

Review Article
Thyroid hormone dependent gene expression
Ghulam Zahara Jahangir1*, Faiza Saleem2, Shagufta Naz2, Neelma Munir2, Rukhama Haq2, Aleena Sumrin1
Adv. life sci., vol. 3, no. 4, pp. 119-124, August 2016
*- Corresponding Author: Ghulam Zahara Jahangir (Email: zahra_jahangir@yahoo.com)
Authors' Affiliations
2- Department of Biotechnology & Microbiology, Lahore College for Women University, Jail Road, Lahore – Pakistan
Abstract![]()
Introduction
Methods
Discussion
References
Abstract
The presented work is destined to review the advances that had been made to study the role of thyroid hormone and thyroid hormone nuclear receptors in regulating the gene expression. Triiodothyronine (T3) and tetraiodothyronine (thyroxine or T4) are most important thyroid hormones. The thyroid hormones bind to their specific nuclear hormone receptors, as ligand, and play important role in gene expression and transcriptional gene regulation in human and higher animals. Thyroid hormone receptors form heterodimers by making combination with retinoid X receptors. The capability of heterodimerization of thyroid hormones generates novel complexes which allow altered specificity and higher affinity for DNA-receptor binding. Thyroid hormone receptors work as ligand activated transcription factor and play with transcriptional gene expression process. The consensus structural features of thyroid hormone receptors are N-terminal regulatory domain that contains activation function, the domain for strong gene expression and the domain for binding to DNA. The structures for individual domains have been extensively and reviewed through several latest and successful techniques.
Keywords: Thyroid, Thyroid receptors, Gene regulation, Hormone response elements, Co-activator, Co-repressor
Introduction
Cells are capable to alter the expression of some of their genetic entities in accordance with their particular spatiotemporal context. In higher organisms, cells must respond to the external stimuli and signals for proper coordination, development, and the functioning of all organ systems, tissues, and whole organism (human and higher organisms) [1,2]. This ability is vital for their adaptation and survival in the fluctuating surroundings. Signal transduction through nuclear receptors is simpler because the cells are able to recognize the signal and show response directly by to interacting the nuclear genome [1].
Thyroid hormones are iodine containing peptides and are produced in the thyroid gland. Two very important thyroid hormones are triiodothyronine (T3) and tetraiodothyronine (thyroxine or T4). Thyroxine is reported to be the first hormone that was isolated and studied for structure determination, whereas, the T3 was identified in the circulation, many decades later [3]. On molar basis, T3 is 10-fold more active than T4 and plays dominant role in maintaining balance in thyroid levels. These hormones are vital for life and affect a broader range of mechanisms involved in development, differentiation, growth, morphogenesis, metabolism, and reproduction of humans and other higher organisms [4,5]. The thyroid stimulating hormone (
At cellular level thyroid hormones perform their function as ligand and bind to the nuclear hormone receptors; which in turn behave as ligand-inducible transcription factors [4]. Thyroid hormone receptors bind to small ligand molecules, the thyroid hormone, and regulate transcription by binding to the sequences with high specificity for thyroid hormone [4,7,8]. These thyroid hormone-responsive sequences are observed in the promoter regions of target genes [7]. These molecules control important functions related to different diseases, for instance, diabetes, osteoporosis and cancer; and can be modified by drug design. Owing to this property nuclear hormone receptors are promising pharmacological targets [8].
Methods
Search Strategy and Selection Criteria
In this study the domain structure of thyroid nuclear receptors and the functional mechanism of thyroid hormone receptors have been reviewed along with the discussion upon DNA hormone response elements to which they bind generally as homo- or heterodimers. The description has been limited around DNA binding-dependent regulation by thyroid ligand. The mechanism of transcriptional repression has also been described with more detail. For the completion of this work the published research data has been studied from various sources.
Discussion
Thyroid Hormone Receptors
How thyroid hormone plays role in gene expression. The answer can only be found when the working three-dimensional structure of thyroid hormone receptors is known. The thyroid hormone receptors are grouped, in a large subfamily of the superfamily designated for the nuclear receptors. The receptors for small lipophilic hormones like retinoic acid, different orphan and peroxisome proliferator-activated receptors and vitamin D receptors are grouped in this subfamily among the thyroid receptors [4,7]. Thyroid hormone receptors in combination with retinoid X receptors form heterodimers [7].
Due to heterodimerization novel complexes are produced by thyroid nuclear receptors that can bind DNA with altered specificity and higher affinity [1,9]. Whereas, in the monomeric form they show no considerable contributions in the transcriptional properties and only a little is known [10]. Molecular cloning and structural and functional analyses revealed that all the members of the superfamily share common functional domain structure [11]. Nuclear hormone receptors work as ligand-activated transcription factors and provide clear and direct connection between the transcriptional responses and the signaling molecules that govern these processes [8].
Overview of Structure
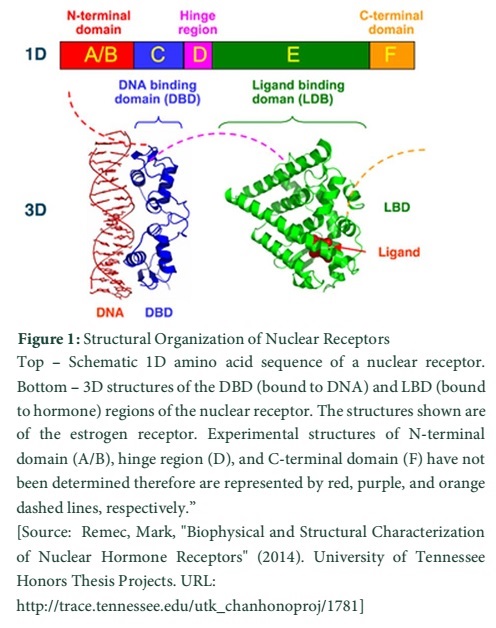
The N-terminal regulatory domain also known as (A-B) contains AF-1 or Activation Function 1. It is important for transactivation of transcription and works without depending on the ligand [11,12]. The transcriptional activation of AF-1 is weak and functions in cooperation with AF-2 in the E-domain for strong gene expression. Various nuclear receptors possess variable sequence for A-B domain. The C domain or DNA-binding domain (DBD) is crucial for recognition of specific DNA sequences and for protein interactions. It is well conserved and its zinc fingers, called hormone response elements (HRE), are responsible for DNA binding specificity [11,13]. The Hinge region, (D) domain, is reported for connecting DNA-binding domain to the Ligand binding domain (LBD). It is recognized as flexible and for influencing intracellular trafficking [11,13]. LB domain or (E) shows strong conservation in structure and a bit less in sequence for various nuclear receptors. The LB domain bears AF-2 and its action depends upon the presence of bound ligand. In different nuclear receptors the sequence of F or C-terminal domain is highly variable [12]. The N-terminal, DNA-binding, and ligand binding domains (A/B, C, and E respectively) possess independent folding and stable structures while a disordered and flexible conformational trend has been noted in hinge region and the C-terminal domains (D and F respectively) [14].
Crystallographic Studies
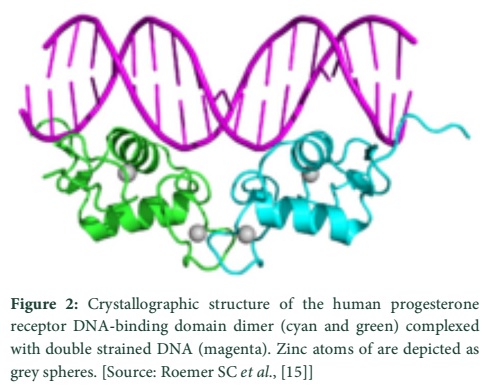
It is depicted from the results of crystallographic studies carried upon thyroid receptors that ligand plays a structural role in the LB domain of thyroid receptor and influences the folding of its hydrophobic core. Further structural analysis directed to the genetic and biochemical studies. These studies have revealed that sites are essential to be present on the thyroid receptor ligand binding domain for the binding of co-activator proteins and for dimerization. When the patients possessing thyroid hormone resistance, syndrome of generalized resistance, were studied; it revealed that there was a mutation on thyroid receptor LB domain in a site very close to the thyroid hormone binding site [15].
Two subtypes of thyroid hormone receptor, TRα and TRβ, are involved in most of the physiological effects of thyroid hormones. Valadares et al., determined the affinity and selectivity of TRα and TRβ subtype for TR ligands using ligand and structure-guided approach which was pharmacophore-based [16]. Using CoMFA method, the comparative molecular field analysis, they obtained the three-dimensional quantitative structure-selectivity and activity relationships (3D-QSSR) and (3D-QSAR) respectively, which were statistically reliable. With the help of 3D-QSSR models they determined halogen-bonds’ function in the selectivity of receptor for thyroid hormone [16,17].
Functional Mechanism of Thyroid Hormone Receptors
Triiodothyronine (T3) binds to the nuclear receptors specified for thyroid hormone and regulates gene expression. Nuclear receptor co-activators are associated with ligand bound receptors and enhance gene transcription [18]. To promote transcription histones are acetylated with the help of enzymes recruited by co-activators and this course is sensitive to the metabolic state of cell [19,20]. Other wider level studies have established the conclusion that direct interaction between DNA and nuclear receptors regulate the gene expression. However, these nuclear receptors may help opening chromatin in a way distinct from direct DNA binding [19]. For the stimulation of T3-regulated genes’ transcription, the TR complex employs co-activators (like some members of SRC family of proteins) [18]. Research studies regarding the T3 action reveal the importance of SRC1 for the increased TSH value in response to hypothyroidism.
A heterodimer complex is formed between thyroid receptor and retinoid X receptor (RXR), this complex is capable to alter the gene expression by stimulating or repressing the gene transcription after binding onto a specific DNA sequences in the target genes. This DNA sequence is referred as thyroid response element (TRE) [6]. These hormone response elements are studied to be positioned in the regulatory sequences of target gene towards 5’ sides. In some situations these are present numerous kilobases upstream from the site of start of transcription, in the enhancer regions of target genes. Several studies revealed that a core recognition motif is made up of 6 bp sequence. Two consensus idealized recognition motif sequences have been identified and AGG/TTCA has been described for thyroid hormone receptors and for other non-steroidal receptors of nuclear receptors superfamily [4,21]; but a significant deviation from idealized consensus has been observed in the naturally occurring HREs. Most receptors (homo- or heterodimers) bind to the two core motifs elements (hexameric). In dimeric elements, the half-sites are described to be as direct repeats (DRs), palindromes (Pal), or inverted palindromes (IPs). The receptors may bind to these hormone response elements with receptor’s differential selectivity and specificity which is determined by spacing between the motifs and their arrangement [4]. Most of the studies have been focused on the transcriptional activation of target genes by thyroid ligand-nuclear receptor binding to positive elements but gene expression may also be repressed by nuclear receptors in the same manner. Thyroid hormones regulate gene expression by palindromic elements, AGGTCATGACCT along with retinoic acid [22]. Resultantly the thyroid and retinoic acid ligands can control gene expression through overlapping networks. Furthermore, the attachment of homodimers on to the DNA is repressed by thyroid hormone whereas it does not inhibits the binding of the heterodimers to latter; this way the creation of heterodimeric complexes is enhanced at the HREs [23].
The co-repressor binds to the nuclear receptor in unliganded state and endorses gene repression. When unliganded, TR complex associates with co-repressors like nuclear receptor corepressor (NcoR) or silencing mediator for retinoid and thyroid hormone receptor (SMRT) and causes decreased transcription of T3-regulated genes and antagonize the action of other nuclear receptor complexes [24,25]. It is achieved with the assistance of histone deacetylases (HDACs), recruited by Co-repressors, to the promoters of target genes for enhanced repression by unliganded TR [26]. Sometimes thyroid ligand mediates negative regulation by binding to “negative TREs” in the genes of TSH and TRH. Thyroid negative regulation is also controlled by some other negative elements which bind to thyroid receptor homodimer when thyroid is absent, and to retinoid X receptor- thyroid receptor heterodimer (RXR/TR) when 3, 3′,5-triiodothyronine (T3) is present. The negative elements, generally, are located very close to the transcription initiation sites and sometimes downstream to the TATA box [27-29]. These are also observed to be present at unusual site, at the 3′-untranslated region [30]. These sequence show negative regulation only when these are present downstream of the transcription initiation site. To the date, the properties of the negative elements are not fully understood; but the location of these elements may a play an important role [4]. In addition to thyroid-dependent gene activation, TR represses basal transcription when thyroid is absent. Binding of thyroid to the TR reliefs the transcriptional silencing and promotes gene activation. Presences of active silencing domains in TR were first demonstrated by Bahniahmad et al., [31].
Reference
- Beato M (1993) Gene regulation by steroid hormones. Gene Expression: Springer. pp. 43-75.
- Wolff E, Wolff J. The mechanism of action of the thyroid hormones. The Thyroid Gland, (1964); 1237-282.
- Samuels H, Forman B, Horowitz Z, Ye Z. Regulation of gene expression by thyroid hormone. Journal of Clinical Investigation, (1988); 81(4): 957.
- Aranda A, Pascual A. Nuclear hormone receptors and gene expression. Physiological Reviews, (2001); 81(3): 1269-1304.
- Brent GA. The molecular basis of thyroid hormone action. New England Journal of Medicine, (1994); 331(13): 847-853.
- Mullur R, Liu Y-Y, Brent GA. Thyroid hormone regulation of metabolism. Physiological Reviews, (2014); 94(2): 355-382.
- Zhang J, Lazar MA. The mechanism of action of thyroid hormones. Annual Review of Physiology, (2000); 62(1): 439-466.
- Robinson-Rechavi M, Garcia HE, Laudet V. The nuclear receptor superfamily. Journal of Cell Science, (2003); 116(4): 585-586.
- Glass CK. Differential Recognition of Target Genes by Nuclear Receptor Monomers, Dimers, and Heterodimers*. Endocrine Reviews, (1994); 15(3): 391-407.
- Forman BM, Umesono K, Chen J, Evans RM. Unique response pathways are established by allosteric interactions among nuclear hormone receptors. Cell, (1995); 81(4): 541-550.
- Kumar R, Thompson EB. The structure of the nuclear hormone receptors. Steroids, (1999); 64(5): 310-319.
- Wärnmark A, Treuter E, Wright AP, Gustafsson J-A. Activation functions 1 and 2 of nuclear receptors: molecular strategies for transcriptional activation. Molecular endocrinology, (2003); 17(10): 1901-1909.
- Klinge CM. Estrogen receptor interaction with co-activators and co-repressors☆. Steroids, (2000); 65(5): 227-251.
- Weatherman RV, Fletterick RJ, Scanlan TS. Nuclear-receptor ligands and ligand-binding domains. Annual review of biochemistry, (1999); 68(1): 559-581.
- Roemer SC, Donham DC, Sherman L, Pon VH, Edwards DP, et al. Structure of the progesterone receptor-deoxyribonucleic acid complex: novel interactions required for binding to half-site response elements. Molecular Endocrinology, (2006); 20(12): 3042-3052.
- Valadares NF, Salum LB, Polikarpov I, Andricopulo AD, Garratt RC. Role of halogen bonds in thyroid hormone receptor selectivity: pharmacophore-based 3D-QSSR studies. Journal of Chemical Information and Modeling, (2009); 49(11): 2606-2616.
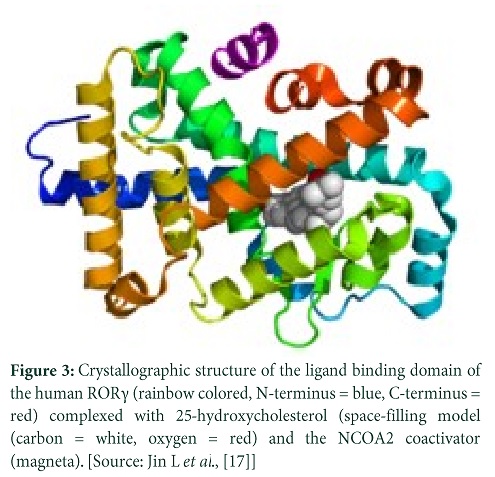
- Jin L, Martynowski D, Zheng S, Wada T, Xie W, et al. Structural basis for hydroxycholesterols as natural ligands of orphan nuclear receptor RORγ. Molecular Endocrinology, (2010); 24(5): 923-929.
- Dasgupta S, Lonard DM, O’Malley BW. Receptor Coactivators: Master Regulators of Human Health and Disease. Annual Review of Medicine, (2014); 65: 279.
- Everett LJ, Lazar MA. Cell‐specific integration of nuclear receptor function at the genome. Wiley Interdisciplinary Reviews: Systems Biology and Medicine, (2013); 5(5): 615-629.
- Lu C, Thompson CB. Metabolic regulation of epigenetics. Cell Metabolism, (2012); 16(1): 9-17.
- Beato M, Herrlich P, Schütz G. Steroid hormone receptors: many actors in search of a plot. Cell, (1995); 83(6): 851-857.
- Umesono K, Giguere V, Glass CK, Rosenfeld MG, Evans RM. Retinoic acid and thyroid hormone induce gene expression through a common responsive element. Nature, (1988); 336(6196): 262-5.
- Ribeiro R, Kushner PJ, Apriletti JW, West BL, Baxter JD. Thyroid hormone alters in vitro DNA binding of monomers and dimers of thyroid hormone receptors. Molecular Endocrinology, (1992); 6(7): 1142-1152.
- Araki O, Ying H, Zhu X, Willingham M, Cheng S. Distinct dysregulation of lipid metabolism by unliganded thyroid hormone receptor isoforms. Molecular Endocrinology, (2009); 23(3): 308-315.
- Mottis A, Mouchiroud L, Auwerx J. Emerging roles of the corepressors NCoR1 and SMRT in homeostasis. Genes & development, (2013); 27(8): 819-835.
- Hsia EY, Goodson ML, Zou JX, Privalsky ML, Chen H-W. Nuclear receptor coregulators as a new paradigm for therapeutic targeting. Advanced Drug Delivery Reviews, (2010); 62(13): 1227-1237.
- Belandia B, Latasa MJ, Villa A, Pascual A. Thyroid hormone negatively regulates the transcriptional activity of the β-amyloid precursor protein gene. Journal of Biological Chemistry, (1998); 273(46): 30366-30371.
- Pérez-Juste G, Garcı́a-Silva S, Aranda A. An element in the region responsible for premature termination of transcription mediates repression of c-myc gene expression by thyroid hormone in neuroblastoma cells. Journal of Biological Chemistry, (2000); 275(2): 1307-1314.
- Saatcioglu F, Deng T, Karin M. A novel cis element mediating ligand-independent activation by c-ErbA: implications for hormonal regulation. Cell, (1993); 75(6): 1095-1105.
- Bigler J, Eisenman RN. Novel location and function of a thyroid hormone response element. The EMBO journal, (1995); 14(22): 5710.
- Baniahmad A, Köhne A, Renkawitz R. A transferable silencing domain is present in the thyroid hormone receptor, in the v-erbA oncogene product and in the retinoic acid receptor. The EMBO Journal, (1992); 11(3): 1015.

