

Full Length Research Article
Evolution of Phosphoenolpyruvate carboxylase encoding transcripts in Chickpea (Cicer arietinum L.)
Syed Rehmat Ullah Shah1*, Muhammad Rashid1, Muhammad Abuzar Jaffar1, Muhammad Aslam 2,3, Hinna Shah2, Ghulam Rasool4
Adv. life sci., vol. 6, no. 4, pp. 152-158, August 2019
*– Corresponding Author: Syed Rehmat Ullah Shah (Email: sru.shah@gmail.com)
Authors' Affiliations
2. Faculty of Marine Sciences, Lasbela University of Agriculture, Water and Marine Sciences, Uthal, Balochistan, Pakistan.
3. Department of Chemistry and Biochemistry, Worcester Poly Technic Institute, Massachusetts, USA
4. Department of PBG, Balochistan Agriculture College, Quetta, Pakistan
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Phosphoenolpyruvate carboxylase (PEPC; EC 4.1.1.31) is an important enzyme encoded by a gene family of at least 2-8 plant type and 1-2 bacterial type genes depending upon genome size or species complexity. This enzyme functions as catalyst for the β-carboxylation of phosphoenolpyruvate (PEP) to form oxaloacetate in cytoplasm. It is involved in carbon fixation and various other plant metabolic pathways.
Methods: In this study we characterized the evolutionary perspective of PPC transcripts and their abundance pattern in different plant tissues of chickpea (Cicer arietinum L.).
Results: The current study revealed that PEPC enzyme in chickpea is encoded by a gene family of at least 6 transcripts. All active site residues of C3 PEPCs were found in transcripts. Phylogenetic analysis of the amino acid sequences showed two major groups PTPC and BTPC from different ancestral lineages. Divergence of PTPC in two groups and further convergence within species was found in most of the plants while multiple evolutionary divergences was likely to be specific in legumes including chickpea.
Conclusion: CaPPC genes are regulated under various abiotic stress. Furthermore, the expression pattern of the identified genes can be helpful to explore plant metabolism of chickpea under abiotic stresses, which can be the next step to explore more into this gene family in chickpea.
Keywords: Phosphoenolpyruvate carboxylase transcripts; Chickpea; Phylogeny
Introduction![]()
The phosphoenolpyruvate carboxylase (EC 4.1.1.31) is an important enzyme involved in primary metabolism in bacteria, archaea, algae and vascular plants. It has different catalytic and replenishing roles in C3 plants. It plays the vital role in plant metabolism, such as carbon fixation of C4 and CAM plants, regulation of pH in the cells, act as source of carbon in plant root metabolites specifically in cluster root and nodules [1, 2, 3]. PEPC catalyzes carboxylation of phosphoenolpyruvate (PEP) through bicarbonate (HCO3–) fixation to produce oxaloacetate, followed by reduction to malate or transamination to asparagine via aspartate [4, 5, 6]. PEPC proteins in plants are divided in to two classes. Class I PEPCs are homotetramers of PTPC subunits [3] while class II PEPCs are the hetero-octamers of BTPC and PTPC subunits complex. Class II PEPCs are reported in developing oilseeds of Ricinus communis, developing pollen of Lilium longiflorum and in some green algae [7, 8]. PEPC enzymes are coded by two distinct lineages, named as plant types (PTPCs) and bacterial types (BTPCs) genes [9]. Several candidate genes in PPC families are reported in agricultural plants. The gene family PPC consists of at least 3 genes of PTPC and 1 gene of BTPC in plants [9, 10]. Initial research on Arabidopsis and Oryza sativa have shown that in Arabidopsis there are three PTPC genes named as Atppc1, Atppc2 and Atppc3, containing 10 exons, while the Atppc4 gene, with 20 exons having sequence identity with bacterial PPCs than plant-type PPC [9]. Later research on these genes showed that there are five PTPCs, including 4 cytosolic and 1 chloroplastic and a bacterial type PPC gene in Oryza sativa [11, 12]. In a legume plant, white lupin (Lupinus albus), four PTPCs have been reported [13]. These results reveal that number of PTPCs members vary among the plant species in contrast to previous studies for Arabidopsis and rice [9]. Interaction of these two PEPC subunits PTPC and BTPC were proved at transcriptional and translational levels in Ricinus communis, where interaction of BTPC RcPPC4 and the PTPC gene RcPPC3 results the Class II PEPC protein in developing castor oil seed [7, 14].
Phosphoenolpyruvate carboxylase may have a great role in plant root metabolism, enhanced transcript levels and PEPC activity in cluster roots and nodules [5, 15] suggest its significance in diverse root tissues. In a study on Lupinus albus, it was found that phosphorus supply influenced the transcript levels of PPC that altered the carbon flux in glyceraldehyde 3-phosphate dehydrogenase and glycolytic bypass [16]. In a leguminous plant alfalfa root nodule exposed to elevated level of CO2 resulted improvement in the N2-fixing activity in nodules, where PEPC might have a contribution to malate biosynthesis for N2-fixing bacteroides through facilitating oxaloacetate for N metabolism [17]. It also contributes in different other metabolic processes such as ripening of fruits, pollen germination, stomatal opening, nitrogen metabolism seed development and germination and against different abiotic stresses [18]. Recently introgression of PPC genes from maize into wheat has significantly improved the tolerance against heat and drought [19, 20].
Keeping in view the importance of PPC genes in plant metabolism, in this research we address the number of candidate genes of PPC family in chickpea and their evolutionary history and tissue specific role.
Methods![]()
Sequence analysis
Full length sequences of PPC gene of chickpea were retrieved through “Basic Local Alignment Search Tool” from chickpea transcriptome database (CTDB) (http://www.nipgr.res.in/ctdb.html). Orthologues of CaPPC transcripts from model plant Arabidopsis thaliana were downloaded from TAIR database (https://www.arabidopsis.org). To retrieve PPC gene sequences of legume plant including model plant Medicago truncatula, the blast analysis was performed against each AtPPC gene of the PPC gene family using NCBI database. (http://www.ncbi.nlm.nih.gov).
Phylogenetic analysis and Pairwise comparison
The orthologues of the PPC gene family of different plants were translated into protein and the protein sequences were aligned with the chickpea PPCs using CLC Main Workbench 7 with default settings. Phylogenetic analysis was performed through the Neighbor-Joining algorithm with bootstrapping of 1000 repeats using MEGA5 [21] and a rooted phylogenetic tree was constructed. Pairwise comparisons of Arabidopsis and chickpea PEPC protein sequences were carried out on aligned sequences using CLC Main Workbench 7.
Transcript abundance and hierarchical clustering:
Transcript abundance for each transcript of PPC were retrieved from the chickpea transcriptome database (CTDB)(http://www.nipgr.res.in/ctdb.html) and values as fragments (paired-end reads) per kilobase per million fragments (FPKM) mapped against each contig were calculated.. Transcript level (FPKM) in the different samples were normalized with averages of FPKM values from different tissues and used for heat map generation and hierarchical cluster analysis using CLC Main Workbench 7. Hierarchical cluster analysis was confirmed in SIMCA (Umetrics, Umeå, Sweden).
Results![]()
Phylogenetic relationship of chickpea PPC genes with PPCs in other plants
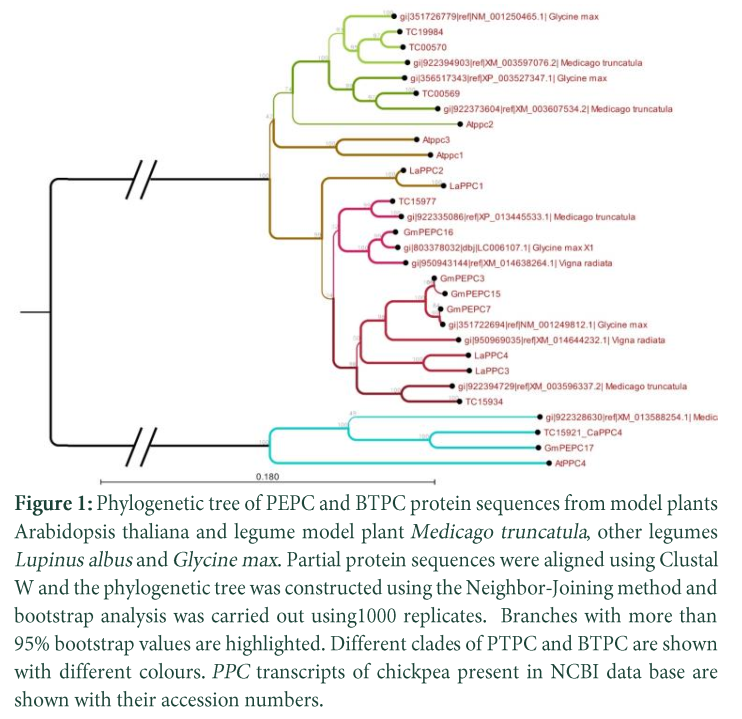
Phylogenetic analysis of full-length protein sequences translated from RNA seq data (Fig.1 & 2) reflected the results of previous studies by separating plant type and bacterial type PPCs of chickpea in separate clades. Phylogenetic tree (Fig. 2) consists of two well-known major clusters of PTPC and a BTPC in all plants. It has been reported that the major lineages are separated before divergence of legumes [ 9, 18, 22]. In the PTPC cluster, Arabidopsis and Lupinus albus were placed in two groups while the legumes such as Medicago truncatula, Glycine max and Cicer aritinum made two major groups including an additional group and further sub-groups within each group. The additional legume specific cluster might have been diverged as different evolutionary lineage. Similar divergence has also been reported in Glycine max [18].
Of the chickpea transcripts, transcript TC15934 shared a group (G.1) with 4 transcripts of Glycine max, 2 of Lupinus albus and 1 of Vigna radiata. Transcript TC15977 shared a group (G. 2) with 1 transcript of each G. max, V. radiata and M. truncatula. Transcripts TC19984 and TC00570 were placed together and made a group (G. 3) with 1 transcript of each M. truncatula and G. max. Transcript TC00569 was placed in a group (G. 4) with 1 transcript of each M. truncatula and G. max. Major cluster of BTPC contained one transcript of each plant species. Arabidopsis and Lupinus albus have the simpler PPC families consisting of two main groups while Medicago truncatula, Glycine max and Cicer aritinum showed great diversity within the PPC family. In all groups two of the legume species Medicago truncatula, Glycine max shared groups with chickpea transcripts, this indicates a close evolutionary history of these species within the legume family.
PTPC and BTPC Transcript model and catalytic domains
The length of the PTPC protein sequence is shorter (963-967) than BTPCs, which are 1043 amino acids longer. In plant and cyanobacteria BTPC are the largest PEPCs of 118 kDa reported to date while PTPC is of 107 kDa [14] but the insertion (residues 328–423) in BTPC in Arabidopsis (AtPPC4) do not match with the insertions observed in the cyanobacterial enzyme [23] which does not prove any co-evolutionary incident with Arabidopsis and cyanobacteria. All the PTPCs and BTPCs showed higher degree of conservation within each type as compared to conserved region between the PTPCs and BTPCs regardless the species differences (Fig. 2). Each type of PPC has also a specific region, missing in either of the other type. These different gene structures of PTPC and BTPC in plants as well as their coding sequences clearly suggest independent origins of these genes [9]. All well-known PEPCs catalytic amino acid residues of the deduced protein sequence were present in PEPC protein sequences of Cicer aritinum. Alignment of the deduced amino acid sequences of PTPCs and BTPCs indicated the occurrence of catalytic residues and domains typical for C3 plants. All the CaPPCs contain alanine instead of the C4 specific serine [24]. The N-terminal PTPC specific phosphorylation site is present in all PTPCs in C. aritinum and Arabidopsis. The HCO3 and PEP binding sites at domain II (GYSDSGKDA) and loop (FHGRGGxxGRGG) are found conserved among the PTPCs and BTPCs [8, 22,25]. The PTPC and BTPC sequences in plants end with last conserved motif Q/RNTG [8, 22]. Alignment of the full-length protein sequences of PTPCs and BTPCs in Arabidopsis and chickpea is shown in supplementary files.
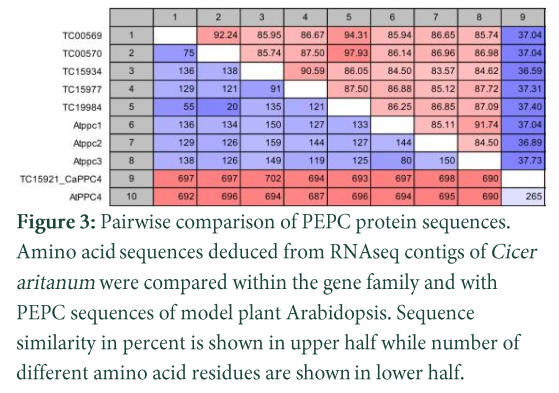
Pairwise comparison of deduced amino acid sequences
Pairwise comparison of full-length protein sequences deduced from PTPC transcripts of Cicer aritinum (Fig. 3) reflected the phylogenetic tree which showed high percentage sequence identity (83-98 %) with difference of 20-159 amino acids when compared within or with
Arabidopsis. Very low percent identity was recorded between all PTPCs and BTPC (<38 %) with >687 amino acid difference. Similar results were also reported in Arabidopsis, Ricinus communis and G. max [9, 18, 22 ]. Within PTPCs in chickpea, transcript TC00570 had highest sequence identity (98%) with TC19984 while lowest identity with transcript TC15934. All others PTPCs are ranged between these two transcripts.
Expression pattern of CaPPCs
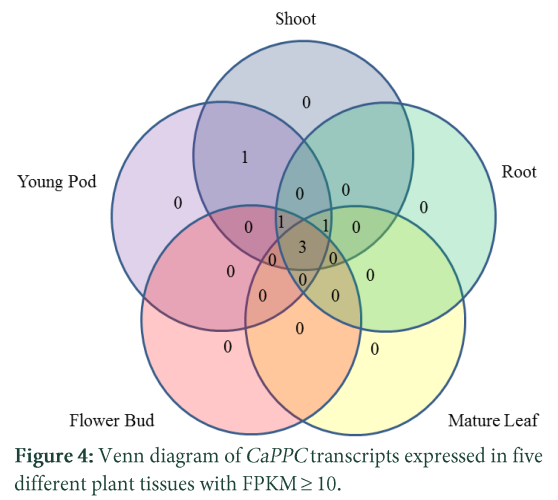
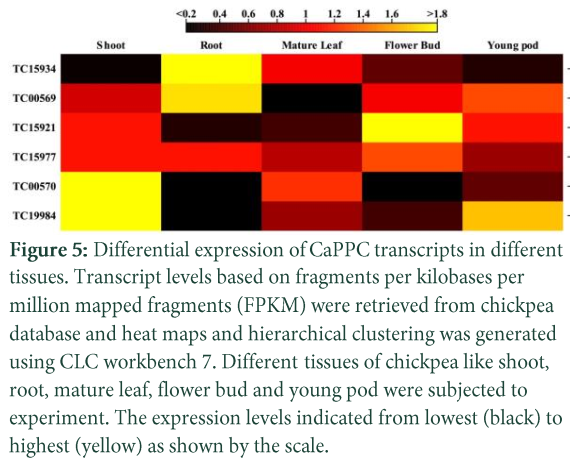
To determine which of the CaPPC transcripts are expressed in more than one tissue, a Venn diagram was constructed (Fig. 4). Three of the transcripts were expressed in all tissues, each of the two transcripts were expressed in three tissues while 1 transcript was expressed in two of the studied tissues. To evaluate the PPC transcript abundance pattern in various tissues, normalized FPKM values of RNA-seq data revealed differential abundance of the PPC transcripts in different tissues of chickpea (Fig. 5). Tissue specific abundance of PPC transcripts indicates different functions of this gene family. The PTPCs transcripts showed higher expression in shoot, root and young pod except TC15977 which had higher expression in flower buds and similarly a BTPC, TC15921 had also higher expression in flower bud. Previously it has been reported that PPC genes were expressed in male reproductive tissues in both lily and Arabidopsis and it was mentioned to have a role in pollen maturation [26]. Two of the transcripts, TC15934 and TC00569 had higher transcript level in roots which indicates role of this gene in organic acid metabolism in roots. Within root system expression of PPC genes may also vary in different developmental stages of roots [5]. In Arabidopsis, AtPPC1 showed higher expression in root [9] while in a recent study AtPPC3 is reported as root specific gene containing citrate binding sites [27]. Two of the transcripts TC00570 and TC19984 showed abundance in shoots which may have different developing organs within shoots. In developing shoots PPC have a role as carbon source of building block for organogenesis of the tissues [22, 28] or high expression in shoot may also have a role in photosynthesis.
Figures & Tables

Discussion![]()
Phosphoenolpyruvate carboxylase is an important enzyme involved in CO2 fixation in photosynthesis in C4 and crassulacean acid metabolism. It plays a role in different metabolic processes in various plant tissues such as fruit, pollen grain, leaf, seed and root in tissue development and against different abiotic stresses [19, 20, 18]. In plants, phosphoenolpyruvate carboxylase comprises a small gene family containing variable number of members in different species such as 4 members in Arabidopsis to 10 members in Glycine max [9, 18].
This study was carried out to investigate that how many members of PPC genes named as CaPPC gene family exist in chickpea and to quantify their transcript levels. We also compared the evolution of PPCs in chickpea being edible representatives of legumes with other plant species. Analysis of RNA sequence data from chickpea database revealed that CaPPC gene family has at least 5 PTPC transcripts and at least 1 BTPC transcript which is much lower number than PPC genes in Glycine max [18]. The possible reason for the discrepancy could be no or lower expression of the transcripts in tissues studied for constructing this transcriptomic data. Arabidopsis and rice PPCs were also consisted of two gene sub-families [9]. Rice has been shown to contain five PTPCs (four cytosolic and one chloroplastic) and one BTPC [ 11, 12] while four PTPCs have been reported in white lupin [13]. Less number of PPC genes in chickpea than soybean could be due to smaller genome size ~738-Mb which is 64-67% of soybean genome [29, 30].
In plants, the plant-type PEPC protein exists as homotetramers (class 1 PEPC) while the bacterial-type PEPCs found in plants exist as heterotetramers of subunits derived from both PTPC and BTPC genes [26]. Co-expression of BTPC, TC15921 and a PTPC, TC15977 in chickpea indicates that these two transcripts may encode for the two sub-units of class II PEPC protein. In R. communis, the BTPC gene RcPPC4 and RcPPC3 had co-expression proposing an interaction of BTPC and PTPC transcripts to participate in a class 2 PEPC protein [7]. Previously in Seabuckthorn PTPC HrPPC5 and BTPC HrPPC4 showed similar transcript patterns [5]. Comparing hierarchical cluster (Fig. 5) and phylogenetic tree (Fig. 6) of CaPPCs revealed that the transcript TC19984 and TC00570 have co-expression and close phylogenetic relationship which specifies that the both regulatory region and coding regions of these transcripts have same evolutionary history.
This study revealed the existence of two diverse types of chickpea PPC genes, PTPC and BTPC, which are diverged as separate evolutionary lineage. A legume specific cluster of PPCs was also separated at early divergence of legumes. CaPPCs include 5 plant type PPCs and at least a bacterial type PPC. Differential expression of CaPPC gene family members occur in various tissues. There are specific transcripts for root, shoot and flower which indicates its diverse role in chickpea.
The authors declare that there is no conflict of interest regarding the publication of this paper.
References![]()
- Shah SRU, Agback P, Lundquist PO. Root morphology and cluster root formation by seabuckthorn (Hippophaë rhamnoides L.) in response to nitrogen, phosphorus and iron deficiency. Plant and soil, (2015); 397: (1-2) 75-91.
- Latzko E, Kelly G. The many-faceted function of phosphoenolpyruvate carboxylase in C3 plants. Physiologie Végétale, (1983); 21: 805-15.
- Izui K, Matsumura H, Furumoto T, Kai Y. Phosphoenolpyruvate carboxylase : a new era of structural biology Annual. Reviews. Plant Biology, (2004); 55: 69-84.
- Plaxton, WC, Carswell, MC. Metabolic aspects of the phosphate starvation response in plants Plant responses to environmental stresses: from phytohormones to genome reorganization Marcel Dekker, New York, (1999); 349-372.
- Shah SRU. Root system of seabuckthorn (Hippophaë rhamnoides L.). Diss. sammanfattning/summary) Uppsala: Sveriges lantbruksuniv., Acta Universitatis agriculturae Sueciae, (2015); 49: 1652-6880.
- Theodorou ME, Plaxton WC. Metabolic adaptations of plant respiration to nutritional phosphate deprivation Plant physiology, (1993); 101: 339-344.
- O’Leary B, et al. Tissue-specific expression and post-translational modifications of plant- and bacterial-type phosphoenolpyruvate carboxylase isozymes of the castor oil plant, Ricinus communis L. Journal of Experimental Botany, (2011); 62: 5485-95.
- O’Leary B, Joonho P, William CP. The remarkable diversity of plant PEPC (phosphoenolpyruvate carboxylase): recent insights into the physiological functions and post-translational controls of non-photosynthetic PEPCs. Biochemical Journal, (2011); 436(1): 15-34.
- Sánchez R, Cejudo FJ. Identification and expression analysis of a gene encoding a bacterial-type phosphoenolpyruvate carboxylase from Arabidopsis and Rice. Plant physiology, (2003); 132: 949-957.
- Mamedov TG, Moellering ER, Chollet R. Identification and expression analysis of two inorganic C‐and N‐responsive genes encoding novel and distinct molecular forms of eukaryotic phosphoenolpyruvate carboxylase in the green microalga Chlamydomonas reinhardtii. The Plant Journal, (2005); 42: 832-843.
- Muramatsu M, Suzuki R, Yamazaki T, Miyao M. Comparison of plant-type phosphoenolpyruvate carboxylases from rice: Identification of two plant-specific regulatory regions of the allosteric enzyme. Plant and Cell Physiology, (2014); 56: 468-80.
- Masumoto C, et al. Phosphoenolpyruvate carboxylase intrinsically located in the chloroplast of rice plays a crucial role in ammonium assimilation Proceedings of the National Academy of Sciences, (2010); 107: 5226-5231.
- Peñaloza E, Muñoz G, Salvo-Garrido H, Silva H, Corcuera LJ. Phosphate deficiency regulates phosphoenolpyruvate carboxylase expression in proteoid root clusters of white lupin Journal of Experimental Botany, (2005); 56: 145-153.
- Gennidakis S, Rao S, Greenham K, Uhrig RG, O'Leary B, et al. Bacterial‐and plant‐type phosphoenolpyruvate carboxylase polypeptides interact in the hetero‐oligomeric Class‐2 PEPC complex of developing castor oil seeds. The Plant Journal, (2007); 52: 839-849.
- Johnson JF, Vance CP, Allan DL. Phosphorus deficiency in Lupinus albus – Altered lateral root development and enhanced expression of phosphoenolpyruvate carboxylase. Plant physiology, (1996); 112: 31-41.
- Peñaloza E, Corcuera LJ, Martinez J. Spatial and temporal variation in citrate and malate exudation and tissue concentration as affected by P stress in roots of white lupin Plant and Soil, (2002); 241: 209-221.
- Fischinger SA, Hristozkova M, Mainassara ZA, Schulze J. Elevated CO2 concentration around alfalfa nodules increases N2 fixation Journal of experimental botany, (2010); 61: 121-130.
- Wang N, Zhong X, Cong Y, et al. Genome-wide analysis of phosphoenolpyruvate carboxylase gene family and their Response to abiotic stresses in soybean. Scientific Reports, (2016); 6: 38448.
- Qi X, Xu W, Zhang J, Guo R, Zhao M, et al. Physiological characteristics and metabolomics of transgenic wheat containing the maize C4 phosphoenolpyruvate carboxylase (PEPC) gene under high temperature stress. Protoplasma, (2017); 254: 1017–1030.
- Qin N, Xu W, Hu L, Li Y, Wang H, et al. Drought tolerance and proteomics studies of transgenic wheat containing the maize C4 phosphoenolpyruvate carboxylase (PEPC) gene. Protoplasma, (2016); 253: 1503–1512.
- Tamura K, Peterson D, Peterson N, Stecher G, Nei M, et al. MEGA5: molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular biology and evolution, (2011); 28: 2731-2739.
- O’Leary B, Rao S, Plaxton W. Phosphorylation of bacterial-type phosphoenolpyruvate carboxylase at Ser425 provides a further tier of enzyme control in developing castor oil seeds. Biochemical Journal, (2011); 433(1): 65-74.
- Sánchez R, Flores A, Cejudo FJ. Arabidopsis phosphoenolpyruvate carboxylase genes encode immunologically unrelated polypeptides and are differentially expressed in response to drought and salt stress. Planta, (2006); 223, 901-909.
- Bläsing OE, Westhof P, Svensson P. Evolution of C4 phosphoenolpyruvate carboxylase in Flaveria, a conserved serine residue in the carboxyl-terminal part of the enzyme is a major determinant for C4-specific characteristics Journal of Biological Chemistry, (2000); 275: 27917-27923.
- Paulus JK, Schlieper D, Groth G. Greater efficiency of photosynthetic carbon fixation due to single amino-acid substitution. Nature Communication. (2013); 4: 1518.
- Igawa T, Fujiwara M, Tanaka I, Fukao Y, Yanagawa Y. Characterization of bacterial-type phosphoenolpyruvate carboxylase expressed in male gametophyte of higher plants. BMC Plant Biology, (2010); 10: 200.
- Connell MB, Lee MJY, Li J, Plaxton WC, Jia Z. Structural and biochemical characterization of citrate binding to AtPPC3, a plant-type phosphoenolpyruvate carboxylase from Arabidopsis thaliana. Journal of Structural Biology, (2018); 204(3): 507-512.
- Sangwan RS, Singh N, Plaxton WC. Phosphoenolpyruvate carboxylase activity and concentration in the endosperm of developing and germinating castor oil seeds Plant physiology, (1992); 99: 445-449.
- Schmutz J, et al. Genome sequence of the paleopolyploid soybean. Nature, (2010); 463: 178-183.
- Varshney RK, Song C, Saxena RK, et al. Draft genome sequence of chickpea (Cicer arietinum) provides a resource for trait improvement. Nature Biotechnology, (2013); 31, 240–246.
![]()
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0