

Full Length Research Article
Molecular cloning and expression of recombinant Trichoderma harzianum chitinase in Pichia pastoris
Muhammad Saleem Iqbal Khan1, Anwar Khan2, Olawale Samuel Adeyinka1, Iqra Yousaf1, Saman Riaz1, Bisma Bashir1, Muhammad Tariq1, Bushra Tabassum*1
Adv. life sci., vol. 7, no. 3, pp. 122-128, May 2020
*– Corresponding Author: Bushra Tabassum (Email: bushratabassum.cemb@pu.edu.pk)
Authors' Affiliations
2. Department of Microbiology, BUITEMS, Quetta – Pakistan
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: The importance of chitinases over the years had attracted huge biotechnological attention because its usage cut across wide range of field. It plays a significant role in the defensive mechanism against fungal pathogens.
Methods: In this study, an endochitinase gene was isolated from Trichoderma harzianum, and characterized in-silico by using various bioinformatics tools. Further, the gene was cloned in eukaryotic expression vector (pPICZA) under the control of AOX1 promoter for recombinant expression in Pichia pastoris GS115 host strain.
Results: The chitinase cDNA was ~1000 bp long, while in-silico studies revealed an open reading frame of 888 bp encoding 295 amino acids with a calculated molecular mass of 37332.76 Da and an estimated isoelectric point of 4.07. Recombinant chitinase protein expressed intracellularly and revealed high expression in P. pastoris host. The 37 kDa recombinant chitinase protein developed with antigen antibody confirmed its expression in P. pastoris.
Conclusion: Conclusively, T. harzianum derived chitinase gene was successfully over expressed in P. pastoris where recombinant protein was expressed intracellular in the form of inclusion bodies.
Keywords: Trichoderma harzianum; Chitinase; Pichia pastoris
Introduction![]()
Chitin is a linear polymer of N-acetyl-β-d-glucosamine (GlcNAc) units linked with β-1-4 glycosidic bond and encompass enormous glycans families [1]. It is a main constituent of cell wall in many microbes and provides strength to the exoskeletons of crustaceans, insects, and fungus cell walls. Scientists over the years have utilized the advantages of chitin hydrolyzing enzymes (endochitinases, exochitinases and N-acetylglucosaminases) to produce many kinds of its derivatives with potential applications in medicine, food, cosmetics, biocontrol, health products and environmental protection [2, 3]. Different enzymes cleave chitin in different manners: Endochitinases randomly digest the chitin chain from β-1,4-glycosidic bonds while exochitinases digest from the nonreducing end of chitin chain and diacetyl-chitobiose (GlcNAc2) is formed; which is converted into GlcNAc by acetylglucosaminases or from the nonreducing end of N-acetyl-chito-oligosaccharides [4].
Several mycoparasites have been identified as bio-control agents against plant phytoparasitic fungi. Trichoderma harzianum is a soil borne and non-pathogenic fungus that has been characterized for its mycoparasitic activities [5, 6]. Mycoparasitism is the defense mechanism present in fungi, in which one fungus kill the other fungus [7]. During this complex defense mechanism, a variety of inhibitory proteins are secreted including ribosome-inactivating proteins (RIP), antimicrobial peptide and pathogenesis related (PR) proteins like chitinases and gluconates which have strong activity against fungal pathogens. The mycoparasitism potential of T. harzianum has been extensively investigated as bio-fungicide to control Fusarium graminearum [8], Alternaria alternate, Fusarium oxysporum [9], Rhizoctonia solani [10] and the root-knot nematode Meloidogyne javanica [11]. Several investigations have focused on the usage and manipulation of chitinase genes to enhance a plant’s ability to resist fungal pathogens. As an extracellular enzyme, chitinase hydrolyzes the chitin present in the cell wall of the phyto-pathogenic fungi. Recently, genes which encode chitinase are being cloned from various microorganisms including Bacillus circulans [12], Streptomyces thermophiles [13], and Serratia marcescens [14] for heterologous expression in E. coli. Heterologous expression of several different proteins in Pichia pastoris (methylotrophic yeast) has been reported as it has certain advantages over E. coli. It is cost effective, gave higher expression levels and have ability to perform many post-translational modifications specific to eukaryotes including glycosylation, proteolytic processing, disulfide bond formation and minimal protein misfolding [15, 16].
Agricultural crops suffer significant yield losses of about 26-30% from fungal pathogens and constitute serious threat to global food security. The incorporation of inhibitory protein genes like chitinases in economically important crop plant will reduce the growth of pathogen, hence increasing tolerance against fungal pathogens. In this study, we cloned T. harzianum cDNA chitinase, amplified by successive PCR and efficiently expressed in P. pastoris GS115. The expression of chitinase protein in a heterologous system permits future functional enhancement in biotechnological application against phytogenic fungi.
Methods![]()
Fungal strain and culture conditions
T. harzianum chitinase producing mutant was obtained from the Institute of Agricultural Sciences, University of the Punjab, Pakistan. Potato dextrose agar (PDA) medium was used to maintain and propagate fungal cultures. The expression vector pPICZA and yeast expression host Pichia pastoris GS115 were obtained from Pichia EasySelect Kit (Invitrogen).
Trichoderma harzianum chitinase amplification and cloning in pCR2.1 vector
T. harzianum hyphae were homogenized in liquid nitrogen with pestle mortar. TRI Reagent (Sigma-Aldrich) was used according to the manufacturer instructions to extract total RNA. The quality of RNA was checked on 1% agarose and proceeded for cDNA synthesis using RevertAid™ H Minus First strand cDNA synthesis kit (Thermos Scientific). The chitinase gene sequence submitted in NCBI GenBank (accession # ARJ31758.1) was used to design forward 5′-CGCCTCGACGCCAGCTTTCTG-3′ and reverse 5′-CGCTGTCGCCAAGTGTCCAGTTA-3′ primer for amplification. PCR reaction was carried out in thermal cycler (ABI; 9700 system). The cycling profile consisted of amplification reaction with following parameters: 5 min initial denaturation at 95ºC followed by 35 cycles with denaturation at 95ºC for 30 sec, annealing at 61ºC for 30 sec and extension at 72ºC for 45 sec followed by final extension at 72ºC for 7 min. The amplicon of the resulting PCR products was detected on 1% agarose and cloned into T-A end of pCR2.1 vector followed by transformation in E. coli top10 cells. Positive clones were confirmed by restriction digestion and sequencing.
Bioinformatic Analysis of Trichoderma harzianum chitinase
The sequence was compared with known sequences in NCBI database using the NCBI BLAST server (http://www.ncbi.nlm.gov/BLAST). For the prediction of TM helix, The Predict protein function was used (https://ppopen.informatik.tu-muenchen.de/) and structural function was predicted by CFSSP Server – Chou & Fasman Secondary Structure Prediction Server (http://www.biogem.org/tool/chou-fasman). The molecular weights and theoretical pIs were predicted using the Compute pI/Mw tool (https://web.expasy.org/compute_pi/). DeepLoc-1.0 (http://www.cbs.dtu.dk/services/DeepLoc/) was combined to predict the subcellular localization of the proteins. Possible protein-polynucleotide binding sites was predicted from https://open.predictprotein.org/. for the analysis of hydrophobicity or hydrophilicity of protein, Protscale (http://web.expasy.org/protscale/) was used and to predict the presence and location of signal peptide cleavage sites. SignalP 4.1 (http://www.cbs.dtu.dk/services/SignalP/) was employed. Domain was identified with Interpro (https://www.ebi.ac.uk/interpro/) and scanprosite https://prosite.expasy.org/scanprosite) was used to identified specific active site. The sequence were align by muscle algorithm and was used to construct phylogenetic trees using MEGA-X.
Construction of recombinant pPICZ-chiF construct and Transformation of Pichia pastoris GS115
For directional cloning of amplified fungal chitinase gene in yeast expression vector, pPICZA, the restriction sites EcoRI and XhoI were added to the forward 5'- CGCGAATTC CTCGACGCCAGCTTTCTG-3´ and reverse primers 5′- CGCCTCGAG TGTCGCCAAGTGTCCAGTTA-3′ respectively. The amplified product was ligated in frame with 6 X His-tag C-terminus for expression in P. pastoris under the control of AOX promoter. The construct was transformed in chemically competent E. coli TOP10 cells and plated on low-salt LB plates (0.5% NaCl, 0.5% yeast extract, 1% bacto-peptone) containing tetracycline (12.5 µg/ml) and zeocin (50µg/mL) at 37ºC overnight. Positive clones were confirmed through restriction digestion. A total of 3µg of recombinant plasmid, pPICZ-chiF was linearized with SacI enzyme and introduced into P. pastoris GS115 strain by transformation in chemically competent P. pastoris, GS115 cells as per Pichia Easy Comp kit manual. Transformants were selected on Yeast Extract–Peptone–Dextrose (YPD) media supplemented with100 µg/µl zeocin. Zeocin+ transformants were replicated further on YPD plates with double concentration of zeocin (200 µg/µl). Subsequently, the white, large colonies were selected and were inoculated in antibiotic free YPD broth and placed at 30ºC for 96 hrs of continuous shaking. After this, cells were streaked onto YPD plates with 100 µg/µl zeocin. Survived colonies were supposed to be with multi copy insertions and hence selected for further protein expression. Microscopic observations were made to verify P. pastoris. The clones were confirmed through PCR amplification with gene specific primers and with promoter AOX specific primers AOXIF:
5′-GACTGGTTCCAATTGACAAGC-3′ and AOXIR: 5′-GCAAATGGCATTCTGACATCC-3′.
Expression and purification of recombinant chitinase in Pichia pastoris
The expression of fungal chitinase in P. pastoris was carried out as per P. pastoris expression manual (Invitrogen) with some optimizations. 250 ml Buffered Glycerol-complex (BMGY) medium (2% peptone, 1% yeast Extract, Yeast Nitrogen base with ammonium sulfate, 100mM potassium phosphate pH 6.0, biotin and glycerol) was inoculated with Positive P. pastoris colony harboring pPICZ-chiF construct and cultured at 30oC in a shaking incubator. The cells were harvested at OD600 of 1 and the pellet was re suspended in 250 ml of Buffered Methanol-complex (BMMY) medium (1% yeast extract, 2% peptone, Yeast Nitrogen base, biotin, 100mM potassium phosphate buffer, pH 6.0, and 0.5% methanol). The cultures were grown at 30°C with continuous shaking at 250 rpm. Further, grown culture was induced with 0.5% methanol (v/v) and samples were taken at 24 hr, 48 hr, 72 hr and 96 hr. Desalting chromatography (Amersham pharmacia) was performed with concentrated supernatant, equilibrated with 20mM Tris–HCl buffer (pH 8.0). Purification of recombinant chitinase protein fractions was done by anion-exchange chromatography Source 30Q (Amersham pharmacia). The purified protein was dialyzed against 20mM sodium phosphate buffer (pH 6.0).
SDS-PAGE and Western blot analysis
Purified recombinant protein was separated on a 10% polyacrylamide slab gel (SDS). SDS-PAGE was further processed for Western blot. Membrane was left in blocking solution for half an hour with gentle shaking. After blocking, the membrane was washed with wash buffer and primary antibody (His probe; G-18) (Santa Cruz) was added onto the membrane and left overnight at 4ºC. Next day, the membrane was washed again and Bovine anti-rabbit IgG-AP conjugated (Santa Cruz) was added as the secondary antibody and incubated for an hour at 37oC with constant shaking. Membrane was washed again, twice, and then substrate (Appendix-6) BCIP/NBT tablets that were specific for alkaline phosphatase conjugated antibodies was added and left in dark at room temperature, until the hybridization signal were detected.
Results![]()
Isolation, cloning of T. harzainum derived chitinase
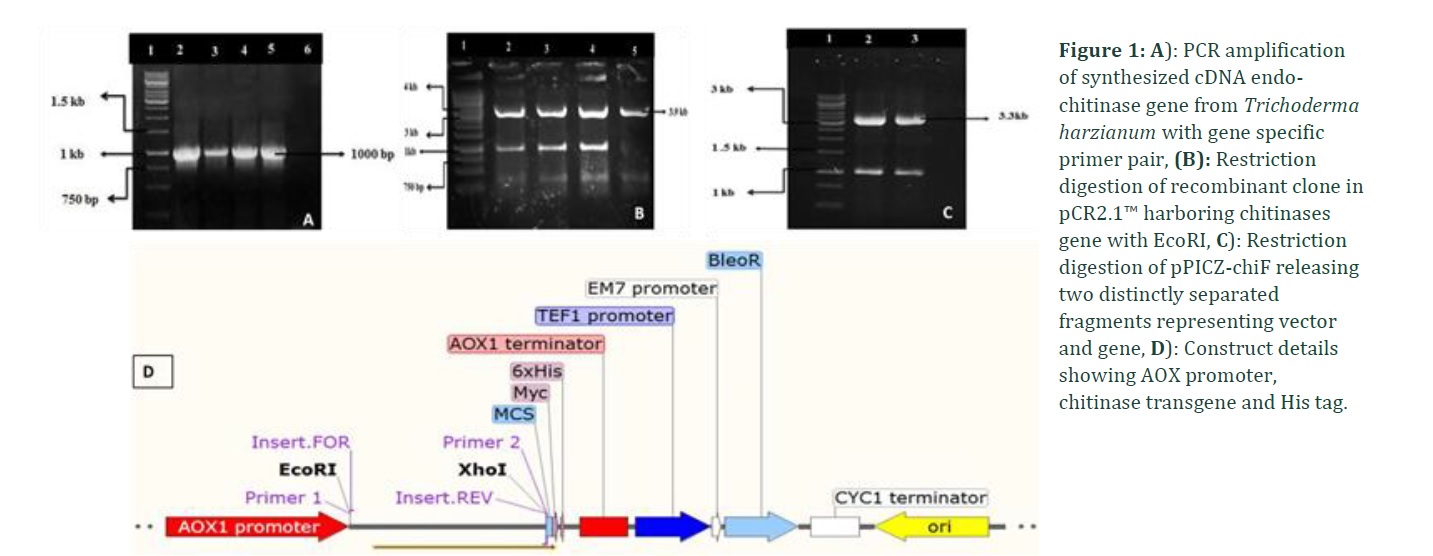
To characterize T. harzianum chitinase, high quality RNA was isolated to synthesize complementary DNAs. The chitinase gene of ~1000 bp was successfully amplified and was cloned in pCR2.1™ vector. The restriction digestion revealed two distinct fragments; ~3.9kb fragment represent pCR2.1™ vector while ~1kb fragment denotes insert (Figure 1A). Out of many screened colonies, only three were positive for insert presence.
Bioinformatic Characterization of Trichoderma harzianum chitinase
The in-silico study revealed that chitinase comprised of an open reading frame of ~888 bp encoding 295 amino acids with a calculated molecular mass of 37332.76 Da and an estimated isoelectric point of 4.07. The evolutionary relationships of chitinase group T. harzianum in the same clades among several Trichoderma sp. (Figure 2). The proportion of various Secondary Structure predicted are: 209 (62.8) % of helix, 162 (48.6%) sheet and 34 (10.2%) turns (Figure 2A). Hydrophobicity plots of the deduced amino acid sequences of putative peptides were hydrophobic, with a high proportion of consecutive cysteine, alanine residues. The hydrophobic domain suggests that chitinase is a membrane protein as part of biological function (Figure 2B). Also, active sites were identified at position 149-157. Signal peptide cleavage site between was at position 21 and 22. Additionally, the classification showed that it belongs to chitinase II of Glycoside hydrolase family 18, catalytic domain within 35 – 324 (Figure 2C). Multiple alignment analysis revealed that the deduced amino acid sequence of chitinase shared a high identity with some eukaryotes (Figure 2D). It is predicted as extracellular, soluble at the subcellular localization. GO term prediction indicated that chitinase mainly involved in biological process, molecular function and not in cellular component.
Recombinant yeast expression vector harboring chitinase
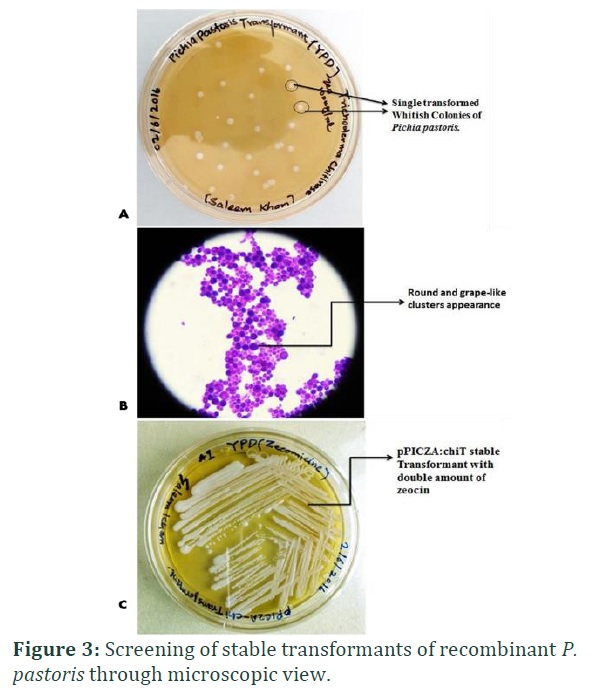
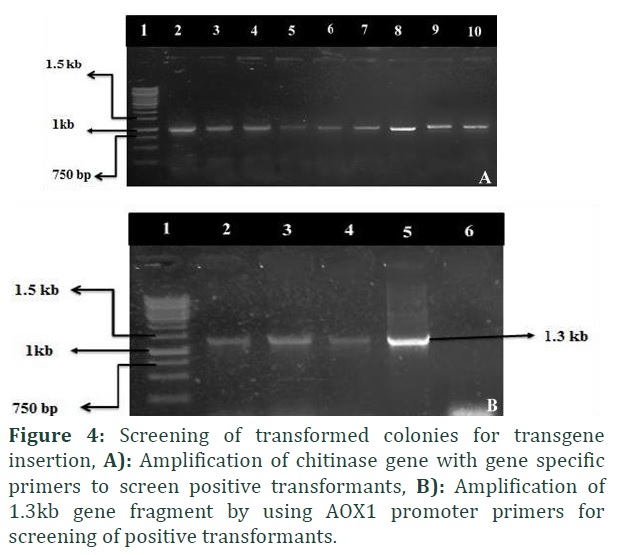
The isolated chitinase gene was subsequently modified by adding up recognition sites of particular restriction enzymes (EcoRI and XhoI at 5′ and 3′ respectively). This was done to clone the chitinase gene directionally in yeast expression vector, pPICZA vector. The positive recombinant colonies were screened initially through microscopic observations as depicted in figure 3. The gene was under the control of the AOX promoter while in fusion with 6 X His tag (Figure 1D). The recombinant plasmid was confirmed through restriction digestion. Appearance of two distinctly separated fragments; ~3.3kb represent vector, pPICZA, while ~1kb DNA fragment represent chitinase gene (Figure 1C). PCR screening of positive transformants harboring recombinant plasmid revealed amplification at ~1 kb when gene specific primers were used (Figure 4A) while AOX1 promoter primers gave amplification at ~1.3 kb (Figure 4B).
Expression of recombinant chitinase in P. Pastoris
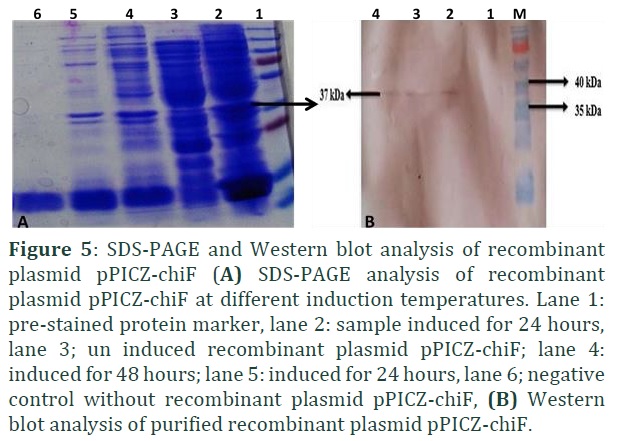
SDS-PAGE revealed that the recombinant chitinase protein was not included in extracellular media when expressed in P. pastoris rather it was found in the form of inclusion bodies. Further, methanol induction resulted in increased expression of recombinant chitinase (Figure 5A). A bright and thick fragment of ~37 kDa was visualized corresponding to mature chitinase in fusion with myc-epitope and detectable by His-Tag 24 hrs post induction. The same recombinant chitinase was further visualized at ~37kDa in Western blotting through specific His-tag primary antibody and AP conjugated secondary antibody. The appearance of purple colored signals in the blot confirmed the presence of chitinases protein (Fig 5B). P. pastoris used as an outstanding host for protein expression in yeast. It is a methylotrophic yeast having tightly regulated promoters and fast growth rate.
Figures & Tables

Discussion![]()
Biocontrol is an alternative approach in modern agriculture to avoid the use of fungicides and pesticides who contributes in uplifting environmental pollution and contamination but also develop resistance in pathogen’s strain which keep on demanding the new formulations [17]. Many chitinase genes have been isolated from Trichoderma spp. and been cloned and characterized against many fungal pathogens like Rhizoctonia solani, Botrytis cinerea and Sclerotinia sclerotium [18]. These chitinases causes the lysis of chitin of plant pathogenic fungi as chitin is the major cell wall component of these mycoparasites [19]. Chitinases are very important biological control agent and belong to family 18 of the glycosyl hydrolase (GH) superfamily [20]. 18 ORF encoding chitinases were reported in database when genome-wide analysis of chitinase gene of Trichoderma sp. was done and these all ORF were belonged to Chitinase family 18 of glycoside hydrolase [21]. It acts on chitin and degrade it to certain monomers. T. harzianum is found in numerous plants: it stimulates root development and plant growth promotion; activate seed germination; increase photosynthetic efficiency and CO2 uptake; ameliorate abiotic stresses and protect the crop against fungal diseases and increase the crop yield. Its protection against fungi has been greatly attributed to the activities of chitinase.
The methylotrophic Pichia pastoris yeast is a new biological tool for the expression of recombinant protein. It is a powerful tool as recombinant protein is derived under AOX1 promoter which expressed the foreign genes quite efficiently and also genetic manipulation is easier than Sachharomyces cerevisiae molecular system [22]. Some integrative vectors lack autonomous replication; however, for successful heterologous gene expression in P. pastoris, the integration of foreign sequences to genome is need to be ensured. This is similar to [23] where recombinant chitinase was express in E.coli and the expressed in pET-28a+ vector for protein expression and got the same ~37kDa band.
The expression of chitin and its reactivity can be enhanced by optimizing pH and temperature upto optimal level of expression as [24]also cloned a novel chi58 chitinase gene which also belongs to chitinase family 18 and by maintaining pH 5.8 and temperature 45oC, they maximize the chitinase activity which was produced and purified by P. pastoris. The expression of chitinase from Trichoderma sp. in P. pastoris will help for large scale production of chitinase for its structural and functional study as well as for its huge production for biocontrol.
Authors' Contribution
MSIK: All the research work/experiments were carried out by this author.
AK: This author helped in the experiments including cloning.
AOS: This author helped in the experiments including real-time PCR assays.
IY: SDS-PAGE has been performed by this author.
SR: This author helped with experiments and edited the manuscript as per format.
BB: This author performed in-silico studies.
MT: This author helped with experiments.
BT: The study was planned and designed by this author. She is the advisor of Mr. Muhammad Saleem Iqbal Khan. This author helped technically and proofread the manuscript.
All the authors declare that they have no competing interest that can negatively affect the current study.
References![]()
- Yu P, Yan Y, Gu Q, Wang X. Codon optimisation improves the expression of Trichoderma viride sp. endochitinase in Pichia pastoris. Scientific Reports, (2013); 3:1–6.
- Tharanathan RN, Kittur FS. Chitin – The Undisputed Biomolecule of Great Potential. Critical Reviews in Food Science and Nutritions, (2003); 43(1):61–87.
- Vyas P, Deshpande M. Enzymatic Hydrolysis of Chitin by Myrothecium verrucaria Chitinase Complex and its Utilization to Produce SCP. Journal of General and Applied Microbiology, (1991); 37:267–75.
- Tanaka T, Fukui T, Imanaka T. Different Cleavage Specificities of the Dual Catalytic Domains in Chitinase from the Hyperthermophilic Archaeon Thermococcus kodakaraensis KOD1. Journal of Biological Chemistry, (2001); 276(38):35629–35.
- Brunner K, Zeilinger S, Ciliento R, Woo SL, Lorito M, Kubicek CP, et al. Improvement of the Fungal Biocontrol Agent. Applied Environmental Microbiology, (2005); 71(7):3959–65.
- Kumar K, Amaresan N, Bhagat S, Madhuri K, Srivastava RC. Isolation and Characterization of Trichoderma spp. for Antagonistic Activity Against Root Rot and Foliar Pathogens. Indian Jounal of Microbiology, (2012); 52(2):137–44.
- Mukherjee PK, Horwitz BA, Herrera-Estrella A, Schmoll M, Kenerley CM. Trichoderma Research in the Genome Era . Annual Review of Phytopathology, (2013); 51(1):105–29.
- Li Y, Sun R, Yu J, Saravanakumar K, Chen J. Antagonistic and Biocontrol Potential of Trichoderma asperellum ZJSX5003 Against the Maize Stalk Rot Pathogen Fusarium graminearum. Indian Jounal of Microbiology, (2016); 56(3):318–27.
- Redda TE, Ma J, Mei J, Li M, Wu B, Jiang X. Antagonistic Potential of Different Isolates of Trichoderma against Fusarium oxysporum, Rhizoctonia solani, and Botrytis cinerea. European Journal of Experimental Biology, (2018); 8(2):1–8.
- Yobo KS, Laing MD, Hunter CH, Morris MJ. Biological control of Rhizoctonia solani by two Trichoderma species isolated from South African composted soil. South African Journal of Plant Soil, (2004); 21(3):139–44.
- Sharon E, Bar-Eyal M, Chet I, Herrara-Estrella A, Kleifeld O, Spiegel Y. Biological control of the root-knot nematode Meloidogyne javanica by Trichoderma harzianum. The American Phytopathological Society, (2001); 91(7):687–93.
- Huang CJ, Chen CY. High-level expression and characterization of two chitinases, ChiCH and ChiCW, of Bacillus cereus 28-9 in Escherichia coli. Biochemical and Biophysical Research Communications, (2005); 327(1):8–17.
- Tsujibo H, Okamoto T, Hatano N, Miyamoto K, Watanabe T, Mitsutomi M, et al. Family 19 Chitinases from streptomyces thermoviolaceus OPC-520: Molecular cloning and characterization. Bioscience, Biotechnology and Biochemistry, (2000); 64(11):2445–53.
- Suzuki K, Sugawara N, Suzuki M, Uchiyama T, Katouno F, Nikaidou N, et al. Chitinases a, b, and c1 of serratia marcescens 2170 produced by recombinant escherichia coli: Enzymatic properties and synergism on chitin degradation. Bioscience, Biotechnology and Biochemistry, (2002); 66(5):1075–83.
- Ciarkowska A, Jakubowska A. Pichia pastoris as an expression system for recombinant protein production. Postepy Biochemii, (2013); 59(3):315–21.
- Cereghino JL, Cregg JM. Heterologous protein expression in the methylotrophic yeast Pichia pastoris. FEMS Microbiology Reviews, (2000); 24(1):45–66.
- Howell CR. Mechanisms employed by Trichoderma species in the biological control of plant diseases: The history and evolution of current concepts. Plant Diseases, (2003); 87(1):4–10.
- Boer H, Munck N, Natunen J, Wohlfahrt G, Söderlund H, Renkonen O, et al. Differential recognition of animal type β4-galactosylated and -fucosylated chito-oligosaccharides by two family 18 chitinases from Trichoderma harzianum. Glycobiology, (2004); 14(12):1303–13.
- Benítez T, Rincón AM, Limón MC, Codón AC. Biocontrol mechanisms of Trichoderma strains. International Microbiology, (2004); 7(4):249–60.
- Mathivanan N, Kabilan V, Murugesan K. Purification, characterization, and antifungal activity of chitinase from Fusarium chlamydosporum, a mycoparasite to groundnut rust, Puccinia arachidis. Canadian Journal of Microbiology, (1998); 44(7):646–51.
- Seidl V, Huemer B, Seiboth B, Kubicek CP. A complete survey of Trichoderma chitinases reveals three distinct subgroups of family 18 chitinases. FEBS Journal, (2005); 272(22):5923–39.
- Cregg JM, Cereghino JL, Shi J, Higgins DR. Recombinant protein expression in Pichia pastoris. Applied Biochemistry and Biotechnology, (2000); 16(1):23–52.
- Yu P, Li JR. Molecular cloning, sequence analysis, expression and characterization of the endochitinase gene from Trichoderma sp. in Escherichia coli BL21. World Journal of Microbiology and Biotechnology, (2008); 24(11):2509–16.
- Wang YJ, Yang Q. Cloning and Expression of a Novel Chitinase chi58 from Chaetomium cupreum in Pichia pastoris. Biochemical Genetics, (2009); 47:547–58.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0