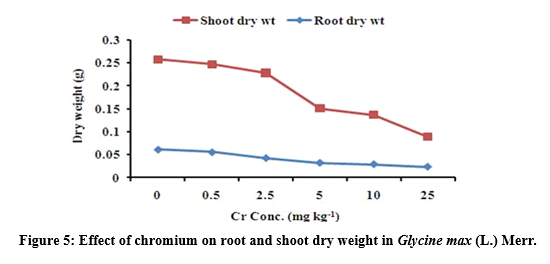
Root and shoot dry weight under chromium stress
The root and shoot dry weight of G. max was decreased with increased Cr level (Figure 5). Result shows that root dry weight was significantly (p<0.05) affected at 25 mg Cr kg-1. The maximum value of root dry weight was 0.055 ± 0.012 followed by 0.042 ± 0.014 at 0.5 and 2.5 mg kg-1 and the minimum value of root dry weight was 0.023 ± 0.004 at 25 mg kg-1 treatment respectively. Similarly, the maximum value of shoot dry weight was 0.191 ± 0.079 followed by 0.185 ± 0.084 at 0.5 and 2.5 mg kg-1 and the minimum value of shoot dry weight was 0.065 ± 0.006 at 25 mg kg-1 treatment respectively (Table 3).
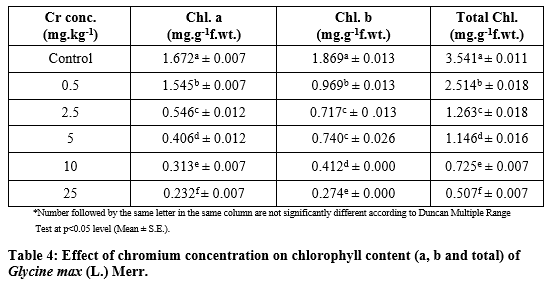
Chlorophyll contents under chromium stress
Table 4 shows the photosynthetic pigments chlorophyll-a, chlorophyll-b, total chlorophyll of G. max were significantly (p<0.05) decreased with increased Cr concentrations at 0.5, 2.5, 5, 10 and 25 mg kg-1 respectively. The chlorophyll-a in G. max leaves was decreased from 1.672 ± 0.007, 1.545 ± 0.007, 0.546 ± 0.012, 0.406 ± 0.012, 0.313 ± 0.007, and 0.232 ± 0.007 respectively. Similarly, the chlorophyll-b was decreased from 1.869 ± 0.013, 0.969 ± 0.013, 0.717 ± 0 .013, 0.740 ± 0.026, 0.412 ± 0.000 and 0.274 ± 0.000 respectively. The total chlorophyll contents were significantly (p<0.05) affected at high Cr concentration by 3.541 ± 0.011, 2.514 ± 0.018, 1.263 ± 0.018, 1.146 ± 0.016, 0.725 ± 0.007 and 0.507 ± 0.007 respectively.
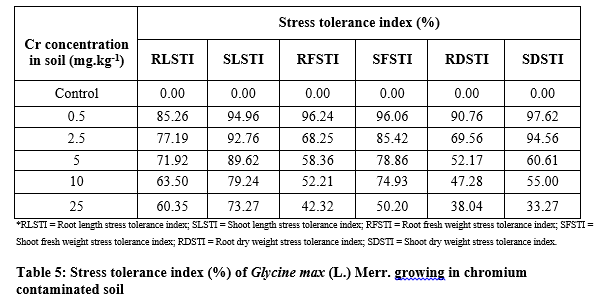
Tolerance index under chromium stress
Table 5 shows the higher Cr treatment lowers the percentage of tolerance in G. max. The highest value (85.26%) of RLSTI was recorded at 0.5 mg kg-1 and the lowest (60.35%) at 25 mg Cr kg-1 respectively. Similarly, the maximum SLSTI value (94.96%) was recorded at 0.5 mg kg-1 and the lowest (73.27%) at 25 mg Cr kg-1 respectively. The RFSTI of G. max was decreased as Cr level increase in the soil. The highest value (96.24%) for RFSTI was recorded at 0.5 mg Cr kg-1 and the lowest value (42.32%) at 25 mg Cr kg-1 respectively. Similarly, the maximum value for SFSTI (96.06%) was recorded at 0.5 mg Cr kg-1 and the lowest value (50.93%) at 25 mg Cr kg-1 respectively. The maximum value for RDSTI (90.76 %) was noted at 0.5 mg Cr kg-1 and the lowest value (38.04%) at 25 mg Cr kg-1 respectively. Similarly, the maximum SDSTI value (97.62%) was recorded at 0.5 mg kg-1 and the lowest value (33.27%) at 25 mg Cr kg-1 respectively.
Discussion
Cr is considered strongly toxic because Cr compounds in the soil are more or less insoluble as the metal ions are tightly bound to humus and clay particles [25]. It was reported that higher concentrations of heavy metal significantly reduced the strength of germination as compared to the lowest concentrations of heavy metal which has least harmful effect on germination [26]. The inhibition of seed germination by Cr was also observed by di Toppi et al. [27]. Similar results of Cr on seedling vigor index were reported by Ganesh Shanker et al. [28]. The root and shoot growth was much more sensitive to Cr toxicity than germination by this study. These results were also in agreement with that of Munzuroglu and Geckil [29]. It has been reported by several authors that the inhibition of root length caused by heavy metals is due to metal interference with cell division and cell elongation [25, 30]. The toxic effect of Cr on the root and shoot fresh weight in eight-day old seedling of Brassica oleracea L. var. acephala DC (kale) were reported by Ozdener et al. [31]. It was treated with various concentrations of chromium in the growth medium. Some other researchers have also observed that there is a gradual decrease of root and shoot fresh weight on treatment with increasing concentrations of Cr [32]. In a study conducted on Vallisneria spiralis to evaluate the Cr accumulation and toxicity in relation to biomass production, Vajpayee et al. found that dry matter production was severely affected by Cr concentration in nutrient medium [33].The elevated level of Cr also showed decreased level of Chlorophyll a and Chlorophyll b supported by Ganesh Shankar et al. [28]. Van Assche and Clijsters reported that heavy metals interfere with chlorophyll synthesis either through direct inhibition of an enzymatic step or by inducing deficiency of an essential nutrient [34]. Some other studies have the same conclusion that the content of chlorophyll, chlorophyll a/b ratio was decreased if kept under Cr 6+ stress (10-40 mg L-1) [35].
The root and shoot growth is an important parameter for classification of heavy metal tolerance [36]. The negative effect of Cr on tolerance indices of fresh and dry weight was also determined in rice [37]. A study on stress tolerance suggests that mechanism of tolerance helps plant to maintain growth even in the presence of potentially toxic metal concentrations [38].
This study concluded that the toxic effect of Cr metal on G. max growth depends upon the amount of toxic metal taken up from the specified environment. Seed germination time, seedling length (root and shoot length), fresh and dry weight (root & shoot weight), chlorophyll contents are apposite parameters to assess Cr toxicity on growth pattern of G. max test plant species. Plant tolerance index limitation showed G. max viability against Cr metal. Cr toxicity significantly inhibits seed germination and seedling growth of G. max at the higher concentration. Results of the study are useful indicator of Cr tolerance to some extent for plantation of G. max in Cr contaminated areas. However, G. max has shown a little potential to counteract the deleterious effect of Cr metal in soil at higher levels.
References
- Chandra P, Sinha S, Rai U, Kruger E, Anderson T, et al. Bioremediation of chromium from water and soil by vascular aquatic plants; 1997. American Chemical Society. pp. 274-282.
- Pandey SK, Pandey SK, Town Z. Germination and seedling growth of field pea Pisum sativum Malviya Matar-15 (HUDP-15) and Pusa Prabhat (DDR-23) under varying level of copper and chromium. J Am Sci, (2008); 4(3): 28-40.
- Gardea-Torresdey J, Peralta-Videa J, Montes M, De la Rosa G, Corral-Diaz B. Bioaccumulation of cadmium, chromium and copper by< i> Convolvulus arvensis</i> L.: impact on plant growth and uptake of nutritional elements. Bioresource Technology, (2004); 92(3): 229-235.
- Salunkhe P, Dhakephalkar P, Paknikar K. Bioremediation of hexavalent chromium in soil microcosms. Biotechnology letters, (1998); 20(8): 749-751.
- Barnhart J. Occurrences, uses, and properties of chromium. Regulatory toxicology and pharmacology, (1997); 26(1): S3-S7.
- Pillay A, Williams J, El Mardi M, Al-Lawati S, Al-Hadabbi M, et al. Risk assessment of chromium and arsenic in date palm leaves used as livestock feed. Environment international, (2003); 29(5): 541-545.
- Kotaś J, Stasicka Z. Chromium occurrence in the environment and methods of its speciation. Environmental pollution, (2000); 107(3): 263-283.
- Zayed A, Gowthaman S, Terry N. Phytoaccumulation of trace elements by wetland plants: I. Duckweed. Journal of environmental quality, (1998); 27(3): 715-721.
- Dube B, Tewari K, Chatterjee J, Chatterjee C. Excess chromium alters uptake and translocation of certain nutrients in citrullus. Chemosphere, (2003); 53(9): 1147-1153.
- Nath K, Saini S, Sharma YK. Chromium in tannery industry effluent and its effect on plant metabolism and growth. Journal of environmental biology/Academy of Environmental Biology, India, (2005); 26(2): 197-204.
- Kumar Sharma R, Agrawal M, Marshall F. Heavy metal contamination of soil and vegetables in suburban areas of Varanasi, India. Ecotoxicology and environmental safety, (2007); 66(2): 258-266.
- Zayed AM, Terry N. Chromium in the environment: factors affecting biological remediation. Plant and soil, (2003); 249(1): 139-156.
- Faisal M, Hasnain S. Chromate resistantBacillus cereus augments sunflower growth by reducing toxicity of Cr (VI). Journal of Plant Biology, (2005); 48(2): 187-194.
- Zupančič M, Bukovec N, Milačič R, Ščančar J. Comparison of various phosphate stabilisation agents for the immobilisation of Ni and Zn in sewage sludge. Water, air, and soil pollution, (2004); 156(1): 57-69.
- Amin H, Arain BA, Amin F, Surhio MA. Phytotoxicity of Chromium on Germination, Growth and Biochemical Attributes of Hibiscus esculentus L. American Journal of Plant Sciences, (2013); 2013.
- Shanker AK, Cervantes C, Loza-Tavera H, Avudainayagam S. Chromium toxicity in plants. Environment international, (2005); 31(5): 739-753.
- Jun R, Ling T, Guanghua Z. Effects of chromium on seed germination, root elongation and coleoptile growth in six pulses. International Journal of Environmental Science & Technology, (2009); 6(4): 571-578.
- Ramasubramanian V, Ravichandran V, Kannan N. Analysis of industrial effluents and their impact on the growth and metabolism of Phaseolus mungo, L. Communications in Soil Science & Plant Analysis, (1993); 24(17-18): 2241-2249.
- Chou C-H, Lin H-J. Autointoxication mechanism ofOryza sativa I. Phytotoxic effects of decomposing rice residues in soil. Journal of Chemical Ecology, (1976); 2(3): 353-367.
- Bush DS. Calcium regulation in plant cells and its role in signaling. Annual review of plant biology, (1995); 46(1): 95-122.
- Akinci IE, Akinci S. Effect of chromium toxicity on germination and early seedling growth in melon (Cucumis melo L.). African Journal of Biotechnology, (2010); 9(29): 4589-4594.
- Abdul-Baki AA, Anderson JD. Vigor determination in soybean seed by multiple criteria. Crop science, (1973); 13(6): 630-633.
- Wilkins D. A technique for the measurement of lead tolerance in plants. (1957).
- Arnon DI. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant physiology, (1949); 24(1): 1.
- Kidd P, Díez J, Martínez CM. Tolerance and bioaccumulation of heavy metals in five populations of Cistus ladanifer L. subsp. ladanifer. Plant and soil, (2004); 258(1): 189-205.
- Peralta J, Gardea-Torresdey J, Tiemann K, Gomez E, Arteaga S, et al. Uptake and effects of five heavy metals on seed germination and plant growth in alfalfa (Medicago sativa L.). Bulletin of Environmental Contamination and toxicology, (2001); 66(6): 727-734.
- di Toppi LS, Fossati F, Musetti R, Mikerezi I, Favali M. Effects of hexavalent chromium on maize, tomato, and cauliflower plants. (2002).
- Ganesh KS, Sundaramoorthy P, Chidambaram A. Chromium toxicity effect on blackgram, soybean and paddy. POLLUTION RESEARCH, (2006); 25(4): 757.
- Munzuroglu O, Geckil H. Effects of metals on seed germination, root elongation, and coleoptile and hypocotyl growth in Triticum aestivum and Cucumis sativus. Archives of Environmental Contamination and Toxicology, (2002); 43(2): 203-213.
- Rout G, Samantaray S, Das P. Differential chromium tolerance among eight mungbean cultivars grown in nutrient culture. Journal of plant Nutrition, (1997); 20(4-5): 473-483.
- Ozdener Y, Aydin BK, Fatma Aygün S, Yürekli F. Effect of hexavalent chromium on the growth and physiological and biochemical parameters on Brassica oleracea L. var. acephala DC. Acta Biologica Hungarica, (2011); 62(4): 463-476.
- Anjum MF, Zia MA, Ashraf M, Khalid Z (2011) Effect of Chromium on Growth Attributes in Sunflower (Helianthus annuus L.). Survival and Sustainability: Springer. pp. 985-994.
- Vajpayee P, Rai U, Ali M, Tripathi R, Yadav V, et al. Chromium-induced physiologic changes in Vallisneria spiralis L. and its role in phytoremediation of tannery effluent. Bulletin of Environmental Contamination and toxicology, (2001); 67(2): 246-256.
- Van Assche F, Clijsters H (1983) Multiple effects of heavy metal toxicity on photosynthesis. Effects of stress on photosynthesis: Springer. pp. 371-382.
- Rai V, Vajpayee P, Singh SN, Mehrotra S. Effect of chromium accumulation on photosynthetic pigments, oxidative stress defense system, nitrate reduction, proline level and eugenol content of Ocimum tenuiflorum L. Plant science, (2004); 167(5): 1159-1169.
- Diwan H, Khan I, Ahmad A, Iqbal M. Induction of phytochelatins and antioxidant defence system in Brassica juncea and Vigna radiata in response to chromium treatments. Plant Growth Regulation, (2010); 61(1): 97-107.
- Panda SK. Chromium-mediated oxidative stress and ultrastructural changes in root cells of developing rice seedlings. Journal of plant physiology, (2007); 164(11): 1419-1428.
- Clemens S. Toxic metal accumulation, responses to exposure and mechanisms of tolerance in plants. Biochimie, (2006); 88(11): 1707-1719.