

![]()
Analysis of reporter proteins GUS and DsRed driven under the control of CaMV35S promoter in syncytia induced by beet cyst nematode Heterodera schachtii in Arabidopsis roots
Muhammad Amjad Ali1,2*, Amjad Abbas1
Adv. life sci., vol. 3, no. 3, pp. 89-96, May 2016
*- Corresponding Author: Muhammad Amjad Ali, (Email: Amjad.Ali@uaf.edu.pk)
Authors' Affiliation
2- Centre of Agricultural Biochemistry and Biotechnology (CABB), University of Agriculture, 38040 Faisalabad – Pakistan
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Cyst nematodes induce specialized feeding structures called syncytia in the plant roots. The expression of CaMV promoter in syncytia has remained topic of debate. The objective of this research was to study the activity of CaMV promoter by using reporter proteins like GUS and DsRed under the control of CaMV35S promoter in syncytia induced by H. schachtii in Arabidopsis roots.
Methods: pMAA-Red and pPZP3425 plasmids were used to study expression of GUS and DsRed in syncytia. The plants were grown in 2% Knop medium under sterile conditions in growth chambers at 25°C in long day conditions. GUS activity in syncytia was studied through staining of syncytia using X-gluc solution. Ds-Red fluorescence in syncytia was detected by using an inverse microscope equipped with UV filter.
Results: The expression analysis of DsRed protein driven by CaMV promoter demonstrated that this promoter is active in syncytia at all the time points. All the syncytia showed DsRed expression at 5 dpi. At 7 dpi, 10 dpi and 15 dpi over 90%, 80% and 50% of the syncytia showed DsRed fluorescence respectively. There was very high fluorescence in the syncytia as compared to the uninfected root segments due to high expression. CaMV::GUS lines showed GUS expression in 80% of 5dpi syncytia. However, unlike expression of DsRed, the number of GUS stained syncytia decreased quickly to around 50% at 7 dpi and to about 5% in the 15 dpi syncytia.
Conclusions: The results conclude that CaMV promoter is more active in younger syncytia as compared to older syncytia but can be used for expression in syncytia. Moreover, DsRed protein could be used as better reporter for evaluation of gene expression in syncytia as compared to GUS.
Keywords: Reporter genes, CaMV35S promoter, GUS, Ds-Red, H. schachtii, Arabidopsis, Syncytia
Introduction
Plant parasitic nematodes have a detrimental effect on agricultural productivity either by a direct damage to crops or serving as vectors for plant invading viruses, thereby resulting in over $150 billion crop losses worldwide annually [1]. The cyst and root-knot nematodes within the family Heteroderidae are the most important plant parasitic species and are obligate endoparasites of plant roots. Cyst nematodes with their destructive migration activity travel intracellularly through the root to reach the elongation zone, where they select a cell at the periphery of the vascular bundle to become the initial syncytial cell. This cell expands through partial dissolution of cell walls of up to 200 times of the surrounding cells into a feeding site known as a syncytium [2,3]. Due to incorporation of many surrounding cells the syncytium becomes multinucleate and substantial hypertrophy of the syncytial elements results in enlargement of the feeding site (Figure 1A). This syncytium becomes the only nutrient source for the nematodes as they develop through subsequent sedentary life stages [4,5]. The common features of a syncytium include high metabolic activity, disintegration of the central vacuole, reduced number of secondary vacuoles, and increased numbers of organelles and wall ingrowths into xylem vessels [6,7]. The females of cyst nematode are fertilized by vermiform adult males. The male nematodes induce comparatively smaller syncytia than females and are covered in a cuticular casing (Figure 1B). The males die after fertilization of several females while the female dies after the completion of egg development, leaving the eggs contained within its enlarged body [8] (Figure 1A). The female mostly detaches from the root surface and hardens to form a protective cyst covering hundreds of eggs inside. The cyst with eggs can remain viable for many years in the soil before infective J2s hatch in favorable conditions to start a new life cycle [9].
The cauliflower mosaic virus 35S (CaMV35S) promoter has been extensively used for the constitutive expression of transgenes in many plant species [10,11]. A combination of CaMV35S promoter with omega element from Tobacco Mosaic Virus (TMV) used as translational enhancer resulted in enhanced expression of the gene of interest delivered with this promoter [12]. Holtorf et al. [13] compared different constitutive promoters (CaMV35S, CaMV35S+omega, UBQ1 and BTH6) and one inducible promoter (soybean heat-shock promoter Gmhsp17.3) and found that CaMV35S+omega is a very strong promoter for constitutive expression in all organs of Arabidopsis thaliana.
The CaMV35S promoter has been used to express a variety of reporter proteins such as GUS [14], Luc [15], GFP [16] or DsRed [17]. The DsRed protein was initially isolated from a sea anemone, Discosoma striata and is responsible for the red colour around the oral disk of a coral of the genus Discosoma [17]. The DsRed protein has a molecular weight of 28 kDa and a maximum emission wavelength of 583 nm and it emits a bright red fluorescent light [18]. Jach et al. [19] first used DsRed delivered by the CamV35S promoter as a reporter for plant gene expression in tobacco and concluded that it is well suited, easy-to detect reporter protein for plant molecular biology instead of GUS, Luc or GFP. Very recently, we have used DsRed successfully as a fluorescent selectable marker for the selection of transgenic lines [20].
The objective of this study was to trace out the expression of CaMV35S promoter which contains two time 35S (CaMV2x35S) region plus omega element in the end of the CaMV35S sequence (Figure 2) by using DsRed and GUS. This study was carried out because of the debate about the activity or expression of CaMV35S promoter in the cyst nematode induced syncytia. For example, Urwin et al. [21] reported that CaMV35S is active in giant cells induced by root-knot nematode (M. incognita) but it was highly downregulated in syncytia induced by cyst nematodes (H. schachtii). Liu et al. [22] showed that a chemoreception-disruptive peptide delivered by CaMV35S considerably inhibits acetylcholinesterase in Globodera pallida induced syncytia in potato. Most of the reports have shown that this promoter is downregulated in cyst nematode induced syncytia [23,24] while Urwin et al. [25] demonstrated that CaMV35S is active in syncytia up to 10 dpi (days post inoculation). However, Goverse et al. [26] used green fluorescent protein (GFP) to monitor the activity of two constitutive promoters, CaMV35S and TR2´, in developing syncytia induced by Globodera rostochiensis in potato roots and found that CaMV35S is upregulated in syncytia until 13dpi. Similarly, CaMV35S has been used to suppress nematode development by overexpressing nematode repellent as well as nematode specific genes involved in nematode establishment on the plants [27,28]. But there is still a debate about the activity of CaMV35S promoter in the cyst nematode induced syncytia. We have constructed CaMV35S promoter:: reporter fusion lines to find out the expression of this promoter using two reporter proteins GUS and DsRed.
Methods
Construction of plasmid vectors CaMV2x35S + omega:: GUS/DsRed fusions
For testing the expression of CaMV promoter in syncytia, promoter::reporter fusion lines were developed using two reporters, GUS (β-glucuronidase) [14] and DsRed [17]. For the promoter::GUS construct we used the vector pMAA-Red which had a CaMV2x35S+omega::GUS cassette. From the pMAA-Red construct, first three lines with maximum expression of GUS from the previous study were used for GUS staining in syncytia at different time points [14]. For the construction of CaMV::DsRed, pPZP3425 [29] was used by replacing GUS with the DsRed sequence as given by Ali et al. [20].
Plant transformation and selection of transgenic lines
The plasmid constructs containing CaMV::GUS (pMAA-Red) and CaMV::DsRed (pPZP3425) fusions were introduced into A. tumefaciens GV3101 for transformation of Arabidopsis plants by the floral dip method [30]. In case of pMAA-Red, the fluorescent transformed seeds were selected under an inverse microscope (Axiovert 200M; Zeiss, Hallerbergmoos, Germany) equipped with a DsRed fluorescence filter and put on soil to grow the next generation. Homozygous lines were selected based on visual observation as described by Ali et al. [20].
In case of pPZP3425 containing CaMV::DsRed cassette, the transgenic lines were selected on MS medium containing 3% sucrose (Murashige and Skoog, 1962) supplemented with 50 mM kanamycin as described by Szakasits et al. [29]. In this case three transgenic lines were selected for further studies.
Plant cultivation
Arabidopsis thaliana (L.) ecotype Columbia plants were grown on soil in growth chambers at 25°C in long day conditions (16 h light / 8 hour dark). For growth in sterile conditions seeds were surface sterilized for 7 min in 10% (w/v) sodium hypochlorite and subsequently washed three times with sterile water. Seeds were placed in Petri dishes (9 cm) on a modified Knop medium with 2% sucrose [31] and grown for 12 days for nematode infection.
Nematode infection
The cysts of the sugar beet cyst nematode Heterodera schachtii were harvested from in vitro stock cultures propagated on mustard (Sinapsis alba cv. Albatros) roots growing on 0.2 concentrated Knop medium supplemented with 2% sucrose [31]. The cysts were soaked in 3 mM ZnCl2 as stimulus for hatching of J2 larvae under sterile conditions. The J2 larvae were then washed three times in sterile water and resuspended in 0.5% (w/v) Gelrite (Duchefa, Haarlem, The Netherlands) before inoculation. Roots of twelve-day-old Arabidopsis seedlings were inoculated under sterile conditions with about 50-60 juveniles per plant.
Histochemical detection of GUS activity
Histochemical detection of GUS activity was performed by staining [14] using X-gluc (Biomol, Hamburg, Germany) in 0.1 M sodium phosphate buffer pH 7.0 containing 0.1% Triton-X 100, 0.5 mM K3[Fe(CN)6], 0.5 mM K4[Fe(CN)6] and 10 mM Na2EDTA. For GUS staining of syncytia, the infected roots of CaMV::GUS plants were incubated with X-gluc for 6 hours at 37ºC. Staining was performed at 5, 7, 10, and 15 dpi (days post inoculation). Stained syncytia and uninfected roots were counted per line from three promoter::GUS lines and photographed under an inverse microscope (Axiovert 200M; Zeiss, Hallerbergmoos, Germany) having an integrated camera (AxioCam MRc5; Zeiss). The GUS expression in the root cap border cells was also photographed in the same way.
Detection of DsRed fluorescence in syncytia
Three lines of the CaMV::DsRed construct were grown on Knop medium and infected with nematodes as mentioned before. The number of syncytia having DsRed expression were counted and imaged at 5, 7, 10 and 15 dpi using an inverse microscope (Axiovert 200M; Zeiss, Hallerbergmoos, Germany) having a DsRed filter and an integrated camera (AxioCam MRc5; Zeiss). The DsRed expression in the root cap border cells was also photographed in the same way.
Results
Expression of reporter genes driven by the CaMV2x35S + omega promoter in the syncytia
The CaMV promoter that was used here contained the omega element as described in [13] and a doubled enhancer [32]. The sequence of CaMV promoter with two copies of 35S region along with omega element is shown in Figure 2. In order to test the activity of CaMV promoter in syncytia, we constructed promoter fusion lines with beta-glucuronidase (GUS) and DsRed florescent protein. For construction of CaMV.2x35S + omega::DsRed we used pPZP3425 [29] in which GUS was replaced by DsRed. The transgenic lines used for the study were having the highest fluorescence in the seeds. For CaMV.2x35S+omega::GUS we used the pMAA-Red vector itself [20].
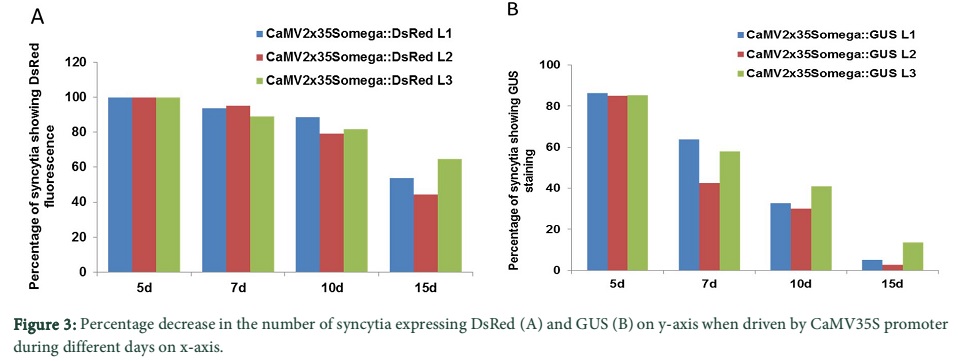
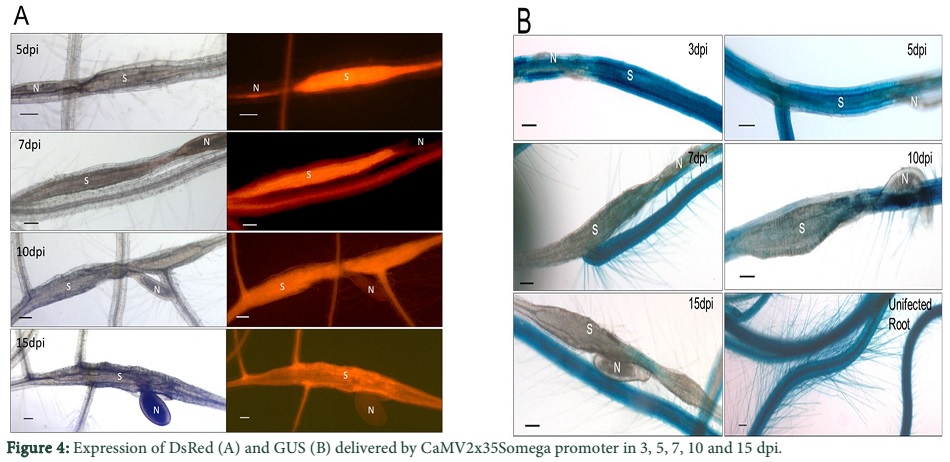
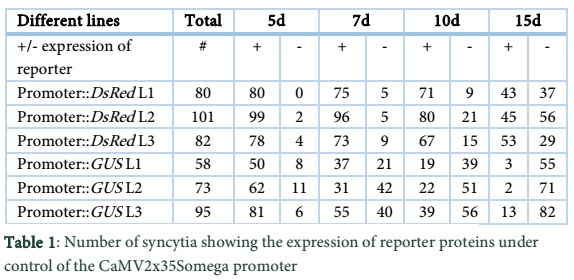
We determined the expression of DsRed and GUS driven by the CaMV promoter at different time points after infection. The lines, CaMV.2x35S + omega::DsRed L1, L2 and L3 contained 80, 101 and 82 syncytia in total, respectively and all of them showed DsRed fluorescence at 5 dpi (Table 1). The percentage decrease of total number of syncytia in fluorescent syncytia at different time points is given in Figure 3A. At 7 dpi, over 90% of the total syncytia were found fluorescent. Similarly, at 10 dpi and 15 dpi on an average from three lines over 80% and 50% of the syncytia showed DsRed fluorescence respectively (Figure 3A). Representative syncytia with DsRed fluorescence were also photographed at different time points (5, 7, 10 and 15 dpi) and are shown in Figure 4A. It is quite obvious from the figure that there was very high fluorescence in the syncytia as compared to the uninfected root segments. However, at 15 dpi, the expression level of DsRed also slightly went down showing decreased activity of the promoter with the passage of time.
For the CaMV.2x35S + omega::GUS construct we used the first 3 transgenic lines harboring the pMAA-Red vector having the highest expression in Ali et al. [20]. The pMAA-Red seedlings were grown on MS medium for 14 days and stained with X-gluc solution (see Materials and Methods) at 37ºC for 6 hours. The seedlings had very high staining, showing that the CaMV.2x35S + omega promoter was highly and constitutively active in the seedlings under the control of this promoter (data not shown). After nematode infection, the transgenic lines CaMV.2x35S + omega::GUS L1, L2 and L3 contained 58, 73 and 95 syncytia in total, respectively and over 80% of them showed GUS staining in syncytia after five days of infection (Table 1 and Figure 3B).
However, unlike expression of DsRed, the number of GUS stained syncytia decreased quickly to around 50% at 7 dpi and to about 5% in the 15 dpi syncytia (Figure 3B). Representative syncytia with GUS staining at different time points (3, 5, 7, 10 and 15 dpi) were photographed and are shown in Figure 4B. The syncytia at 3 and 5 dpi showed strong staining while the GUS staining in case of 7, 10 and 15 dpi was more confined towards the region of nematode attachment with the root. Nevertheless, at 15 dpi, the GUS staining was very low and only found at the point of nematode attachment, indicating that the activity of the CaMV.2x35S + omega promoter decreases with the passage of time in older syncytia.
Expression of GUS/DsRed driven by CaMV.2x35S + omega promoter in root cap border cells
As this promoter was constitutively active throughout the plant tissues, the expression of both the reporter genes was found in the root cap border cells of the root tips. The CaMV.2x35S + omega:: DsRed as well as the GUS cassettes showed high expression in the root cap border cells as shown in Figure 5A and B respectively. All the transgenic lines showed the same expression of both the reporters in the root cap border cells, however, only the representative photos are shown here.
Data and Tables



Discussion
Our results showed that the CaMV.2x35S+omega promoter was active at all the time points in case of DsRed with decreased expression only at 15 dpi. However, in case of GUS, the promoter was active in syncytia at 3, 5 and 7 dpi and was slightly downregulated at 10 dpi and almost inactive at 15 dpi. The molecular weight of GUS gene (UidA) based on the amino acid mentioned by Ali et al. [20] in Genbank is just over 68 KiloDaltons (kDa) which is higher that of DsRed protein is around 25 kDa but this is not the molecular weight difference that the promoter delivered the DsRed (smaller protein) more efficiently into the syncytia as compared to GUS. What could be the reason for the better expression of DsRed in older syncytia as compared to GUS? DsRed is a fluorescent protein which does not need any cofactor or substrate which is needed for GUS staining (i.e. X-gluc) [14]. The substrate thus has to get into the syncytia to be available for the GUS enzyme to interact. Old syncytia have very strong outer cell walls which could make it difficult for the GUS substrate to penetrate into the old syncytia. Similar observations were reported by Goverse et al. [26] who expressed green fluorescent protein (GFP) in transgenic potato (Solanum tuberosum) plants under the control of the CaMV35S promoter. By using confocal laser scanning microscopy (CLSM), they investigated promoter activity in syncytia during potato cyst nematode (Globodera rostochiensis) infection and observed GFP expression up to 13 dpi [26].
The CaMV.2x35S+omega promoter was used to overexpress some defense related genes like RAP2.6 and WRKY33 in syncytia during our studies and we found elevated expression of these genes in syncytia which resulted in enhanced nematode resistance in transgenic lines [20,33,34]. This suggested that this promoter could be used to express the gene of interest constitutively in plants, including syncytia. The CaMV promoter has been used for proteins to increase resistance against nematodes. For example, Liu et al. [22] successfully used the CaMV35S promoter for expressing synthetic chemodisruptive peptides in planta which disrupted the establishment of cyst nematode Globodera pallida in potato. Similarly, the CaMV35S promoter was used to overexpress different proteinase inhibitors which resulted in lower numbers of nematode with impaired development on several host plants including potato, rice and banana (Reviewed by [27]). This promoter have also been used to control the expression of RNAi or double stranded RNA of nematode parasitism genes to induce nematode resistance mediated by RNA interference [35]. However, Urwin et al. [21] indicated that CaMV35S is active in giant cells induced by the root-knot nematode Meloidogyne incognita but was highly downregulated in syncytia induced by cyst nematodes (H. schachtii). It has been shown in many studies that CaMV35S promoter is downregulated in cyst nematode induced syncytia [23,24] while Urwin et al. [25] concluded that CaMV35S is active in syncytia up to 10 dpi.
The results conclude that CaMV.2x35S+omega promoter is induced in younger syncytia and is downregulated in older syncytia with the passage of time but can be used for expression of transgenes in syncytia. Moreover, DsRed could be used as better reporter for evaluation of gene expression in syncytia compared with GUS.
Acknowledgements
The authors are thankful to Dr. Holger Bohlmann, Associate Professor at Division of Plant Protection, Department of Crop Sciences, BOKU University, Vienna, Austria for valuable comments on the article. The authors also extend their acknowledgments to Higher Education Commission (HEC) of Pakistan for supporting the study.
References
-
Abad P, Gouzy J, Aury JM, Castagnone-Sereno P, Danchin EGJ, et al. Genome sequence of the metazoan plant-parasitic nematode Meloidogyne incognita. Nature Biotechnology, (2008); 26(8): 909-915.
-
Jones MGK. Host cell responses to endoparasitic nematode attack: structure and function of giant cells and syncytia. Annals of Applied Biology (1981); 97(3): 353–372.
-
Golinowski W, Grundler FMW, Sobczak M. Changes in the structure of Arabidopsis thaliana during female development of the plant-parasitic nematode Heterodera schachtii. Protoplasma, (1996); 194(1-2): 103-116.
-
Endo B. Ultrastructure of initial responses of susceptible and resistant soybean roots to infection by Heterodera glycines. Revue de Nematologie (1991); 14(1): 73–94.
-
Grundler FMW, Sobczak M, Lange S. Defence responses of Arabidopsis thaliana during invasion and feeding site induction by the plant-parasitic nematode Heterodera glycines. Physiological and Molecular Plant Pathology, (1997); 50(6): 419-429.
-
de Almeida Engler J, Favery B, Engler G, Abad P. Loss of susceptibility as an alternative for nematode resistance. Current Opinion in Biotechnology, (2005); 16(2): 112-117.
-
Siddique S, Sobczak M, Tenhaken R, Grundler FM, Bohlmann H. Cell Wall Ingrowths in Nematode Induced Syncytia Require UGD2 and UGD3. Plos One, (2012); 7(7): e41515.
-
Hussey RS, Grundler FM. Nematode parasitism of plants: The Physiology and Biochemistry of Free-living and Plant-parasitic nematodes. (1998), pp. 213-243. CAB International.
-
Williamson VM, Hussey RS. Nematode pathogenesis and resistance in plants. Plant Cell, (1996); 8(10): 1735-1745.
-
Benfey PN, Ren L, Chua NH. The CaMV 35S enhancer contains at least two domains which can confer different developmental and tissue-specific expression patterns. EMBO Journal, (1989); 8(8): 2195-2202.
-
Odell JT, Nagy F, Chua NH. Identification of DNA sequences required for activity of the cauliflower mosaic virus 35S promoter. Nature, (1985); 313(6005): 810-812.
-
Gallie DR, Walbot V. Identification of the motifs within the tobacco mosaic virus 5'-leader responsible for enhancing translation. Nucleic Acids Research, (1992); 20(17): 4631-4638.
-
Holtorf S, Apel K, Bohlmann H. Comparison of different constitutive and inducible promoters for the overexpression of transgenes in Arabidopsis thaliana. Plant Molecular Biology, (1995); 29(4): 637-646.
-
Jefferson RA. The GUS reporter gene system. Nature, (1989); 342(6251): 837-838.
-
Ow DW, JR DEW, Helinski DR, Howell SH, Wood KV, et al. Transient and stable expression of the firefly luciferase gene in plant cells and transgenic plants. Science, (1986); 234(4778): 856-859.
-
Chalfie M, Tu Y, Euskirchen G, Ward WW, Prasher DC. Green fluorescent protein as a marker for gene expression. Science, (1994); 263(5148): 802-805.
-
Matz MV, Fradkov AF, Labas YA, Savitsky AP, Zaraisky AG, et al. Fluorescent proteins from nonbioluminescent Anthozoa species. Nature Biotechnology, (1999); 17(10): 969-973.
-
Baird GS, Zacharias DA, Tsien RY. Biochemistry, mutagenesis, and oligomerization of DsRed, a red fluorescent protein from coral. Proceedings of National Academy of Sciences (USA), (2000); 97(22): 11984-11989.
-
Jach G, Binot E, Frings S, Luxa K, Schell J. Use of red fluorescent protein from Discosoma sp. (dsRED) as a reporter for plant gene expression. Plant Journal, (2001); 28(4): 483-491.
-
Ali MA, Shah KH, Bohlmann H. pMAA-Red: a new pPZP-derived vector for fast visual screening of transgenic Arabidopsis plants at the seed stage. BMC Biotechnology, (2012); 12(1): 37.
-
Urwin PE, Lilley CJ, McPherson MJ, Atkinson HJ. Resistance to both cyst and root-knot nematodes conferred by transgenic Arabidopsis expressing a modified plant cystatin. Plant Journal, (1997); 12(2): 455-461.
-
Liu B, Hibbard JK, Urwin PE, Atkinson HJ. The production of synthetic chemodisruptive peptides in planta disrupts the establishment of cyst nematodes. Plant Biotechnology Journal, (2005); 3(5): 487-496.
-
Goddijn OJ, Lindsey K, van der Lee FM, Klap JC, Sijmons PC. Differential gene expression in nematode-induced feeding structures of transgenic plants harbouring promoter-gusA fusion constructs. Plant Journal, (1993); 4(5): 863-873.
-
Van Poucke K, Karimi M, Gheysen G. Analysis of nematode-responsive promoters in sugar beet hairy roots. Meded Rijksuniv Gent Fak Landbouwkd Toegep Biol Wet, (2001); 66(2b): 591-598.
-
Urwin PE, Moller SG, Lilley CJ, McPherson MJ, Atkinson HJ. Continual green-fluorescent protein monitoring of cauliflower mosaic virus 35S promoter activity in nematode-induced feeding cells in Arabidopsis thaliana. Molecular Plant-Microbe Interactions, (1997); 10(3): 394-400.
-
Goverse A, Biesheuvel J, Wijers GJ, Gommers FJ, Bakker J, et al. In planta monitoring of the activity of two constitutive promoters, CaMV 35S and TR2 ', in developing feeding cells induced by Globodera rostochiensis using green fluorescent protein in combination with confocal laser scanning microscopy. Physiological and Molecular Plant Pathology, (1998); 52(4): 275-284.
-
Fuller VL, Lilley CJ, Urwin PE. Nematode resistance. New Phytologist, (2008); 180(1): 27-44.
-
Lilley CJ, Wang D, Atkinson HJ, Urwin PE. Effective delivery of a nematode-repellent peptide using a root-cap-specific promoter. Plant Biotechnology Journal, (2011); 9(2): 151-161.
-
Szakasits D, Siddique S, Bohlmann H. An improved pPZP vector for Agrobacterium-mediated plant transformation. Plant Molecular Biology Reporter, (2007); 25(3-4): 115-120.
-
Logemann E, Birkenbihl RP, Ulker B, Somssich IE. An improved method for preparing Agrobacterium cells that simplifies the Arabidopsis transformation protocol. Plant Methods, (2006); 2.
-
Sijmons PC, Grundler FMW, Vonmende N, Burrows PR, Wyss U. Arabidopsis-Thaliana as a New Model Host for Plant-Parasitic Nematodes. Plant Journal, (1991); 1(2): 245-254.
-
Epple P, Apel K, Bohlmann H. Overexpression of an endogenous thionin enhances resistance of Arabidopsis against Fusarium oxysporum. Plant Cell, (1997); 9(4): 509-520.
-
Ali MA, Abbas A, Kreil DP, Bohlmann H. Overexpression of the transcription factor RAP2.6 leads to enhanced callose deposition in syncytia and enhanced resistance against the beet cyst nematode Heterodera schachtii in Arabidopsis roots. BMC Plant Biology, (2013); 1347.
-
Ali MA, Wieczorek K, Kreil DP, Bohlmann H. The beet cyst nematode Heterodera schachtii modulates the expression of WRKY transcription factors in syncytia to favour its development in Arabidopsis roots. Plos One, (2014); 9(7): e102360.
-
Bakhetia M, Charlton WL, Urwin PE, McPherson MJ, Atkinson HJ. RNA interference and plant parasitic nematodes. Trends in Plant Science, (2005); 10(8): 362-367.
-
Ali MA, Abbas A, Azeem F, Javed N, Bohlmann H. Plant-nematode Interactions: From Genomics to Metabolomics. International Journal of Agriculture and Biology, (2015); 17(06): 1071-1082.

