

![]()
Tolerance of Roselle (Hibiscus sabdariffa L.) Genotypes to Drought Stress at Vegetative Stage
Bahaeldeen Babiker Mohamed1, 2, Muhammad Bilal Sarwar1, Sameera Hassan1, Bushra Rashid1*, Beenish Aftab1, Tayyab Husnain1
Adv. life sci., vol. 2, no. 2, pp. 74-82, February 2015
*- Corresponding Author: Dr. Bushra Rashid (Email: bushra.cemb@pu.edu.pk)
Author Affiliations[Date Received: 14/12/2014; Date Revised: 05/02/2015; Date Published Online: 25/02/2015]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Hibiscus sabdariffa L. is an important medicinal and fiber plant in Sudan. Among other stresses, drought extremely limits the growth, quality and net yield of the crop. The drought effects the crop plants by imposing certain morphological, physiological and biochemical changes at different periods of growth.
Methods: Current study was carried out in greenhouse settings at Center of Excellence in Molecular Biology (CEMB) to investigate the effects of drought stress. Five (5) different genotypes of Hibiscus sabdariffa L., namely Baladimostadir (H1), Um shiak (H2), Abu shankal (H3), Rahad mix (H4) and Abu Najma (H5) were studied. Thirty (30) days old Roselle seedlings were drought stressed for 10 days and its implications on plant growth, gas exchange, water relation, chlorophyll content and proline accumulation were estimated. Substantial genotypic differences in their adaptive response to drought were observed.
Results: Drought stress significantly affected the plant height; lowered the relative gas exchange efficiency and altered the physiological and biochemical responses. In comparison with others, H2 and H4 genotypes tolerated the osmotic stress well with lower osmotic potential and higher osmotic adjustment, better water content, higher stomatal conductance, photosynthetic efficiency and chlorophyll content. Accumulation of osmoprotectant and gas exchange indicators clearly distinguished the responses of different genotypes towards water stress.
Conclusion: Our results can be used for evaluation, screening, and manipulations of Hibiscus sabdariffa L. genotypes for improvement of drought tolerance through conventional breeding or drought responsive gene isolation.
Key words: Roselle, Drought stress, Hibiscus sabdariffa, Biochemical indicators, Physiological response
Introduction
Hibiscus sabdariffa L. belongs to Malvaceae family and has more than 1300 species grown all over the world dominanatly in tropical and semitropical areas [1]. It is mainly grown for its fleshy calyx (sepals) that is a good source of natural antioxidants (anthocyanins and protocatechuic acid) [2]. Abiotic stresses, including nutrient deficiency, salinity, chilling, freezing, extreme temperature, mineral toxicities and deficiencies are major limiting factors for plant’s growth and development [3]. Single or combination of these factors stimulate a range of physiological and biochemical responses in plants. These may include stomatal closure, repression in cell growth and photosynthesis, activation of respiration, accumulation of Marco molecules (proline) and antioxidants [4].
It has been documented that drought damages the crop yield especially in arid and semi-arid climate predominates in the range of 20-80% [5]. It is essential to identify the crop varieties or gene pool sources that are well adapted to stress environment. This study was designed to investigate the physiological and biochemical responses of different Sudanese Hibiscus Sabdariffa L. genotypes at initial growth stage, under drought stress.
Methods
Plant Material and growth condition
Seeds of the Roselle genotypes (Hibiscus sabdariffa L.) namely Baladimostadir (H1), Um shiak (H2), Abu shankal (H3), Rahad mix (H4) and Abu Najma (H5) were obtained from Agricultural Research Centre (ARC) and Alobied Research Station (ARS), Sudan. The experiment was conducted in green house at 30±2°C and 250-300 µmol m-2s1 of light intensity under complete randomized design with three replications. The drought was imposed on 30-days (DAS) old plants by withholding the water supply up to 10 days [6]. Well-watered plants were also maintained which were used as a control. Samples were taken from the control and stressed plants to analyze the morphological, physiological and biochemical response under drought stress.
Plant Height
Height of the control and water stressed plants was measured from the soil surface to the apex after 10 days of drought [7].
Osmotic Potential and Gas Exchange
Leaf osmotic potential was measured with pressure chamber (Plant Water Status Console-Model 3005-1412; Soil moisture Equipment Corp., Goleta, California, USA) [8]. Gas exchange parameters such as net photosynthesis rate (A), transpiration rate (E), and stomatal conductance (C), were measured with a handheld Infrared Gas Analyzer (IRGA), gas exchange system (CI-340 Bioscientific Ltd., UK) from drought stressed and control plants [9]. All these measurements were recorded in triplicates from each genotype in the mid-day sun shine.
Leaf Relative Water Content (LRWC)
LRWC was determined from leaves using the procedure described by Shaheen and Shahbaz [10] with little modifications. Five fully developed young leaves (1g) of uniform size were selected from each treatment and fresh weight (FW) of each sample was recorded. The leaf samples were then immersed in double distilled water (ddH2O) for 24 hours (h) to determine the turgor weight (TW). Then the samples were oven-dried at 80˚C for 24 h to determine the dry weight (DW). The LRWC was determined using the following formula;
LRWC = (Fresh Weight – Dry Weight ⁄ Turgor Weight – Dry Weight) × 100
Chlorophyll Content
The photosynthetic pigments were determined as described by Arnon and Whatley [11]. Chlorophyll extract was prepared from 100 mg of fresh leaves by grinding with 10 mL of 80% acetone. The homogenate was left overnight at room temperature under dark condition. Absorbance of the extract was read at 663 nm and 645 nm. The concentration of chlorophyll a, b and total chlorophyll was calculated using Arnon’s equations.
Proline Content
The proline content from leaves of Hibiscus sabdariffa genotypes were estimated under control and drought stress conditions by following the Bates et al., method [12]. For this estimation leaf sample of about 0.5g was taken and processed as above mentioned method. Finaly, the absorbance of the mixture was measured at 520nm in a spectrophotometer (spectra Max plus: molecular devices, USA). A blank was maintained while taking absorbance for all the samples. Standard curve was obtained by known concentrations of authentic L-proline and sample values were calculated as µgg-1 of fresh leaf tissue.
Statistical analysis
All experimental data are the means of at least three independent replicates, and results were determined using analysis of variance (ANOVA) via Statistic software. Variation among treatment means were compared using least significant difference (LSD) test (P<0.05).
Results
Plant Height
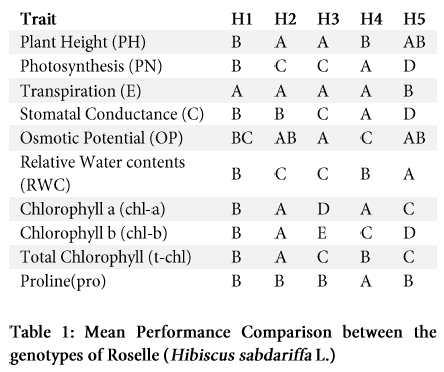
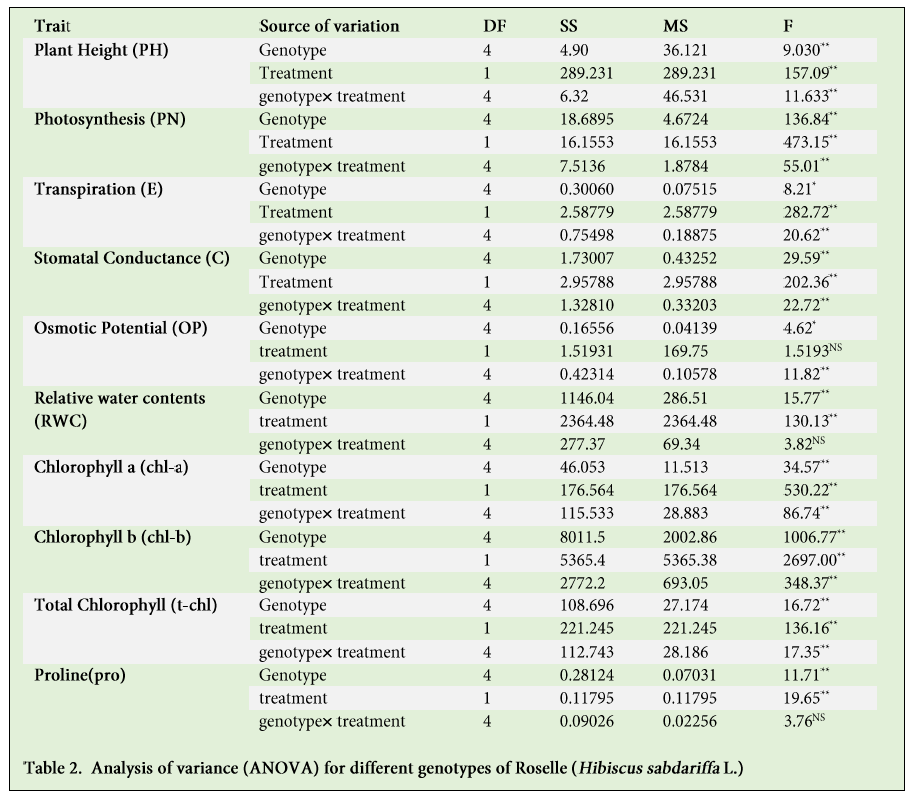
Overall plant growth was severely reduced due to the drought stress. We observed significant inhibitory effect on the growth of different genotypes growing under drought, when compared to the control plants (Table 1 and 2). Stressed plants showed significantly lower plant height than the control plants in H1, H2, H3, H5, while no significant (P >0.05) difference were found related to plant height in H4 genotype as shown in Figures 1 and 2.
Osmotic potential
Increasing osmotic stress or salt concentration generally trim down water related attributes due to declining soil water potential (Ψs), creating a water stress for the plant, and ionic effects due to ion uptake and/or accumulation. Graphical representation of data indicates that there is a reduction in the leaf osmotic potential (ψ) under drought stress. In control group, osmotic potential was observed 1.23, 1.07, 1.04, 1.06, and 0.99 (-MPa). While in H1, H2, H3, H4 and H5 genotypes it declined to 1.32, 1.637, 1.47, 1.8 and 1.377 (-MPa) respectively after the application of drought stress (Figure 3). Osmotic potential was reduced among all the genotypes, showing the capacity of osmotic adjustment efficiency. Genotypes H4 and H2 exhibited the lowest osmotic potential (ψs) (-68.4 and -51.79%) with greatest osmotic adjustment while genotypes H1, H3 and H5 showed comparatively lower (-6.56, -40.28 and -38.44%) insignificant osmotic potential and osmotic adjustment ability respectively (Figure 3; Table 1).
Infrared gas analyzer (IRGA)
Our study pointed out the significant impact of drought stress on physiological parameters of different genotypes of Hibiscus sabdariffa L. However, the difference among the genotypes mean performance does not reach the significance level (Table1 and 2). All the genotypes showed a variable response to photosynthetic rate (PN) under control and stress condition and a considerable reduction in photosynthetic process in all genotypes except H4 under drought (Figure 4A).
After the drought stress, carbon dioxide assimilation rate (stomatal conductance; C) was significantly decreased in all the genotypes. That was maximum for H1 and H2 followed by the H4, H3 and H5 genotypes in control condition (Figure 4B). This was reduced in H4, H3 and H2 respectively under drought, but was reduced more in H1 as compared to the control condition.
Transpiration rate (E) was significantly reduced in all the genotypes except H5 due to water stress (Figure 4C). This value was higher in H1, H2 and H4 genotypes and was lowest in H3 and H4 relatively under the control environment. Net photosynthesis was decreased by 13.95, 37.73, 23.25, 21.73 and 33.3%; stomatal conductance was decreased by 55, 25, -10, 32 and -18.18%; and transpiration decreased by 35.71, 47.05, -25, 15.38, and -45.32% respectively, in H1, H2, H3, H4 and H5 respectively under drought stress condition.
Leaf Relative water content (LRWC)
Stomatal opening and photosynthesis are mainly affected by the overall status of leaf water availability. Significant differences in LRWC were observed among the genotypes under osmotic stress condition (Table 1 and 2). The water content in leaves of untreated plants nearly remained constant throughout the experiment, whereas LRWC after 10 days of water stress were 63, 62, 53, 87 and 85% in genotypes H1, H2, H3, H4 and H5 respectively. Maximum reduction of water content was recorded in H3 genotype (<52%) while H1 and H4 genotypes held the water content above average (>76 and 80%) respectively (Figure 5). ANOVA and comparative mean study indicates variations in LRWC and highly significant impact of drought stress in all the genotypes (Table 2), while Table 1 indicates the individual mean performance for positive correlation between LRWC and drought stress.
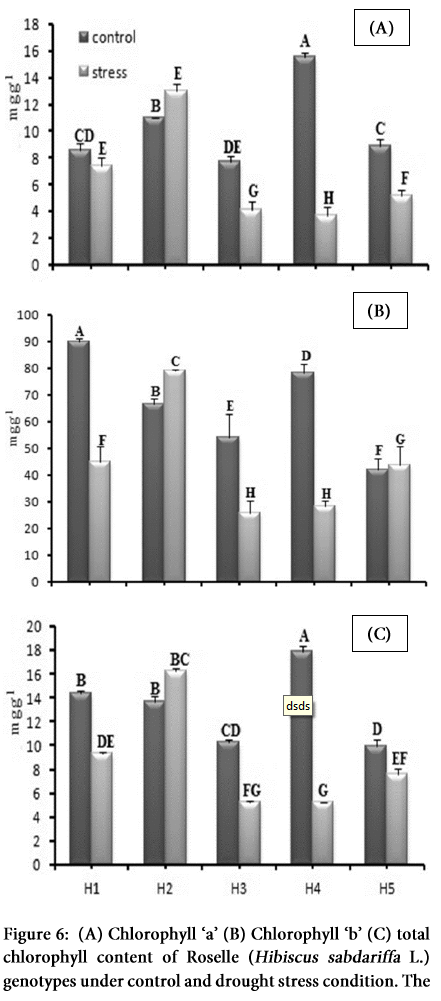
Chlorophyll content
In this study, significant impact of osmotic stress was observed on leaf chlorophyll content in all the genotypes (p>0.01) (Table 1 and 2), however, H2 genotype performed better and showed the highest values for chlorophyll content, which indicates its comparative effective photosynthesis performance against drought stress. Else, the genotype H4 and H5 showed lower chlorophyll content under stress conditions that significantly limit their photosynthesis efficiency. Under drought stress, total chlorophyll content decreased in H1 (35%), H3 (49%), H4 (70%) and in MCC877 (23%) and increased in H2 (by 18%) (Figure 6 A-C). Statistical analysis of chlorophyll a, b and total chlorophyll showed the significant effects (P >0.01) under water stress (Table 2). Chlorophyll a and b showed marked reduction except in H2 and H5 genotypes.
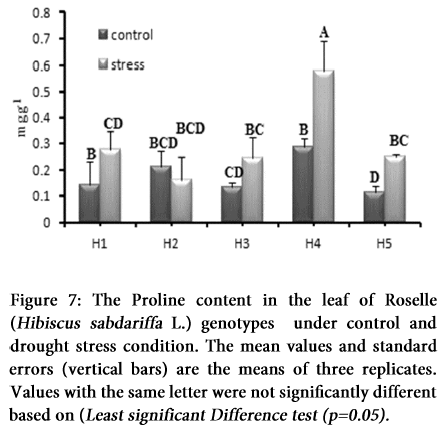
Proline content
There was observed substantial accumulation of endogenous proline under moisture deficiency in all the genotypes except the H2 (Table 2). Highest accretion was observed in H4 (6 mgg-1) followed by the H1, H3 and H5 genotypes under drought stress (Figure 7). Statistically, mean performance of individual genotypes except H4, exhibited non-significant pattern of accumulation of proline with respect to control or stress condition (Table 1).
Figures & Tables
Figure 1 Figure 2 Figure 3 Figure 4
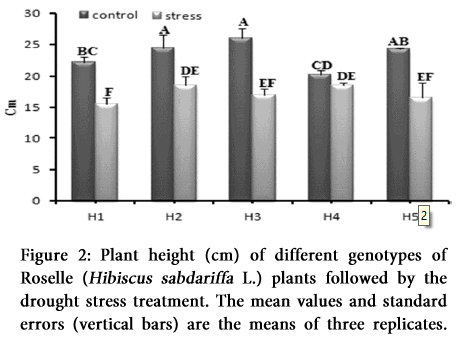
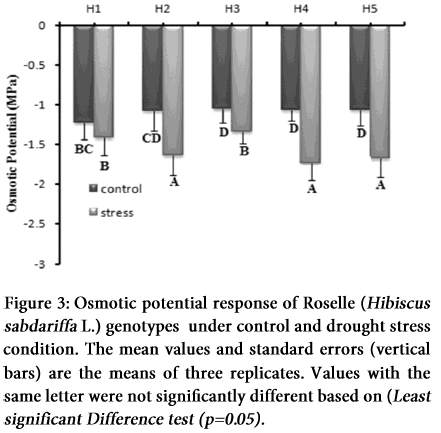
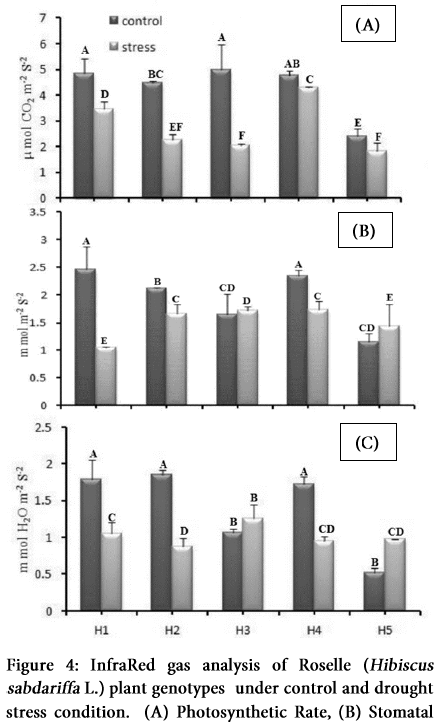
Figure 5 Figure 6 Figure 7 Table 1 Table 2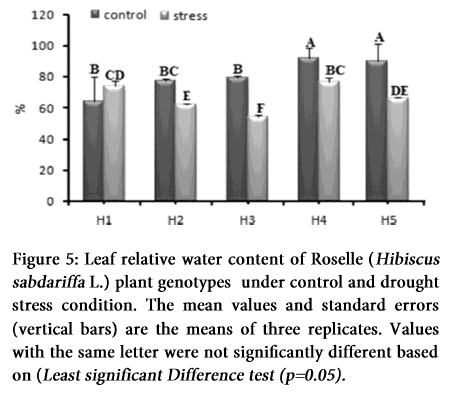
Discussion
Drought is one of the major limiting factors for high productivity and crop yield. This study has been carried out to evaluate the effect of drought on different genotypes of an important crop Hibiscus sabdariffa. Plants undergo a number of physiological and biochemical variations under the drought stress and these mechanisms are altered to adapt the plants under such circumstances. These factors also affect the morphological growth of the plants such as plant growth and development, biomass and the yield [4].
Under any abiotic stress plant undergo osmoregulation by accumulation of solutes within the cells. Comparative analysis of mean values of water stressed and control treatments pointed out that, the drought stress induced a significant cut in osmotic potential.
The decrease in osmotic potential is considered as a potential cellular mechanism of drought tolerance, as it enables turgor maintenance and growth continuation [13]. In this experiment, H4 genotype exhibited low osmotic potential under moisture deficient condition and thus it is considered to be a drought responsive genotype with high osmotic adjustment than the others. However, it also presents a metabolic cost due to synthesis and compartmentation of osmolytes [14]. The low initial ψs can act as a pre-existing force to immediate dehydration buffering which was well documented in one earlier study [15]. A Similar response to osmotic stress was observed in previous study [16] in drought tolerant and sensitive wheat genotypes, olives [17] and Quercus crispula [18].
Decrease in photosynthesis rate is considered among the main factors in growth reduction and yield of plants under drought stress. Possible repression of photosynthetic process may link with alterations in carbon and nitrogen incorporation. The plants respond to water deficit with a rapid closure of stomata to avoid further loss of water through transpiration. As a consequence, the diffusion of CO2 into the leaf is restricted. Occasionally plant water usage efficiency is enhanced due to minor decline in stomatal conductance under mild stress. Low availability of water under drought environment, restricts the movement of water within the plants at organ, tissues or cellular level, leads to closure of stomata [19]. Therefore, it could be hypothesized that stomatal conductance is maintained under lower water potential in the tolerant genotypes [20] and the plants saved the energy and maintained their physio-chemical processes, even after observing the decline in growth and development. This study is in coherence with the preceding reports which suggest that water deficiency induces the reduced photosynthesis (Pn) associated with the other physiological markers, i.e. C and E [21]. However, the degree of reduction differed between the genotypes and species. Severe reduction in the overall PN, C and E may indicate the sensitivity of genotypes to the drought stress; while tolerant or resistant genotypes showed lower rate of reduction in these processes [22].
The overall leaf water content of higher plants under reduced soil moisture is of great importance. Decrease in the LRWC as in response to drought stress has been noted in a wide variety of plants [23-24]. This variation in water holding capacity of genotypes may be attributed to their differential ability to absorb water content from the soil by developing a lower water potential gradient from soil. It may also be due to the difference in the ability of the genotypes to adjustment and maintain osmotic turgor in tissues and hence physiological activities as displayed by genotype H4.
Similar to other factors, stable chlorophyll content during drought is a desirable trait viewed as one of the criteria to decide the tolerance [25, 26]. Some genotypes of sesame and onion subjected to drought stress, initially increased the leaf chlorophyll and then remained unchanged [27, 28]. It has been documented that abiotic stress negatively affects the activity of photosynthetic enzymes, chlorophyll and carotenoids [29]. Increased chlorophyll content under drought stress may be related to a decrease in leaf area and that might be a defensive response to reduce the harmful effects of drought.
Plants respond to a variety of stresses by accumulating certain specific metabolites or amino acids such as proline. Enhanced proline content under water stress is a principle for adaptation of tolerant genotypes to overcome the stress [30]. Statistically mean performance of individual genotypes except H4, exhibited non-significant pattern of accumulation of proline with respect to control or stress condition. This is similar to the previous reports for other crops such as barley [31] and wheat [32]. Proline accumulation in stressed plants has been well established mechanism that plays a key role in osmoregulation defense mechanism, leading to prevent the cell osmotic pressure and survival in the extreme conditions.
In conclusion, Hibiscus Sabdariffa L. genotypes, showed varied response to drought stress for morphological, physiological and biochemical characteristics. This indicates the diverse genetic makeup of the genotypes and confirms the genetically controlled response to the drought stress. Genotypes H2 and H4 were adapted well to drought by modification in various physiological activities such as reduced water losse that make them suitable for cultivation in water deficient areas. Further study at molecular level is necessary for identification, isolation and characterization of the drought responsive genes under drought stress.
Acknowledgements
Authors are thankful to the Third World Academy of Sciences (TWAS) for provision of funds.
References
- Keshavarzi MHB, Moussavinik SM. The effect of different NaCl concentration on germination and early growth of Hibiscus sabdariffa seedling. Annals of Biological Research, (2011); 2(4): 143-149.
- Ali HM, Siddiqui MH, Basalah MO, Al-Whaibi MH, Sakran AM, et al. Effects of gibberellic acid on growth and photosynthetic pigments of Hibiscus sabdariffa L. under salt stress. African Journal of Biotechnology, (2012); 11(4): 800-804.
- Vinocur B, Altman A. Recent advances in engineering plant tolerance to abiotic stress: achievements and limitations. Current opinion in biotechnology, (2005); 16(2): 123-132.
- Shinozaki K, Yamaguchi-Shinozaki K. Gene networks involved in drought stress response and tolerance. Journal of Experimental Botany, (2007); 58(2): 221-227.
- Iqbal MA, Iqbal A. A Study on Dwindling Agricultural Water Availability in Irrigated Plains of Pakistan and Drip Irrigation as a Future Life Line. American-Eurasian Journal of Agricultural & Environmental Sciences, (2015); 15(2): 184-190.
- Silvente S, Sobolev A, Lara M. Metabolite adjustments in drought tolerant and sensitive soybean genotypes in response to water stress. PloS One, (2012); 7(6): e38554.
- Hassan S, Sarwar MB, Sadique S, Rashid B, Aftab B, et al. Growth, Physiological and Molecular Responses of Cotton (Gossypium arboreum L.) under NaCl Stress. American Journal of Plant Sciences, (2014); 5(5): 605-614.
- Mao X, Zhang H, Tian S, Chang X, Jing R. TaSnRK2.4, an SNF1-type serine/threonine protein kinase of wheat (Triticum aestivum L.), confers enhanced multistress tolerance in Arabidopsis. Journal of Experimental Botany, (2010); 61(3): 683-696.
- Akram M, Ashraf M, Jamil M, Iqbal R, Nafees M, et al. Nitrogen application improves gas exchange characteristics and chlorophyll fluorescence in maize hybrids under salinity conditions. Russian Journal of Plant Physiology, (2011); 58(3): 394-401.
- Shaheen HL, Shahbaz M. Salt-induced effects on some key morpho-physiological attributes of cotton (Gossypium hirsutum L.) at various growth stages. Soil and Environment, (2012); h31(2): 125-133.
- Arnon D, Whatley F. Is chloride a coenzyme of photosynthesis? Science (1949); 110(2865): 554-556.
- Bates L, Waldren R, Teare I. Rapid determination of free proline for water-stress studies. Plant and Soil, (1973); 39(1): 205-207.
- Munns R. Comparative physiology of salt and water stress. Plant, Cell & Environment, (2002); 25(2): 239-250.
- Bajji M, Lutts S, Kinet JM. Physiological Changes after Exposure to and Recovery from Polyethylene Glycol-induced Water Deficit in Callus Cultures Issued from Durum Wheat (Triticum durum Desf.) Cultivars Differing in Drought Resistance. Journal of Plant Physiology, (2000); 156: 75-83.
- Mohamed FM, Azza AT. Dehydration-induced alterations in growth and osmotic potential of callus from six tepary bean lines varying in drought resistance. Plant Cell, Tissue and Organ Culture, (2006); 87: 255-262.
- Gupta NK, Sunita G, Arvind K. Effect of Water Stress on Physiological Attributes and their Relationship with Growth and Yield of Wheat Cultivars at Different Stages. Journal of Agronomy and Crop Science, (2001); 186(1): 55-62.
- Dichio B, Xiloyannis C, Sofo A, Montanaro G. Osmotic regulation in leaves and roots of olive trees during a water deficit and rewatering. Tree Physiology, (2006); 26(2): 179-185.
- Saito T, Terashima I . Reversible decreases in the bulk elastic modulus of mature leaves of deciduous Quercus species subjected to two drought treatments. Plant, Cell and Environment, (2004); 27(7): 863-875.
- Robinson M, Very A, Sanders D. How can stomata contribute to salt tolerance? Annals of Botany, (1997); 80(4): 387-393.
- Lidon Z, Cebola F. An overview on drought induced changes in plant growth, water relations and photosynthesis. Emirates Journal of Food and Agriculture, (2012); 24(1): 57-72.
- Chaves MM, Maroco JP, Pereira JS. Understanding plant responses to drought from genes to the whole plant. Functional Plant Biology, (2003); 30(3): 239-264.
- Silim S, Nash R, Reynard D, White B, Schroeder W. Leaf gas exchange and water potential responses to drought in nine poplar (Populus spp.) clones with contrasting drought tolerance. Trees, (2009); 23: 959-969.
- Siddique M, Hamid A, Islam M. Drought stress effects on water relations of wheat. Botanical Bulletin of Academia Sinica, (2000); 41: 35-39.
- Nayyar H, Gupta D. Differential sensitivity of C3 and C4 plants to water deficit stress: Association with oxidative stress and antioxidants. Environmental and Experimental Botany, (2006); 58(1–3): 106-113.
- Long S, Bernacchi C. Gas exchange measurements, what can they tell us about the underlying limitations to photosynthesis? Procedures and sources of error. Journal of Experimental Botany, (2003); 54(392): 2393-2401.
- Sairam RK. Effects of homobrassinolide application on plant metabolism and grain yield under irrigated and moisture-stress conditions of two wheat varieties. Plant Growth Regulation, (1994); 14: 173-181.
- Mensah J, Obadoni B, Eruotor P, Onome-Irieguna F. Simulated flooding and drought effects on germination, growth, and yield parameters of sesame (Sesamum indicum L.). African Journal of Biotechnology, (2009); 5(13): 1249-1253.
- Saeidi M, Zabihi-e-Mahmoodabad R. Evaluation of drought stress on relative water content and chlorophyll content of sesame (Sesamum indicum L.) genotypes at early flowering stage. Research Journal of Environmental Sciences, (2009); 3(3): 345-350.
- Stępień P, Kłbus G. Water relations and photosynthesis in Cucumis sativus L. leaves under salt stress. Biologia Plantarum, (2006); 50610-616.
- Sarwar MB, Batool F, Rashid B, Aftab B, Hassan S, et al. Integration and expression of heat shock protein gene in segregating population of transgenic cotton plant for drought tolerance. Pakistan Journal of Agricultural Sciences, (2014); 51(4): 935-941.
- Thameur A, Ferchichi A, López-Carbonell M. Quantification of free and conjugated abscisic acid in five genotypes of barley (Hordeum vulgare L.) under water stress conditions. South African Journal of Botany, (2011); 77: 222-228.
- Hong-Bo S, Xiao-Yan C, Li-Ye C, Xi-Ning Z, Gang W, et al. Investigation on the relationship of proline with wheat anti-drought under soil water deficits. Colloids and surfaces B, Biointerfaces, (2006); 53(1): 113-119.