

Full Length Research Article
Effect of Salinity (NaCl) stress on germination and early seedling growth of three medicinal plant species
Fatemeh Bina1 and Abdolamir Bostani2,*
Adv. life sci., vol. 4, no. 3, pp. 77-83, May 2017
*– Corresponding Author: Dr. Abdolamir Bostani (Email: bostani@shahed.ac.ir)
Authors' Affiliations
2- Department of Soil Science, Faculty of Agriculture, Shahed University, Tehran – Iran
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Salinity stress negatively affects the growth and yield of plants. Due to the increasing demand for products derived from medicinal plants and with regard to the growing problems caused by salinity of arable lands, the use of salt-tolerant species can be a strategic approach to cope with this problem. The aim of the present study was to evaluate the effect of salinity stress on germination and seedling growth of three medicinal plant species.
Methods: Seeds of three species of medicinal herbs including isabgol (Plantago ovata L.), zucchini (Cucurbita pepo L.) and clove (Caryophyllus aromaticus L.) were exposed to different concentrations (0, 10, 20, 30 and 40 dS·m-1) of sodium chloride (NaCl).
Results: It was observed that, except for zucchini, germination was inhibited at higher salinity regimes. NaCl treatment caused a serious decrease in the early seedling growth by means of reduced root and shoot length at higher salinity levels. Regression analysis of studied indices over salinity levels revealed that the highest and lowest slopes of regression lines belonged to isabgol and zucchini, respectively indicating high tolerance of zucchini and also, high sensitivity of isabgol to the imposed salinity levels.
Conclusions: Based on the findings, zucchini was able to germinate more than 90% at 40 dS·m-1 of NaCl. Therefore, zucchini is suggested to be cultivated in farmlands which are relatively faced with the problem of soil salinity while, it is recommend that isabgol and clove not to be planted in such soils.
Key words: Salt Stress, Germination analysis, Clove, Isabgol, Regression analysis, Zucchini
Introduction
All cultures, from ancient time to the present, have used plants as a source of medicine. According to the World Health Organization (WHO), most of the world’s population depends upon plants as an important element in primary health care system [1, 2].
Natural products are a source for novel antibiotics. In the area of cancer, from around the 1940s to date, 47% of the antibiotics used have been actually either natural products or directly derived therefrom. In other areas, the influence of natural product structures is quite marked, with the anti-infective area being dependent on natural products and their structures [3].
Isabgol (Plantago ovata), has been used as a dietary fiber supplement, bulk-forming laxative. During the last decade, dietary supplementation with Isabgol has been shown to lower blood cholesterol level (especially LDL cholesterol) and to maintain blood glucose homeostasis, which together is the most effective preventive measures against diabetes and cardiovascular disease [4].
Zucchini (Cucurbita pepo), is a widely used food and an important component of folk medicine around the world. Extracts and metabolites of this plant, particularly those from seeds and fruits possess useful pharmacological activities. C. pepo, is mainly known for its improvement in prostatic hyperplasia, urinary dysfunction and cytotoxic properties, also it has been used extensively as a hypoglycemic agent. Many pharmacological studies have demonstrated hepatoprotection, inhibition of benign prostatic hyperplasia, antioxidant, anticancer, antimicrobial, anti-inflammatory, antidiabetic, and antiulcer activities supporting its traditional uses [5].
Clove (Caryophyllus aromaticus L.), has been widely used in the flavoring industries, fragrance and cosmetics. Clove extract (CE) is isolated from the buds of clove and is not harmful when consumed in food products. Radha Krishnan et al. [6] reported that the addition of CE could effectively inhibit microbial growth, decrease lipid oxidation, maintain or improve sensory attributes and extend the shelf-life of raw chicken meat during refrigerated storage. The natural extracts from clove applied to raw pork could help to reduce the total bacterial count and retard lipid oxidation during storage at ambient temperature for up to 9 days. Therefore, CE exhibited the most effective antibacterial and the highest antioxidant activity.
Abiotic stresses, such as salinity, always limits the growth, distribution and production of plants. According to a recent estimate, 1128 million ha of global land is affected by salinity and sodicity [7, 8]. Due to high evapotranspiration and low rainfall, the majority areas of Iran has been classified as arid and semi-arid. According to statistics released by the Ministry of Agriculture, total area of saline soils of Iran is estimated to be about 44 million hectares which is about 30% of plains and more than 50% of the country’s irrigated lands. Considering that salinity is a stable stress, plants will be permanently exposed to it during their growth period. Seed germination “as the most important stage of plant growth” is also affected by salinity. Increased Na+ uptake in plants under salinity disturbs those metabolic processes that require low Na+ and high K+, Ca2+ or both for normal functioning [8].
The growing use of medicinal plants has led to the increase of their cultivation area. Thus, study of their tolerance to adverse environmental conditions, such as salinity of soil and/or irrigation water is necessary because it leads to a greater level use of lands that are somewhat faced with problems caused by salinity. Studies have shown that salinity has a negative effect on germination characteristics of some herbs [9]. However, investigations are still needed to understand the germination of Isabgol, Zucchini and Clove under saline stress conditions.
The purpose of this experiment was to evaluate the effects of salinity stress on germination indices of three medicinal plants; Isabgol, Zucchini and Clove.
Methods
The experiment was conducted in 2015 at Shahed University in the form of factorial based on completely randomized design with three replications. Studied factors were: A) three species of medicinal plants including: Isabgol (Plantago ovata), Zucchini (Cucurbita pepo) and Clove (Caryophyllus aromaticus L.) and B) applying salinity stress using NaCl at five concentrations of 0 (Control), 10, 20, 30 and 40 dS·m-1. Seeds needed for the experiments were obtained from Medicinal Plant Research Center of Shahed University, Tehran, Iran. First, the seeds were disinfected with sodium hypochlorite 10% for 3 minutes. Seeds were then rinsed with distilled water. Afterward, the number of 50 seeds from each plant species were sown on a sterilized Whatman paper in each petri dish and were exposed to five levels of salinity stress as explained above. Petri dishes were stored at room temperature (20-25°C). From the second day and at a specified time, the number of germinated seeds was daily counted for 14 days. According to International Seed Testing Association [10], seeds with a root length greater than 2mm were considered as germinated. After 14 days, shoot length (SL) and root length (RL) were measured.
Calculated germination indices
Germination percentage was calculated according to Ranal et al. [11]. Also, SVI was calculated by the formula proposed by Abdul- Baki and Anderson [12] as follows:
SVIij = SMLij x GP%ij
Where, SVI is the seed vigor index, SML is the seedling mean length in cm (root + shoot), and GP is the germination percentage for plant species i at salinity level of j. Mean comparisons were carried out with Duncan method at the significance level of 0.05 by using the SAS statistical software (SAS 2002). Regression illustrations have been drawn using Microsoft Excel 2013.
Results
Germination percentage
Table 1 shows the mean germination percentage of the three studied herb species exposed to salinity stress treatment. According to the results, zucchini showed a high and stable value of GP and as a result, a very high resistance to salinity stress at all treatment levels, as compared with clove and isabgol. No remarkable decrease was found for zucchini’s GP even at 40 dS·m-1 of salinity level. On the other hand, a severe decrease was observed for isabgol’s GP, concurrent with increase in the intensity of the salinity stress. The germination percent of isabgol at control and 40 dS·m-1 of salinity were 99.23 and 1.47%, respectively showing a significant decrease. As shown in Table 1, response of clove to salinity stress was at intermediate level among the two other studied plants. The germination percent of clove at control, 10, 20, 30 and 40 dS·m-1 of salinity were 100, 75, 55.67, 47.67 and 34.67%, respectively. Results of the regression analysis for GP as dependent variable, and salinity stress levels as independent variable showed that the slope of the regression line for clove, isabgol and zucchini were -1.58, -2.35 and -0.13, respectively (Figure 1). Therefore, isabgol and zucchini had the highest and lowest slopes, respectively suggesting the high tolerance of zucchini and at the same time, the high sensitivity of isabgol to the imposed salinity levels (Figure 1).
Seed vigor index
The averages of seed vigor index of the three studied herb species under the impact of salinity stress have been presented in Table 1. According to this Table, increase of stress severity led to the dramatic reduction of SVI for clove and isabgol while, the pattern of reduction in SVI for zucchini was found to be different. Seed vigor index for zucchini at concentrations of control, 10, 20, 30 and 40 dS·m-1 were 11.12, 3.98, 3.52, 2.22 and 1.63, respectively (Table 1).
In fact, similar to the results obtained for GP, here again, the SVI was reduced with a more lenient slop than isabgol and clove. Results of the regression analysis for SVI, where salinity stress level was considered as independent variable, showed that the slope of the regression line for clove, isabgol and zucchini were -0.61, -0.64 and -0.21, respectively (Figure 1). Therefore, here again, isabgol and zucchini had the highest and lowest slopes, respectively suggesting the high tolerance of zucchini and at the same time, the high sensitivity of isabgol to the imposed salinity stress levels (Figure 1).
Growing indices
Growing indices including root and shoot length were also measured and reported in Table 1. According to this table, for all the three studied medicinal plants, the pattern of variation due to salinity observed for root and shoot length showed a downward trend. For example, the average of root length recorded for isabgol at 0 and 40 dS·m-1 of salinity was 92.4 and 0.33 mm, respectively. Similarly, the root length was 52 and 4.67 mm for zucchini and 99.73 and 17.33 mm for clove at 0 and 40 dS·m-1 of salinity, respectively. Moreover, the estimated regression coefficients for root length where salinity stress levels was considered as predictor variable, showed that the slopes of the regression lines for clove, isabgol and zucchini were -1.78, -2.03 and -1.08, respectively. Therefore, similar to the results obtained for GP, the highest and lowest regression slopes belonged to zucchini and isabgol, respectively indicating high tolerance of zucchini and at the same time high sensitivity of isabgol to the imposed salinity stress levels (Figure 2).
In this study, the response of shoot length to the imposed salinity stress levels was very similar to that of observed for root length (Table 1). Here again, a decreasing pattern was observed under salinity stress. The slope of regression lines estimated for shoot length, where salinity stress level was considered as predictor variable were -3.95, -4.71 and -0.97 for clove, isabgol and zucchini, respectively. According to the results, except for zucchini, the regression slopes calculated for shoot length was greater than those slopes that were estimated for root length for two other medicinal plants under study. Thus, stem length was more affected by salinity as compared to root length for clove and isabgol. However, zucchini showed a reverse trend with reference to greater slope of the regression line, calculated for root length (-1.08) than shoot length (-0.97).
Tables & Figures
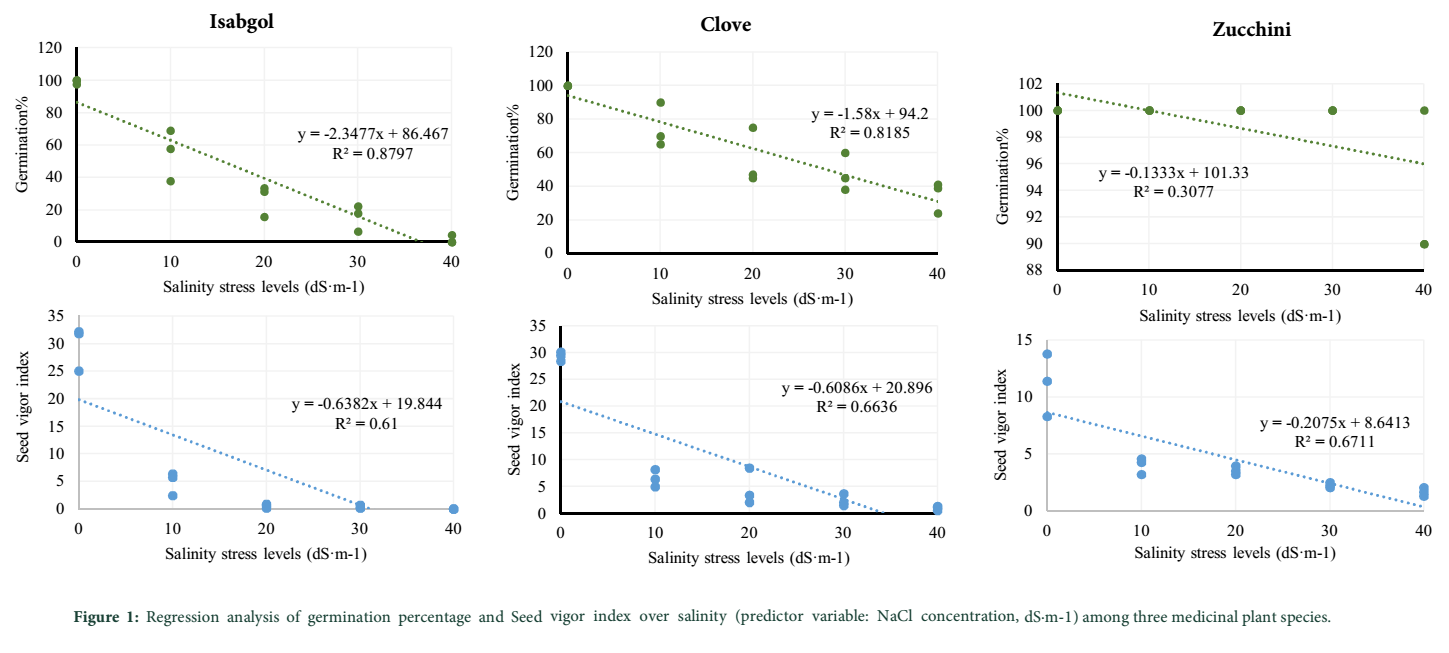
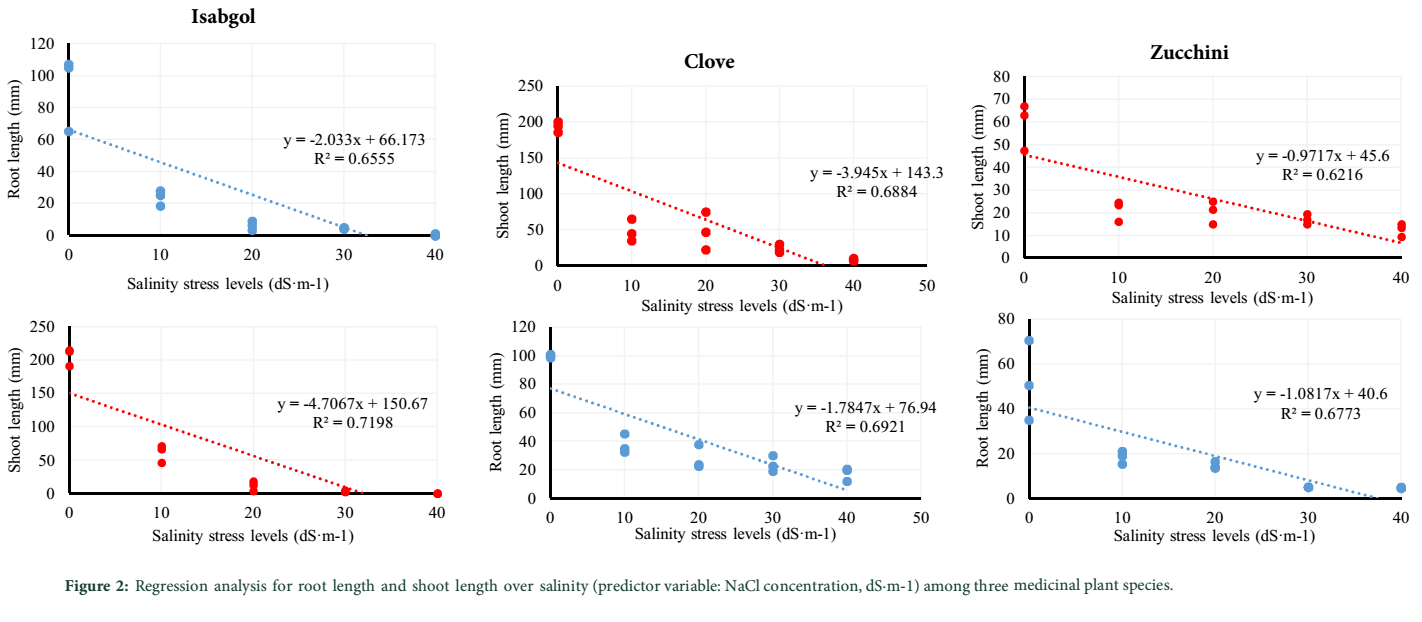

Discussion
Among one of the main environmental factors, salinity adversely affects the growth and yield of plants. Due to the increasing demand for medicinal plants and with regard to the cultivation problem due to salinity of arable lands, the use of salt-tolerant species can be a strategic approach to deal with this problem. Resistance at germination stage is of the most important aspects of salt tolerance. Hence, in this study, salt tolerance of 3 medicinal species was evaluated in the presence of five different concentrations of NaCl solution at seed germination stage.
Recent studies on the heritability of salt tolerance have shown that salt tolerance is mainly controlled by genetic effects and thus the majority of the phenotypic variation observed for this trait is due to genetic effects. Also, narrow-sense heritability of salt tolerance indicated that the majority of the phenotypic variation is mainly due to additive gene effects [13]. Therefore, study of different plant species in order to find sources of salt resistance is important because, as mentioned above, due to high heritability of salinity resistance, they can be used to breed commercial cultivars through the transferring of genes responsible for resistance. Besides, the availability of sufficient information regarding the salinity of arable lands and resistant/tolerant plant species, it can help us to scientifically deal with the phenomenon of salinity of arable lands.
Results of the present study showed that the response to salinity stress at germination stage was variable among the three studied medicinal plants. As mentioned by Prado et al. [14] the sensitivity of plants to salinity depends on plant species and their developmental stage. The decrease in GP under salt stress condition may be due to the fact that seeds seemingly develop an osmotically enforced dormancy under water-stress conditions. This may be an adaptive strategy of seeds to prevent germination in stressful environments, thereby ensuring the proper establishment of seedlings [15, 16].
It has been reported that at high salinities, the adaptation of plants to saline environments relies on salt tolerance or salt-avoidance mechanisms [17]. Typical salt tolerance strategies are reflected by high germination percentages and/or germination rates under high salinities [18]. Therefore, in this study, the studied species can be divided into three categories in terms of resistance to salinity as: i) marginally tolerant (able to germinate more than 50% at 10 dS·m-1 NaCl, isabgol) ii) moderately tolerant (able to germinate more than 50% at 20 dS·m-1 NaCl, clove) and highly tolerant (able to germinate more than 50% at 40 dS·m-1 NaCl, zucchini).
In this study, root length under salinity stress was significantly decreased as compared with control treatment. This implied that salt stress remarkably inhibited root development showing inhibition in root cell proliferation. As shown by Long et al. [19], there are some mechanisms in this regard such as: (1) High concentrations of NaCl in the environment led to the decrease in water potential. Consequently, plant cells faced some difficulties to absorb external water. Although the expression of some genes involved in the proline synthesis pathway, such as Pyrroline-5-carboxylate synthase, could balance the water potential inside and outside the cells however, this change could not help cells absorb enough water to perform related substrate synthesis for cell development. Thus, root development was decreased [19]. (2) Salt stress induces reactive oxygen species (ROS), which leads to secondary oxidation stress, disturbs cellular redox homeostasis, and damages cell components and structures [20]. Under salinity stress, high amount of ROS generated in root cells might have damaged the existing active cells, which had a negative effect on cell proliferation. (3) Salinity stress negatively affects cell wall reconstruction and re-synthesis, which would interfere with roots cell expansion and division [19]. Similar to root length, stem length was also significantly reduced under salt stress. It seems that similar to root length, the osmotic differences were the main cause of such phenomenon.
In conclusion, results of the present study revealed that isabgol and zucchini had the highest and lowest tolerance to the imposed salinity levels, respectively. Based on the findings, zucchini was able to germinate more than 90% at 40 dS·m-1 of NaCl. Therefore, cultivation of zucchini is suggested for farmlands which have the problem of soil salinity. Although different many researchers have had developed diverse technique, e.g. transgenic, to improve various traits of field crops [21-28] but results of this kind of work can surely be useful as a strategy to deal with the problem of soil salinity and/or irrigation water.
References
- Tomlinson TR. Promoting the worldwide use of medicinal plants: Medicinal plants: their role in health and biodiversity. 2015; 1: pp 206. University of Pennsylvania press Philadelphia.
- Khan M. Indian medicinal plants. Indian Journal of Experimental Biology (2011); 49(2): 797-798.
- Newman DJ, Cragg GM. Natural products as sources of new drugs over the last 25 years. Journal of Natural Products, (2007); 70(3): 461-477.
- Deokar G, Kshirsagar S, Deore PA, Kakulte H. Pharmaceutical benefits of Plantago ovate (Isabgol seed): a review. Pharmaceutical and Biological Evaluations, (2016); 3(1): 32-41.
- Gutierrez RMP. Review of Cucurbita pepo (Pumpkin) its Phytochemistry and Pharmacology. Medicinal Chemistry, (2016); 6(1): 12-21.
- Radha krishnan K, Babuskin S, Azhagu Saravana Babu P, Sasikala M, Sabina K, et al. Antimicrobial and antioxidant effects of spice extracts on the shelf life extension of raw chicken meat. International Journal of Food Microbiology, (2014); 171(3): 32-40.
- Chen H, Diao J, Li Y, Chen Q, Kong B. The effectiveness of clove extracts in the inhibition of hydroxyl radical oxidation-induced structural and rheological changes in porcine myofibrillar protein. Meat Science, (2016); 111: 60-66.
- Akhtar SS, Andersen MN, Naveed M, Zahir ZA, Liu F. Interactive effect of biochar and plant growth-promoting bacterial endophytes on ameliorating salinity stress in maize. Functional Plant Biology, (2015); 42(8): 770-781.
- Seghatoleslami M. Effect of Salt Stress on Germination of Satureja hortensis L., Cichorium intybus L. and Cynara scolymus L. Journal of Iranian Field Crop Research, (2011); 8(5): 818-823.
- ISTA (2016) ISTA Certificates. International rules for seed testing. Zürichstr. 50, CH-8303 Bassersdorf, Switzerland: The International Seed Testing Association (ISTA). pp. i-1-14
- Ranal MA, Santana DGd, Ferreira WR, Mendes-Rodrigues C. Calculating germination measurements and organizing spreadsheets. Brazilian Journal of Botany, (2009); 32(4): 849-855.
- Abdul- Baki AA, Anderson JD. Vigour determination in soybean by multiple criteria. Crop Science, (1973); 13(6): 630-633.
- Koch MJ, Meyer WA, Bonos SA. Inheritance of Salinity Tolerance in Perennial Ryegrass. Crop Science, (2015); 55(4): 1834-1842.
- Prado FE, Boero C, Gallardo M, Gonzalez JA. Effect of NaCl on germination, growth, and soluble sugar content in Chenopodium quinoa Willd. seeds. Botanical Bulletin of Academia Sinica, (2000); 4: 27-34.
- Gill PK, Sharma AD, Singh P, Bhullar SS. Changes in germination, growth and soluble sugar contents of Sorghum bicolor (L.) Moench seeds under various abiotic stresses. Plant Growth Regulation, (2003); 40(2): 157-162.
- Nasri N, Maatallah S, Kaddour R, Lachâal M. Effect of salinity on Arabidopsis thaliana seed germination and acid phosphatase activity. Archives of Biological Sciences, (2016); 68(1): 17-23.
- Nichols PGH, Craig AD, Rogers ME, Albertsen TO, Miller SM, et al. Production and persistence of annual pasture legumes at five saline sites in southern Australia. Australian Journal of Experimental Agriculture, (2008); 48(4): 518-535.
- Zhang H, Zhang G, Lü X, Zhou D, Han X. Salt tolerance during seed germination and early seedling stages of 12 halophytes. Plant and Soil, (2014); 388(1): 229-241.
- Long W, Zou X, Zhang X. Transcriptome Analysis of canola (Brassica napus) under salt stress at the germination stage. PloS one, (2015); 10(2): e0116217.
- Shalata A, Neumann PM. Exogenous ascorbic acid (vitamin C) increases resistance to salt stress and reduces lipid peroxidation. Journal of Experimental Botany, (2001); 52(364): 2207-2211.
- Yasmeen A, Kiani S, Butt A, Rao AQ, Akram F, et al. Amplicon-Based RNA Interference Targeting V2 Gene of Cotton Leaf Curl Kokhran Virus-Burewala Strain Can Provide Resistance in Transgenic Cotton Plants. Molecular Biotechnology, (2016); 58(12): 807-820.
- Ramzan M, Tabassum B, Nasir IA, Khan A, Tariq M, et al. Identification and application of biocontrol agents against Cotton leaf curl virus disease in Gossypium hirsutum under greenhouse conditions. Biotechnology & Biotechnological Equipment, (2016); 30(3): 469-478.
- Ali A, Ahmad Nasir I, Muzaffar A, Shahzad Iqbal M, Qayyum Rao A, et al. Screening of potato germplasm resistant against low temperature sweetening. Journal of Food Quality, (2016); 39(4): 301-310.
- Awan MF, Ali A, Muzaffar A, Abbas MA, Rao AQ, et al. Transgenic cotton: Harboring Broad Term Resistance against Insect and Weeds through Incorporation of CEMB Double Bt and cp4EPSPS Genes. Pakistan Journal of Agricultural Sciences, (2016); 53(3): 501-505.
- Ali A, Ahmed S, Nasir IA, Rao AQ, Ahmad S, et al. Evaluation of two cotton varieties CRSP1 and CRSP2 for genetic transformation efficiency, expression of transgenes Cry1Ac+ Cry2A, GT gene and insect mortality. Biotechnology Reports, (2016); 966-73.
- Puspito AN, Rao AQ, Hafeez MN, Iqbal MS, Bajwa KS, et al. Transformation and Evaluation of Cry1Ac+ Cry2A and GTGene in Gossypium hirsutum L. Frontiers in Plant Science, (2015); 6: 943.
- Bajwa KS, Shahid AA, Rao AQ, Bashir A, Aftab A, et al. Stable transformation and expression of GhEXPA8 fiber expansin gene to improve fiber length and micronaire value in cotton. Frontiers in plant science, (2015); 6: 838.
- Awan M, Abass M, Muzaffar A, Ali A, Tabassum B, et al. Transformation of insect and herbicide resistance genes in cotton (Gossypium hirsutum L.). Journal of Agricultural Science and Technology, (2015); 17(2): 287-298.