

![]()
Defense strategies of cotton against whitefly transmitted CLCuV and Begomoviruses
Muhammad Azmat Ullah Khan*1, Ahmad Ali Shahid1, Abdul Qayyum Rao1, Naila Shahid1, Ayesha Latif1, Salah ud Din1, Tayyab Husnain1
Adv. life sci., vol. 2, no. 2, pp. 58-66, February 2015
*–Corresponding Author: Muhammad Azmat Ullah Khan (Email: mazmatullahkhan@yahoo.com)
Author Affiliations[Date Received: 13/12/2014; Date Revised: 12/02/2015; Date Published Online: 25/02/2015]
Abstract![]()
Introduction
Methods
Discussion
Conclusion
References
Abstract
Cotton leaf curl virus (CLCuV) is economically important monopartite Geminivirus which is transmitted by whitefly in persistent circulative manner. In Pakistan, CLCuV causes severe damage to Gossypium hirsutum whereas G. arboreum is resistant to this virus. A total of 86 articles were included in this study that were searched through web of knowledge, web of science and google scholar by using the keywords of cotton, CLCuV, waxes, Begomoviruses and transgenic techniques used against Begomoviruses in plants. Various transgenic strategies i.e., pathogen derived resistance (with and without protein expression) and non – pathogen derived resistance have been adopted to control this virus or its vector. Beside these, some natural defense mechanisms of plant also protect it against the vector. The cuticular waxes make the insects’ attachment difficult to plant surfaces and act as a physical barrier. The cuticular waxes in G. arboreum act as first line of defense against whiteflies and thus CLCuV. Some other defense strategies may involve hindering the insect movements or depriving it from food due to thick waxy layer. Biotechnological strategies against various Begomoviruses including CLCuV were found successful in some crops except cotton. Whereas, the natural defense strategies in G. arboreum i.e., long trichome or presence of inorganic salts with increased concentration of waxes, provide good defense strategy against whiteflies, CLCuV and other pathogens.
Key words: Cotton, CLCuV, Begomoviruses, Waxes, Defense Strategy
Introduction
Cotton
The English word cotton came from Arabic word “al qatan” or “kutun” [1]. At first cotton means merely a fine textile and the word broadly included the flax. History of cotton plant cultivation in the world is very old. Pakistan is an ancient home of cotton cultivation. The oldest and one of the first discoveries of cotton usage in the world excavated at Mohenjo-Daro in Sindh, bear testimony to the proficiency of Indus civilization in the use of cotton as far backs as 3000 B.C. [2].
Cotton is the backbone of textile industry, it is a cheap source of fiber as well as used as an oilseed crop. It has also gained importance in bioenergy production. Top four cotton growing countries, India, China, USA and Pakistan are contributing for approximately two third of the world’s cotton. The statistics presented by the OECD-FAO (Food and Agriculture Organization) of the United Nations in 2013, depicts 1.7% increase in cotton production across the world. Cotton share 4.6% area among all seeds and grain crops and world total cotton production was around 25 Mt (million tons) in 2012 [3].
Among all the species of Gossypium, the upland cotton, Gossypium hirsutum also known as Maxican or long staple cotton, rank first in total cotton share which is about 90%. Gossypium barbadense is ranked 2nd having the 8% of the world’s cotton shares while Gossypium herbaceum (Levant Cotton) and Gossypium arboreum (Tree Cotton) both contribute 2% of the world’s cotton [3].
G. arboreum (Tree Cotton), although it gives low yield but has many important agronomical characteristics such as disease and insect resistance, high fiber strength and tremendous plasticity, which are absent in G. hirsutum (Upland Cotton). Beside these G. arboreum can be grown under poorest growing conditions and low managed area.
Problems associated with cotton crop
Biotic and abiotic factors equally contribute toward reduction of cotton yield. Various pathogens including fungi, bacteria and viruses attack cotton plant and cause many diseases. Among all pathogens thriving on cotton plant whitefly is responsible for 50% yield loss. It reduces boll formation and most importantly serves as a vector for Cotton Leaf Curl Virus (CLCuV). CLCuV is a single stranded DNA (ssDNA) virus that belongs to Geminiviruses and serves as a major threat to subcontinent cotton [4]. The Cotton Leaf curl Disease (CLCuD) was first recorded in 1967 in Pakistan on G. hirsutum. The first severe epidemic in Pakistan appeared in 1992-93 which caused 7.9 million reductions in cotton bales. It was estimated that during 1992-97 CLCuV caused $5 billion losses to cotton sector only in Pakistan [5].G. arboreum (Desi cotton) possess high resistance to root rot disease [6] and to the Geminiviridae viruses [7]. It also contains desirable genes for stress tolerance not only against abiotic but also against biotic factors i.e., resistant against insects (bollworms, aphids and whiteflies) as well as fungal diseases (root and stem rot) [8,9].
Viruses
Viruses are able to infect all type of organisms including plants, animals, bacteria, and Archaea [10]. Viruses are ubiquitously present in all ecosystem around world [11].Viruses influence all cellular life. Viruses are very specific in their host infectivity and cause infection only to their specific host [12]. Some viruses can only replicate within cells that have already been infected by another virus. These are known as satellites viruses [13]. The virus that complete their life cycle in plants are called as plant viruses. As all the viruses are obligate intracellular parasites, plant viruses are also dependent upon plant cell machinery to complete their replication.
Geminiviruses
Geminiviruses are group of small insect transmitted plant-pathogenic viruses responsible for various crop diseases in tropical and subtropical regions of the world [14]. Significant crop losses are caused by these viruses sometimes leading to the epidemics. Various factors resulting into crop epidemics involve the recombination of different Geminiviruses coinfecting the same plant, the expansion of agriculture into new growing areas and transport of infected plant material to new locations. Furthermore, the expansion and migration of vectors that can spread the virus from one plant to another can also cause large epidemics [15]. Geminiviruses have become the subjects of intensive research because of their implications to plant pathology, plant molecular biology, and plant biotechnology.
Begomoviruses
The most important genus of Geminiviruses is Begomoviruses. Begomovirus constitute the largest and economically most important genus containing over 200 species so far and their number is still rising [16]. Begomoviruses can be divided into two groups based on their genome organization; most have their genome split between two DNA molecules of approximately 2600 n.t. each, termed DNA-A and DNA-B (bipartite), while others have a single DNA-A-like genome of about 2800 n.t. (monopartite) [17]. In the last decade, production of important crops such as cassava, cotton, cucurbits, legumes, peppers and tomato had been affected to a greater extent by Begomoviruses [4]. Virus species in this genus are transmitted in a persistent circulative manner by the whitefly Bemisia tabaci (Gennadius). They infect dicotyledonous plants and cause several of the most devastating and important plant diseases including cassava mosaic, cotton and tomato leaf curl and bean golden mosaic with losses estimated up to several billions of dollars [18].
Whitefly and Begomoviruses
Over the last three decades, insects belonging to the genus Bemisia (Homoptera: Aleyrodidae), commonly known as whiteflies, became one of the major pests in the worldwide agricultural systems [19]. The most widespread and important are those whiteflies that are associated with viruses classified in the family Geminiviridae, more specifically to those belonging to the genus Begomovirus, The Begomoviruses are transmitted by the whitefly Bemisia tabaci (Gennadius) in a persistent circulative manner. Their spread may be directly linked to the inadvertent world-wide dissemination of the "B" or silverleaf biotype of the whitefly Bemisia tabaci. This vector is an indiscriminate feeder, encouraging rapid and efficient spread of viruses from indigenous plant species to neighboring crops [20]. A major impact of the increase in the population of whiteflies was the proliferation of disease problems caused by viruses that whiteflies transmit.
Methods
The following study was conducted by searching the key terms; CLCuV, Cotton, Gossypium, waxes of cotton, Insect-wax relation, Begomoviruses, plant defense strategy against Begomoviruses in the search engines of web of sciences, web of knowledge and google scholar. A total of 86 articles were included in this study from 143 papers which present the information about Begomoviruses, cotton and defense strategies against Begomoviruses. The defense strategy was further categorized into defense by biotechnological methods and natural defense strategy by cotton against microbes, viruses and pest.
Discussion
Different approaches used against “Begomoviruses”
The basic strategy used for control of Begomoviruses is the use of insecticide against whiteflies but this method is not reliable because it might be too late to control the infection in plants as whiteflies might have already transmitted the viruses. Furthermore, insecticides have adverse effect on environment and also cause acute toxicity to the living organisms [21]. Keeping the insects away from the plants is not the best option to control the disease instead, resistance against the viruses in plants can be incorporated by biotechnological methods which could be the best solution [22]. Resistance against viruses may be achieved by pathogen derived resistance or non-pathogen derived resistance.
Pathogen derived resistance
Pathogen derived resistance can be divided in two categories as:
- With protein expression
- Without protein expression.
Pathogen derived resistance with protein expression:
Replication associated protein generally called Rep proteins play vital role in replication of virus inside host cell. They are thought to be involved in regulation of host cell replication. Rep proteins bring the cell to “S” phase and trigger the synthesis of the viral component [16,23]. So, interfering the structure of Rep proteins may result in failure of the synthesis of viral components. This strategy was first described by the Hong and Stanley (1996) by truncating the N-terminal portion of Rep in African Cassava Mosaic Virus (ACMV) which ultimately inhibited the replication of the virus in host protoplasts. Similar strategies were developed against Tomato Yellow Leaf Curl Sardinia Virus (TYCSV) and Tomato Leaf Curl New Dehli Virus (ToLCNDV) [24]. No commercial variety of cotton contain siRNA technology which might question the successfulness of this technology. Currently intergenic region of Ageratum conyzoides has been transformed against the CLCuV in cotton but the transgenic plants are under trial (un-published).
Pathogen derived resistance without protein expression:
Gene Silencing: The main component of pathogen derived resistance without protein expression is RNA interference. In this phenomenon, the mRNA produced by viral gene is either degraded or demethylation of DNA takes place hence viral proteins are not formed due to induced production of interfering RNA. This strategy was successfully used to control African Cassava Mosaic Virus (ACMV) [25], Mungbean Yellow Mosaic Virus (MYMV) [26] Tomato Leaf Curl Virus (ToLCV) [27] Tomato Yellow Leaf Curl Virus (TYLCV) [26] Sri Lankan Cassava Mosiac Virus and East African Cassava Mosaic Virus (EACMV) [28].
Antisense RNA: An RNA molecule that is complementary to the particular mRNA is called “Antisense RNA”. This antisense RNA molecule makes duplex with mRNA and prevent it for being translated [29]. This technique has also been widely used since 1991 and very effective against several viruses like Tomato golden mosaic virus [30] TYLCV [31] Cotton Leaf Curl Kokhran Virus (CLCuKoV) [29] and ACMV [32].
Non pathogen derived resistance
The first report of transgenic resistance against a plant virus involved the expression of coat protein (CP) gene of Tobacco mosaic virus (TMV) [33] and this strategy was subsequently tried for Geminiviruses [34]. Tomato plants expressing the CP of TYLCV exhibited delayed symptoms development and later on showed recovery of symptoms which was dependent on the expression level of the CP [34]. Although the precise mechanism remains unclear, it may be due to inappropriate timing of expressionor overexpression of the CP which confiscate viral genomic ssDNA. It has also been suggested that CP prevents the association of virus movement proteins with the ssDNA and thus inhibits viral movement into the next cell [35].
Plant self defense mechanisms
Plants have some effective and amplified defense methods against viruses. One of the most considerable is the presence of resistance (R) genes. Each R gene provide resistance to a peculiar virus by triggering localized areas of cell death around the infected cell, which could be seen by unaided eye as large spots. So, the infection is blocked from being spread by this way [27]. RNA interference is also an effective defense mechanism in plants [36].When plants are infected, they usually produce natural disinfectants such as salicylic acid, nitric oxide and reactive oxygen molecules that kill the pathogens [37]. Plant defense against viral infection also constitute, the use of siRNA in response to dsRNA [38]. Most of the plant viruses encode a protein to suppress this response [39]. Plants may also reduce transport through plasmodesmata in response to injury [40].
Plant physiological adaptation against insects
Basic requirement in plant – insect interaction is attachment of insect with plant surface. If insects are unable to attach themselves, no plant – insect pathogen interaction would be possible. Most plants have developed some physiological properties that either increase the grip of the insect or inhibit them [41].
Hindering insect movement
The wild cotton G. arboreum has the ability to hinder the insect movement by its specialized structure called trichome. Trichomes offer difficulty in moving, piercing and sucking of insects [42]. Length of the trichome and its density is very important for insect behavior. Sometimes trichome also act as an indicator of insect attack as plant release immediate defense chemicals when trichomes are broken [43].
Depriving of food for insect
Terrestrial plants including cotton outer leaf surfaces also contain calcium oxalate and other inorganic salts. When insect tries to eat plants, these chemical compounds damage the mouthparts and gut [44].
Epicuticular waxes
A protective epicuticular wax layer is present on a plant surface that is highly variable depending upon species, cultivar or plant part. Chemically wax can be defined as a polyester matrix of hydroxyl- and hydroxyl epoxy fatty acids (C16-C18). Biochemical analysis of wax showed that it is the lipid fraction removed from plant surfaces after brief immersion in an organic (nonpolar) solvent, like chloroform or hexane [45]. The epicuticular wax is a complex mixture of organic compounds like hydrocarbons, alcohols, acids and esters [46] among them esters are most common organic compounds. Ester may be of different types i.e., alkyl esters, amyryl esters, cycloartenyl esters and triacylglycerols [47].
A layer of cuticle composed of cutin and wax, is present on plant surfaces particularly nonwoody plant surfaces which protect the plant from environmental stresses [48]. Intracuticular (impregnated in) and epicuticular (exterior to the cutin biopolymer) both types of wax are present on plant surface. In some species epicuticular wax is present in the form of smooth film, while in some other species it is in the form of protruding wax crystals [48]. Wax layer is not only present on plant epidermal cells, but it is also part of specialized cells of plant reproductive organs e.g. stigmatic papillae [49]. Long chain fatty acids and phenylpropanoids (sporopollenin) may also present on outer wall of pollen [50].
Complex mixtures of very long chain aliphatics and cyclic compounds constitute plant cuticular waxes. The aliphatic compounds consist of fatty acids, aldehydes, primary and secondary alcohols, ketones, and alkanes having chain lengths ranging from C16 to C36 in homologous series. Along with these aliphatic compounds, cyclic compounds such as triterpenoids, tocopherols, and aromatic compounds may be present in either large or small quantities, depending upon the species [51].
Role and significance of wax in plants
Unlike animals, plants are sessile they do not show any physical motion to escape from the extreme of biotic and abiotic stresses. So plants have developed specialized structures to tolerate these stresses and epicuticular wax is one of them [52]. It is present on plant surfaces that are directly exposed to the environmental or biotic stresses and there it acts as first line of defense to prevent stomatal water loss, which is the primary function of cuticle [53]. Other secondary functions of epicuticular waxes are protection from UV reflection [54], first physical barrier to the external organisms i.e. plant insect and role in plant-pathogen interactions [55].
Epicuticular waxes are chemically composed of such components that form a continuous hydrophobic layer, which restricts water loss from plant organs. It had been shown that plant surfaces having reduced amounts of wax layer have been associated with increased rates of transpiration. Brushing waxes off the excised leaves significantly increased the rate of water loss [56]. The cuticular waxes play vital role in plant defense against insect as follows:
Making insect attachment difficult
Plant surfaces might be slippery because of several ways. The most common is because of the covering of epicuticular waxes. The basic mechanism of this slimy surface is the falling of the crystals when insects try to attach with them [57]. On the other hand the epicuticular wax also creates small crystals that make the surface rough and decrease the surface area for insect attachment [58].
Plants epicuticular wax can be a major strategy to control abiotic as well as biotic diseases [52].The epicuticular wax also acts as a barrier against natural predators overcoming their success to penetrate the plants through cuticle and other openings [59]. For example, Aphidius ervi take more time for actively foraging on wax-deficient pea mutants [60]. The key factor of plant waxes that determine the foraging success of different predators is their ability to attach to the plant surface. Better inset attachment to the plant surface is thought to be associated with abridged wax load [59].
Epicuticular wax genes
Biochemical analysis of the mutant phenotypes has played an important role in the identification of plant epicuticular wax genes. Glossy appearance of mutant plants appeared to be linked with the deficiency in wax layer. Several mutants of different plant species deficient in wax layer have been studied [61].
The wax composition due to CER2 gene of Arabidopsis (Accession No. CP002684.1) resulted in obstruction of the elongation of fatty acids C26 [62]. When the CER2 gene was isolated [63], the resulted amino acid sequence had no homology to any previously known gene product, giving no idea of its biochemical function.
Epicuticular wax and cotton
Epicuticular wax ridges protect the upper and lower surfaces of cotton leaf by forming an amorphous layer of cuticle [64]. The thickness of epicuticular wax in cotton leaf is about 30 µm, and this thickness increases up to 30% with treatment of dehydration [1]. The wax deposition on cotton leaf, bract and boll increases upto 68.57%, 46.8% and 4.1% respectively under water stress and act as a barrier against non-stomatal water loss [65].
Main constituents of the leaf epicuticular wax under water stress were long chain alkanes like, n-octacosane, n-nanocosane, n-triacontane, dotricocontane, and n-tetracotane [65]. Similarly, in bract wax n-octacosane and n-tricotane, in boll n-nanocosane and n-tricosane were the major long chain alkanes. Wax morphology of cotton leaf, bract and boll under both water stress and well watered condition were same, as shown by scanning electron microscopy. Exposure of cotton leaf to UV-B radiation also increases epicuticular wax deposition [54].
Wax mutants
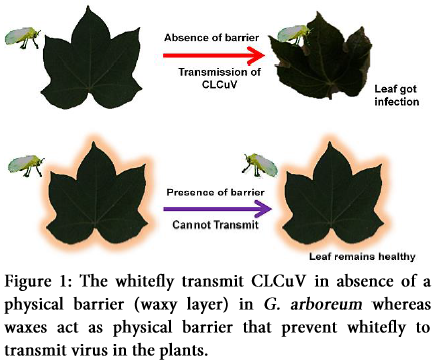
Analysis of wax mutant phenotypes has also been carried out after the epicuticular wax genes identification through molecular biology techniques. Most wax mutants share visual morphological similarities to the wild type, except for their glossy appearance [66]. Epicuticular wax mutants were first identified by Dellaert et al. [67] in Arabidosis referred them as eceriferum (cer), which in Latin is “without wax”. Wax deficient mutants have been isolated in a number of plant species, including barley (Hordeum vulgare), Arabidopsis, maize (Zea mays) and Brassica napus. Barley and Arabidopsis have eceriferum (cer) mutant loci, whereas in maize and B. napus were termed as glossy [68]. An attempt was made to evaluate epicuticular wax of leaves and whitefly relationship based on the quantity (determined by biochemical test) and quality of wax (determined by GC-MS) present in different plant species. Study revealed that when less waxy mutant of G. arboreum; GaWM3 were created and viruliferous whiteflies were allowed to feed on them, the plants become infected with the visible symptoms of leaf curling and thickened veins. Viruses were also screened through PCR among these plants with G. hirsutum as positive control and G. arboreum as negative control [52].
Conclusion
The pathogen derived resistance with protein expression was bit successful in tomato against Begomoviruses but not in cotton. Similarly, the pathogen derived resistance without protein expression i.e., gene silencing and anti-sens RNA was successful against Begemoviruses including CLCuV but in other crops not in cotton. But natural defenses in wild species of cotton (G. arboreum) i.e., long trichome or presence of inorganic salts with increased concentration of waxes, provide good defense strategy against whiteflies, CLCuV and other pathogens. Cuticular wax in G. arboreum not only protects plants from heat, drought and UV stresses but also make it difficult for whiteflies to feed and transmit viruses to the plants.
References
- Weete JD, Leek GL, Peterson CM, Currie HE, Branch WD. Lipid and surface wax synthesis in water-stressed cotton leaves. Plant Physiology, (1978); 62(5): 675-677.
- Rahman M, Hussain D, Zafar Y. Estimation of genetic divergence among elite cotton cultivars–genotypes by DNA fingerprinting technology. Crop Science, (2002); 42(6): 2137-2144.
- Jiang C-X, Wright RJ, El-Zik KM, Paterson AH. Polyploid formation created unique avenues for response to selection in Gossypium (cotton). Proceedings of the National Academy of Sciences, (1998); 95(8): 4419-4424.
- Mansoor S, Briddon RW, Zafar Y, Stanley J. Geminivirus disease complexes: an emerging threat. Trends in Plant Science, (2003); 8(3): 128-134.
- Sabahat H. Differential eefect of planting time on cotton leaf curl disease and yeild of cotton vriety CIM-598. Journal of Novel Research in Life Sciences, (2015); 2(1): 1-7.
- Wheeler T, Gannaway J, Keating K. Identification of resistance to Thielaviopsis basicola in diploid cotton. Plant Disease, (1999); 83(9): 831-833.
- Zafar Y, Mansoor S, Asad S, Briddon R, Idrees M, et al. Genome Characterization of whitefly-transmitted geminivirus of cotton and Development of Virus-resistant Plants through Genetic Engineering and conventional Breeding. ICAC Recorder USA, (2003); 12-16.
- Borole V, Dhumale D, Rajput J. Embryo culture studies in interspecific crosses between arboreum and hirsutum cotton. Indian Journal of Genetics & Plant Breeding, (2000); 60(1): 105-110.
- Huang G-T, Ma S-L, Bai L-P, Zhang L, Ma H, et al. Signal transduction during cold, salt, and drought stresses in plants. Molecular Biology Reports, (2012); 39(2): 969-987.
- Koonin EV, Senkevich TG, Dolja VV. The ancient Virus World and evolution of cells. Biol Direct, (2006); 1(1): 29.
- Edwards RA, Rohwer F. Viral metagenomics. Nature Reviews Microbiology, (2005); 3(6): 504-510.
- Dimmock NJ, Easton AJ, Leppard K, Dimmock N, Easton A, et al. Introduction to modern virology. Chapter: Book Name. 2007 of publication; Blackwell Pub. Malden, MA, USA.
- La Scola B, Desnues C, Pagnier I, Robert C, Barrassi L, et al. The virophage as a unique parasite of the giant mimivirus. Nature, (2008); 455(7209): 100-104.
- Moffat AS. Geminiviruses emerge as serious crop threat. Science, (1999); 286(5446): 1835-1835.
- Gray SM, Banerjee N. Mechanisms of arthropod transmission of plant and animal viruses. Microbiology and Molecular Biology Reviews, (1999); 63(1): 128-148.
- Gutierrez C, Ramirez-Parra E, Mar Castellano M, Sanz-Burgos AP, Luque A, et al. Geminivirus DNA replication and cell cycle interactions. Veterinary Microbiology, (2004); 98(2): 111-119.
- Fauquet CM, Stanley J. Geminivirus classification and nomenclature: progress and problems. Annals of Applied Biology, (2003); 142(2): 165-189.
- Varma A, Malathi V. Emerging geminivirus problems: a serious threat to crop production. Annals of Applied Biology, (2003); 142(2): 145-164.
- Ribeiro S, Ambrozevicius L, Avila Ad, Bezerra I, Calegario R, et al. Distribution and genetic diversity of tomato-infecting begomoviruses in Brazil. Archives of Virology, (2003); 148(2): 281-295.
- Brown JK, Czosnek H. Whitefly transmission of plant viruses. Advances in Botanical Research, (2002); 3665-100.
- Seal S, Jeger M, Bosch FVd. Begomovirus evolution and disease management. Advances in Virus Research, (2006); 67297-317.
- Lehmann Pa. Structure and evolution of plant disease resistance genes. Journal of Applied Genetics, (2002); 43(4): 403-414.
- Stanley J. Analysis of African cassava mosaic virus recombinants suggests strand nicking occurs withinthe conserved nonanucleotide motif during the initiation of rolling circle DNA replication. Virology, (1995); 206(1): 707-712.
- Brunetti A, Tavazza M, Noris E, Tavazza R, Caciagli P, et al. High expression of truncated viral Rep protein confers resistance to tomato yellow leaf curl virus in transgenic tomato plants. Molecular Plant-Microbe Interactions, (1997); 10(5): 571-579.
- Vanderschuren H, Alder A, Zhang P, Gruissem W. Dose-dependent RNAi-mediated geminivirus resistance in the tropical root crop cassava. Plant Molecular Biology, (2009); 70(3): 265-272.
- Shepherd DN, Martin DP, Thomson JA. Transgenic strategies for developing crops resistant to geminiviruses. Plant Science, (2009); 176(1): 1-11.
- Zrachya A, Kumar PP, Ramakrishnan U, Levy Y, Loyter A, et al. Production of siRNA targeted against TYLCV coat protein transcripts leads to silencing of its expression and resistance to the virus. Transgenic Research, (2007); 16(3): 385-398.
- Chellappan P, Masona MV, Vanitharani R, Taylor NJ, Fauquet CM. Broad spectrum resistance to ssDNA viruses associated with transgene-induced gene silencing in cassava. Plant Molecular Biology, (2004); 56(4): 601-611.
- Asad S, Haris W, Bashir A, Zafar Y, Malik K, et al. Transgenic tobacco expressing geminiviral RNAs are resistant to the serious viral pathogen causing cotton leaf curl disease. Archives of Virology, (2003); 148(12): 2341-2352.
- Day A, Bejarano E, Buck K, Burrell M, Lichtenstein C. Expression of an antisense viral gene in transgenic tobacco confers resistance to the DNA virus tomato golden mosaic virus. Proceedings of the National Academy of Sciences, (1991); 88(15): 6721-6725.
- Praveen S, Mishra AK, Dasgupta A. Antisense suppression of replicase gene expression recovers tomato plants from leaf curl virus infection. Plant Science, (2005); 168(4): 1011-1014.
- Zhang P, Vanderschuren H, Fütterer J, Gruissem W. Resistance to cassava mosaic disease in transgenic cassava expressing antisense RNAs targeting virus replication genes. Plant Biotechnology Journal, (2005); 3(4): 385-397.
- Abel PP, Nelson RS, De B, Hoffmann N, Rogers SG, et al. Delay of disease development in transgenic plants that express the tobacco mosaic virus coat protein gene. Science, (1986); 232(4751): 738-743.
- Kunik T, Salomon R, Zamir D, Navot N, Zeidan M, et al. Transgenic tomato plants expressing the tomato yellow leaf curl virus capsid protein are resistant to the virus. Nature Biotechnology, (1994); 12(5): 500-504.
- Briddon R, Markham P. Cotton leaf curl virus disease. Virus Research, (2000); 71(1): 151-159.
- Shors T. Chapter 20. Understanding viruses: (2011). 1: 614-641. Jones & Bartlett Publishers
- Soosaar JL, Burch-Smith TM, Dinesh-Kumar SP. Mechanisms of plant resistance to viruses. Nature Reviews Microbiology, (2005); 3(10): 789-798.
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, et al. Chapter 3. Molecular Biology of the Cell, (1997). 1:95-106. Garland Science, New York.
- Ding S-W, Voinnet O. Antiviral immunity directed by small RNAs. Cell, (2007); 130(3): 413-426.
- Oparka KJ, Roberts AG. Plasmodesmata. A not so open-and-shut case. Plant Physiology, (2001); 125(1): 123-126.
- Koch K, Barthlott W. Superhydrophobic and superhydrophilic plant surfaces: an inspiration for biomimetic materials. Philosophical Transactions of the Royal Society A: Mathematical, Physical and Engineering Sciences, (2009); 367(1893): 1487-1509.
- Whitney HM, Federle W. Biomechanics of plant–insect interactions. Current Opinion in Plant Biology, (2013); 16(1): 105-111.
- Alcorn K, Whitney H, Glover B. Flower movement increases pollinator preference for flowers with better grip. Functional Ecology, (2012); 26(4): 941-947.
- Lucas PW, Turner IM, Dominy NJ, Yamashita N. Mechanical defences to herbivory. Annals of Botany, (2000); 86(5): 913-920.
- Post-Beittenmiller D. The cloned Eceriferum genes of Arabidopsis and the corresponding Glossy genes in maize. Plant Physiology and Biochemistry, (1998); 36(1): 157-166.
- Chachalis D, Reddy KN, Elmore CD. Characterization of leaf surface, wax composition, and control of redvine and trumpetcreeper with glyphosate. Weed Science, (2001); 49(2): 156-163.
- Shepherd T, Robertson G, Griffiths D, Birch A. Epicuticular wax ester and triacylglycerol composition in relation to aphid infestation and resistance in red raspberry (Rubus idaeus L.). Phytochemistry, (1999); 52(7): 1255-1267.
- Riederer M, Muller Chapter 20. Annual Plant Reviews, Biology of the Plant Cuticle, (2009). 35:165-192. John Wiley & Sons.
- Lolle SJ, Pruitt RE. Epidermal cell interactions: a case for local talk. Trends in Plant Science, (1999); 4(1): 14-20.
- Guilford WJ, Schneider DM, Labovitz J, Opella SJ. High resolution solid state 13C NMR spectroscopy of sporopollenins from different plant taxa. Plant Physiology, (1988); 86(1): 134-136.
- Buschhaus C, Jetter R. Composition differences between epicuticular and intracuticular wax substructures: How do plants seal their epidermal surfaces? Journal of Experimental Botany, (2011); 62(3): 841-853.
- Khan MAU, Shahid AA, Rao AQ, Kiani S, Ashraf MA, et al. Role of epicuticular waxes in the susceptibility of cotton leaf curl virus (CLCuV). African Journal of Biotechnology, (2011); 10(77): 17868-17874.
- Baur P. Mechanistic aspects of foliar penetration of agrochemicals and the effect of adjuvants. Recent Research Developments in Agricultural and Food Chemistry, (1998); 2809-837.
- Kakani V, Reddy K, Zhao D, Mohammed A. Effects of ultraviolet‐B radiation on cotton (Gossypium hirsutum L.) morphology and anatomy. Annals of Botany, (2003); 91(7): 817-826.
- Carver TL, Gurr SJ. 12 Filamentous fungi on plant surfaces: Annual Plant Reviews, Biology of the Plant Cuticle. (2008); 23: 368. Wiley Publishers
- Hall D, Jones R. Physiological significance of surface wax on leaves. Nature, (1961); 191: 95 – 96.
- Gaume L, Perret P, Gorb E, Gorb S, Labat J-J, et al. How do plant waxes cause flies to slide? Experimental tests of wax-based trapping mechanisms in three pitfall carnivorous plants. Arthropod Structure & Development, (2004); 33(1): 103-111.
- Bullock JM, Federle W. The effect of surface roughness on claw and adhesive hair performance in the dock beetle Gastrophysa viridula. Insect Science, (2011); 18(3): 298-304.
- Eigenbrode S. The effects of plant epicuticular waxy blooms on attachment and effectiveness of predatory insects. Arthropod Structure & Development, (2004); 33(1): 91-102.
- Chang GC, Neufeld J, Durr D, Duetting PS, Eigenbrode SD. Waxy bloom in peas influences the performance and behavior of Aphidius ervi, a parasitoid of the pea aphid. Entomologia Experimentalis et Applicata, (2004); 110(3): 257-265.
- Mariani M, Wolters-Arts M. Complex waxes. The Plant Cell Online, (2000); 12(10): 1795-1798.
- Hannoufa A, McNevin J, Lemieux B. Epicuticular waxes of eceriferum mutants o Arabidopsis thaliana. Phytochemistry, (1993); 33(4): 851-855.
- Xia Y, Nikolau BJ, Schnable PS. Cloning and characterization of CER2, an Arabidopsis gene that affects cuticular wax accumulation. The Plant Cell Online, (1996); 8(8): 1291-1304.
- Oosterhuis D, Hampton R, Wullschleger S. Water deficit effects on the cotton leaf cuticle and the efficiency of defoliants. Journal of Production Agriculture, (1991); 4(2): 260-265.
- Bondada BR, Oosterhuis DM, Murphy JB, Kim KS. Effect of water stress on the epicuticular wax composition and ultrastructure of cotton Gossypium hirsutum L.) leaf, bract, and boll. Environmental and Experimental Botany, (1996); 36(1): 61-69.
- Barozai MYK, Husnain T. Development and characterization of the asiatic desi cotton (gossypium arboreum l.) leaf epicuticular wax mutants. Pakistan Journal of Botany, (2014); 46(2): 639-643.
- Dellaert L, Van Es J, Koornneef M. Eceriferum mutants in Arabidopsis thaliana (L.) Heynh. II. Phenotypic and genetic analysis. Arabidopsis Inforation Service, (1979); 16(1): 1.
- Rostás M, Ruf D, Zabka V, Hildebrandt U. Plant surface wax affects parasitoid’s response to host footprints. Naturwissenschaften, (2008); 95(10): 997-100.