

Review Article
Dog Coat Colour Genetics: A Review
Rashid Saif1,2*, Ali Iftekhar1, Fatima Asif1, Mohammad Suliman Alghanem3
Adv. life sci., vol. 7, no. 4, pp. 215-224, August 2020
*– Corresponding Authors: Rashid Saif (Email: rashid.saif37@gmail.com)
Authors' Affiliations
2. Decode Genomics, 323-D, Town II, Punjab University Employees Housing Scheme, Lahore – Pakistan
3. Department of Biology, Tabuk University – Kingdom of Saudi Arabia
Abstract![]()
Introduction
Methods
Discussion
Conclusion
References
Abstract
Canis lupus familiaris is one of the most beloved pet species with hundreds of world-wide recognized breeds, which can be differentiated from each other by specific morphological, behavioral and adoptive traits. Morphological characteristics of dog breeds get more attention which can be defined mostly by coat color and its texture, and considered to be incredibly lucrative traits in this valued species. Although the genetic foundation of coat color has been well stated in the literature, but still very little is known about the growth pattern, hair length and curly coat trait genes. Skin pigmentation is determined by eumelanin and pheomelanin switching phenomenon which is under the control of Melanocortin 1 Receptor and Agouti Signaling Protein genes. Genetic variations in the genes involved in pigmentation pathway provide basic understanding of melanocortin physiology and evolutionary adaptation of this trait. So in this review, we highlighted, gathered and comprehend the genetic mutations, associated and likely to be associated variants in the genes involved in the coat color and texture trait along with their phenotypes. Moreover, genetic diversity of other associated genes were also pointed out to understand this phenomena in detail along with their genotypes for better understanding the expression and mode of inheritance of this trait for describing dog breeds with more accuracy.
Keywords: Dog breeds; Dog color variants; Dog texture genetics; Dog breed differentiation
Introduction![]()
Coat color and texture are important characteristics of domestic dog breeds. Though the genetic groundwork of coat color has been well characterized, incredibly little is known about the genes persuading coat pattern. In domestic dogs there is a wide range of coat colors, patterns and textures. Conceivably, throughout evolution of the canine from their wild wolf ancestors, coat color in dog breeds have been the unintentional effect of some different selective process [1]. Divergence of the canine into a different type and ultimately separate breeds accelerated color variation as aspects such as camouflage and visibility aided the dogs’ functionality. Coat color and texture of the fur are important characteristics of dog breeds. Studying these coat variations in dog breeds can reveal how the genes responsible for coat colors and texture are transferred and evolved over time giving rise to such great morphological diversity.
Genetics of dog breeds
In the early 1990s, the dog genome project was initiated, motivated by the desire of researchers to discover the genes that led to many of the illnesses that purebred dogs endured. Some dog breeds have been in nature for only a couple hundred years. Most display reduced genetic diversity owing to the common descent of dog breeds from a small number of owners, produced by crossing closely related individuals [2]. In addition, breeds often experience population obstacles as breed popularity waxes and wanes. Modern breeds of dogs have a broad range of colors, patterns and textures. Knowledge of the genetics of canine coat coloring and patterning and coat texturing and length has improved a great deal in recent years. The dog has 39 pairs of chromosomes coming from each parent [3] and a pair of these determines the sex of dog and all other may possess the coat color and texture genes.
Canine genetic diversity
The closest relative to the dog is undeniably the grey wolf. The dog differs by only 0.04% in nuclear DNA coding sequence from its ancestor wolf [4,5]. Mitochondrial DNA (mtDNA) phylogenetics studies between domestic dogs and wolves show multiple backcrossing events that took place between 15,000 and 100,000 years ago [6,7]. One may wonder if today's grey wolf comprises all the variability needed to create the variance found in modern domestic dogs.
The artificial selection for breed standards started during the Victorian era, indicating that astute breeders, novel genes and effective breed selection have all led to the morphological variability found in dogs in the past 200 years [8].
There are over 400 domestic dog species which show distinctive differences in morphology, physiology and behavior [9]. The main reason for phenotypic diversity in dogs is because of two reasons: 1) Initial wolf domestication more than 15,000 years ago [6,7,9] and 2) The development of distinct breeds in the last few hundred years in which humans picked small groups of dogs from the gene pool and actively searched for unique and attractive features [5,9,10].
Variation in Canine genome
Variation in canine genome results from artificial selection and domestication of the dogs. Possible genome variation mechanisms in dog include: mutational hotspots, copy number variants (CNVs), chromosomal fission, and gene duplications, microsatellites or simple sequence repeats, high levels of repeat purity and the plentifulness of SINEC_Cf (carnivore-specific short-interspersed nuclear elements) elements in the canine genome [8,11].
Variation in certain genes are associated with the coat texture and color in dogs. These variations are of great interest for breeders and pet owners. For example, two variants of KRT71 (Keratin-71) gene exists and are found to be associated with curly coat in certain dog species [12,13] and variations in the SGK3 gene are highly associated with hairlessness in Scottish deerhound dogs and American hairless terriers [14,15].
Methods![]()
Literature search strategy and selection criteria
A systematic search was carried out from PubMed, Google Scholar and Google Web Browser by providing key terms “dog, dog coat genetics, dog coat color, dog coat texture, dog genetics, dog coat inheritance etc.”. According to the particular contents further literature was screened and analyzed. In this study 40 research articles were selected to make a comprehensive review.
Discussion![]()
Dog coat texture
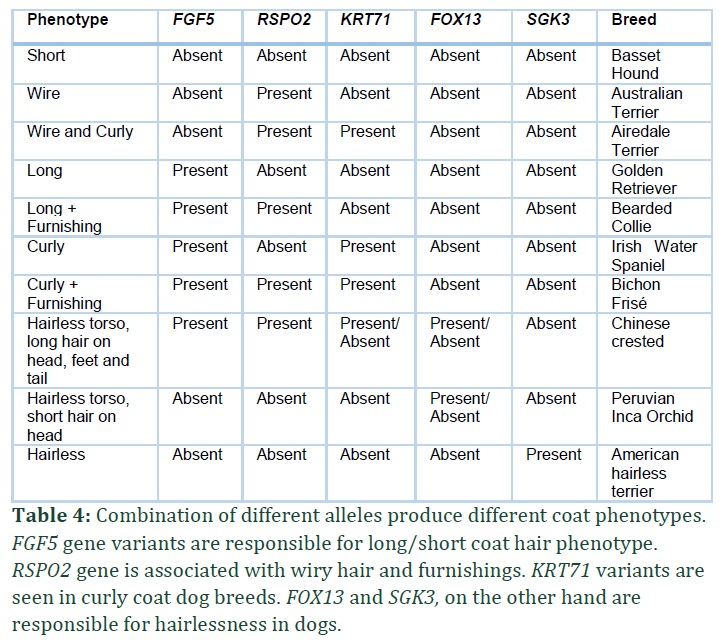
The appearance of a canine coat can be determined not only by the color and distribution of a pigment, but also by the features of the coat such as length, curls or presence or absence of coat hair. Some of the most fascinating advancements in recognizing the evolution of dog variability are the dynamic features of the texture and color of the fur. Coat texture and coat color are most important features of domestic dogs. Canine fur texture, pattern, length and curl are controlled by variation in only three genes and account for almost all coat phenotypes in domestic dogs. These three genes include: RSPO2 (R-spondin–2), FGF5 (fibroblast growth factor–5), and KRT71 (keratin-71) [16].
Genes associated with dog coat texture
Three genes RSPO2, FGF5 and KRT71 are mostly responsible for phenotypic variations in dog coat. A 167 base pair deletion at the 3′ end of RSPO2 is responsible for wiry hair and furnishings (increased hair growth on face and legs) as seen in Portuguese Water Dog and Schnauzer [16,17]. KRT71, on the other hand, is associated with curly hair in dogs [13], however, the variant of the gene is not present in all curly coated hair dogs like curly coated retrievers (CCRs) and later a second variant of KRT71 gene was studied that is found in CCRs associated with curly and wavy hair and can be possibly associated with follicular dysplasia in dogs [12].
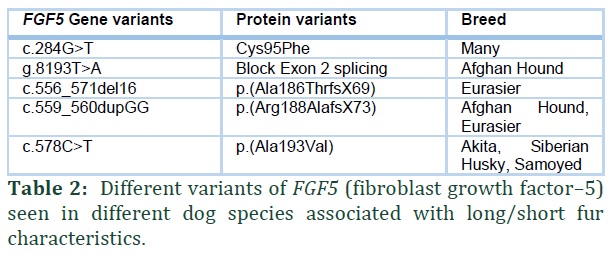
FGF5 gene is associated with long or short hair coat in dog. Examples of FGF5 carrier dogs include Afghan Hound, Eurasier and many other breeds [8,18].
Apart from these three genes there are some other genes associated with hairlessness seen in many dog breeds. SGK3 and FOX13 genes are associated with lack of coat phenotype or hairlessness. FOX13 is studied in Chinese Crested, Peruvian Inca Orchid (Peruvian Hairless) and Mexican Xoloitzcuintle breeds [19] and SGK3 is studied in Scottish Deerhound dogs [15] and American hairless terriers [14].
Gene variants associated with dog coat texture
Genetic variation is seen is the genes of dog associated with coat and fur texture and most of these variants have similar characteristics. Variants of the genes associated with dog coat texture with respect to their chromosome location, mutation and role in dog coat genetics are described below.
Curly coat in dogs
Classification of dogs’ breed on the basis of curly hair is challenging. Large variability of curl types is present. Curly hair coat is a multifarious trait. Dog breeds can be classified into different classes on the basis of coat texture. The large Curly Coated Retriever is a superlative hunting dog who holds all drive and determination that all retrievers retain. Their coat is made of dense mass of small, tight, crisp black and reddish-brown curls. Coat texture in curly hair varies from soft waves to tight curls.
KRT71 gene (R or Cu Locus)
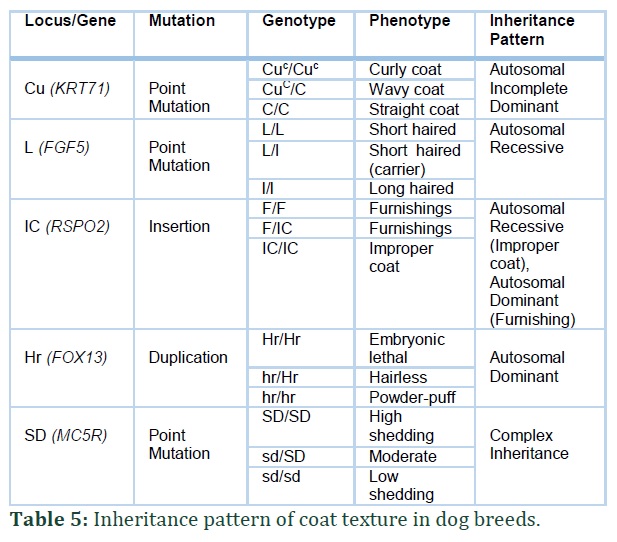
Curly coat type in domestic animals is a relatively common phenotype. KRT71 gene in dogs is found to be associated with curly hair phenotype. Two variants are found in dogs that are associated with curly and wavy hair coat. The variant c.451 C>T is a missense variant in Exon 2 [12] of KRT71 gene and it results in p.Arg151Trp substitution [13,16]. This variant is highly associated with curly coat in Irish Water Spaniel and Standard Poodle [13].
However, during the testing of Curly Coated Retrievers, none of them carried this variant of gene suggesting that there is a second variant of KRT71 gene that is associated with curly coat in CCRs [13]. A 2018 studies found a second variant of KRT71 gene from whole-genome sequencing of Curly Coated Retrievers. This is an indel variant, c.1266-1273 del-in ACA, in Exon 7 and cause frameshift and it results in p.(Ser422ArgfsTer?) (Table 1). This variant is fixed in Curly Coated Retrievers [12].
Hair Length (Long/Short)
Hair length in dogs is usually classified both as long or short and is controlled by two alleles: L > l. L represents short hair and l represents long hair. Hair length is a very important trait and varies among different dog breeds. Hair length variation is mainly controlled by variation in FGF5 gene [8,16,20].
FGF5 gene (L Locus)
Coat appearance is the main morphological distinctions among various dog breeds. FGF5 gene is associated with short or long fur in many dog breeds [8,16,18]. Long coat is inherited in autosomal recessive way [18,20], When two FGF5 carriers are mated, theoretically, 25% long coated offspring will born. Five FGF5 variants studied in different dog breeds, responsible for long/short hair phenotype, are discussed in Table 2 [18].
Wiry hair and furnishings
Wire coats in canine are rough and spiky. It is not soft as like other coat textures. Shedding capability is lost in wire coat texture breeds due to allergy. Dogs with this texture have found another feature that is beard. This additional feature is considered as furnishing. Mutation in RSPO2 signaling protein gene will lead to wiry hair. The insertion of 167 base pairs in this signaling protein gene results in the production of more keratin which ultimately produce more hair.
RSPO2 gene
At the 3′ end UTR of the R-spondin-2 (RSPO2) chromosome (position 11,634,766), a deletion of 167 bp is strongly associated with wiry hair and the furnishings; the latter is indicative of the moustache and eyebrows in many dog breeds [8,17]. This mutation has no impact on the RSPO2 gene protein coding region, but the 3′UTR often codes for elements that affect the stability of mRNA [16,18]. In 2009, Cadieu investigated whether the insertion was linked to a change in RSPO2 expression frequency. In muzzle skin biopsies of dogs with furnishings, they found triple spike in RSPO2 transcriptions [16].
Furnishings (Wh “wire hair” Locus or F Locus)
Furnishing is inherited as an autosomal dominant trait. Furnishing refers to distinct eyebrows and moustaches in certain dog species. It also represents increased hair growth on face and legs. RSPO2 gene is closely associated with furnishing in dogs. Cadieu tested RSPO2 gene in seven dog breeds with distinct eyebrows and moustaches and found a 167 base-pair insertion in untranslated region of the gene. A dog lacking furnishing carries two wild type alleles without any insertion in RSPO2 gene [16]. Furnishing is seen in the following breeds: Brussels Griffon, Chinese crested dog, Dachshund, German Shorthaired Pointer, Irish Soft-Coated Wheaten Terrier
Improper coat (IC Locus)
Insertion in the RSPO2 gene can sometimes result in improper or non-standard coat phenotype in some dog breeds as seen in Portuguese Water Dogs (PWD), the wild type allele results in Improper Coat [17]. Improper Coat is inherited as an autosomal recessive trait. A non-standard fur of short hair on the head, face, and legs characterizes the Improper coat phenotype. There is smooth, straight or moderately wavy hair on the entire body. Dogs with improper coat phenotype are homozygous for wt-allele whereas, dogs with typical coat carries 1 or 2 alleles with insertion.
Wild type allele rate was estimated from randomly selected genotypic samples in the American population of PWD to be 12% [17]. Although Improper coat phenotype do not affect the health of dogs but their breeding and potential of selling offspring is difficult.
Lack of coat / hairlessness
Hairlessness or lack of coat is a very common phenotype seen in several dog species and the best examples include American hairless terrier, Peruvian Inca Orchid and Chinese crested. Hairlessness in dogs mainly results from mutation in two genes: FOX13 and SGK3 [7,14,15,19]. Mutations in these genes result in different levels of hairlessness, most of the times the whole body is hairless. Based on the species, there can be hair tufts on the distal limbs as well as the top of the head. Mutation in these genes may also result in abnormal teeth patterns and missing teeth. However, some seem only to have minor anomalies like fragile toenails [19].
FOX13 gene (Hr Locus)
A mutation in FOX13 results in canine ectodermal dysplasia which is characterized by a lack of hair coat and abnormal teeth. It is a frameshift mutation resulting from a 7-bp duplication in exon 1 of FOX13 gene [19]. Mutations in this gene results in different levels of hairlessness, most of the times the whole body is hairless. Based on the species, there can be hair tufts on the distal limbs as well as the top of the head. Mutation in these genes may also result in abnormal teeth patterns and missing teeth. However, some seem only to have minor anomalies like fragile toenails [19].
Hairlessness is inherited as an autosomal dominant trait and this trait phenotype will only appear in dogs when they carry only 1 mutant allele. The dogs carrying 2 mutant alleles i.e. from both parents die in prenatal stage. Dogs carrying FOX13 gene may have two phenotypes: 1) Hairless Dogs 2) Powder-Puff or Coated [19].
Hairless dogs have a clean smooth body appearance with no hair on body except a crest on head feet or tail. Hairless Dogs carry 1 copy of mutation in FOX13 gene. They have weak and abnormal teeth and neonatal deaths are very common. Powder-Puff carries no mutation in FOX13 gene. The whole body of the dog, in this case, is covered with long silky coat and no notable abnormalities are seen [19].
SGK3 gene
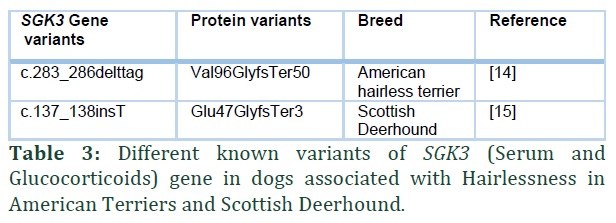
SGK3 is another gene strongly associated with hairlessness in a few dog breeds. Variants of these genes were first studied in American Hairless Terrier [14] and later a new variant of the gene was found to be associated with hairlessness in hairless Scottish Deerhound (Table 3) [15].
American Hairless Terriers
Hairlessness in Hairless American Terrier dog breed is inherited as an autosomal recessive trait and the dog needs to inherit it from both parents. For American Hairless Terriers, homozygosity is not fatal as in other species of hair. This hairlessness mutation is not related to any medical problems affecting the appearance of teeth or fertility of the dog as seen in the case of mutation in FOX13 gene. The dogs with Mutation in SKG13 gene are born healthy and with a light/scattered coat that is lost within the first month of their conception, this condition may be referred to as hypotrichosis [14].
A mutation in Serum and Glucocorticoids (SGK3) resulting in 4 base pair deletion and frame-shift c.283_286delttag was identified in 2017 which results in Hairlessness in American Terrier dog species [14].
Hairless Scottish Deerhounds
Recently a new variant of SKG13 gene has been discovered associated with hairlessness in some dog breeds specially in Scottish Deerhounds [15]. Scottish Deerhound dogs are known to have a 3 to 4 inches long and wiry coat and hairless Scottish Deerhound dogs are extremely rare. In 2019, Parker et.al found a new variant of the gene SGK3 in hairless Scottish Deerhounds after carrying out whole genome analysis. They predicted the inheritance pattern of this mutation to be recessive and both parents should carry the variant allele. The variant is a single base pair insertion in exon 2 of the gene and results in frame-shift c.137_138insT. This mutation leads to early termination of protein and is predicted to be associated with hairlessness in Scottish Deerhound [15].
Association of genes with dog coat phenotypes
There are four main coat characteristics seen in different dog breeds: 1) Hair length (short or long), 2) The presence or absence of curls, 3) Furnishings and Improper coat and 4) Lack of coat (hairlessness). With regard to hair types, various variations of the genes RSPO2 (R-spondin-2), FGF5 (fibroblast growth factor-5), and KRT71 (Keratin 71) were found to be representing, together, the majority of the phenotypes of purebred dogs (Table 4) [16].
Shedding
Shedding is a very common and natural phenomena seen in different dog breeds. However, all dogs do not shed the same. Some dogs shed more hair while others less or not at all. Poodles are known to shed very little. The degree of shedding, as in many instances, is not strictly controlled by one gene but rather by a combination of genes. There is a close relationship between shedding MC5R (SD locus) and furnishing RSPO2 (F locus).
MC5R gene (SD locus)
MC5R gene impacts the level of shedding in different dog breeds [21]. It is very closely linked with the RSPO2 gene. It is a complex inheritance trait and found in all dog breeds.
MC5R gene is normally expressed in hair follicle glands where it produces a waxy substance known as sebum. Sebaceous glands secretions keep the hair flexible and supports lubrication, thermal control and water repellence. Mutation in MC5R gene, which is a point mutation, disrupts the sebum structure resulting in increased shedding [21]. This gene exists in two variants: A and G. G variant is the original natural variant and a result of evolution. Dogs with 1 copy of G variant have high level of shedding whereas dogs carrying 2 copies have much higher levels of shedding.
Dog coat color
Canine coat color is substantiated by the eight genes found in their genome. Variation in these specific genes alter its expression which results in different coat colors. A precise alteration at a single locus of the particular gene affects the coat color which in results classify dog breeds in various classes.
Dog coat color is ruled by their parent genes. There are about 19,000 genes in a dog genome [22]. A limited number of genes control the phenotypic appearance of dogs such as color, patterns and textures. There are several coat colors. Eumelanin: which is black, chocolate brown and greyish in color, Phaeomelanin: containing tan pigment with all shades of red and pink and White: there is no pigment (lack of pigment).
Coat color is systematized by melanin pigment. There are only two basic pigments which result in the color variations in canines. These include Eumelanin – black and Phaeomelanin – red [22]. Deviations in these pigments are responsible for different colors (Figure 3; A-F).
Melanocytes production occur by two types of melanin the one is black/brown photo-protective eumelanin pigment, and the other is red-yellow cytotoxic phaeomelanin pigment. Many melanogenic pathways are controlled by numerous paracrine factors secreted by surrounding keratinocytes by stimulating the alteration among phaeomelanin and eumelanin. In this pathway, microphthalmia transcription factor (MITF) plays a vital role by regulating the expression of the TYR (Tyrosinase), TRP-1 (Tyrosine Related Protein) and DCT (Dopachrome Tautomerase) genes that code enzymes involved in pigment production.
Pigment-type switching
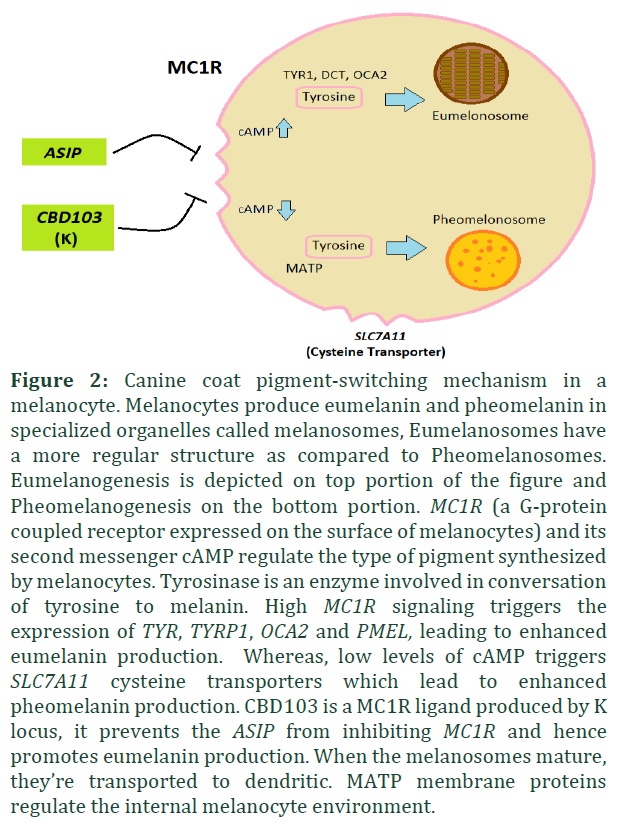
Eumelanin and pheomelanin synthesis is regulated through an intercellular signaling pathway in the hair follicle at a specific location and time. The pigment-type switching gene MC1R and Agouti signaling protein (ASIP) gene are the contributors of this pathway. MC1R is a G-protein coupled receptor expressed on the surfaces of melanocytes and it promotes eumelanin synthesis. This receptor is regulated by ASIP, which inhibits the MC1R signaling pathway promoting the synthesis of pheomelanin. Mutations in ASIP or MC1R genes lead to altered distribution of eumelanin and pheomelanin in dogs. MC1R gain of function mutation results in excessive eumelanin production, whereas ASIP gain of function mutation is responsible for excessive pheomelanin production. As a consequence, ASIP has dominant yellow/red allele and recessive black allele and MC1R has a dominant black allele
Generation of huge range of dog coat colors
Eumelanin and phaeomelanin produce a wide variety of dog coat colors in all various combinations (Figure 3; A-E) [23]. White hair on dogs tends to occur when the cells do not produce any pigment (Figure 3; F). This impacts some areas of the dog’s coat most often. When eumelanin is not produced in the nose, the dog may end up with a pink nose or if it is not present in eyes it results in blue eyes. In rare cases, the whole body of dog is affected which results in albino dogs [24].
Gene variants associated with Canine coat color diversity
There are 8 major genes and their variants responsible for huge diversity in dog coat color. Dog coat color genes can be characterized into Pigment Dilution genes (C Locus, B Locus, D Locus, M Locus and G Locus) and the Pigment switching genes (A Locus, E Locus and the K Locus). Some of the genes associated with some Loci (like C Locus or G Locus) are not completely investigated yet. Dog coat color genes and their variants are discussed further:
ASIP gene (A or Agouti locus)
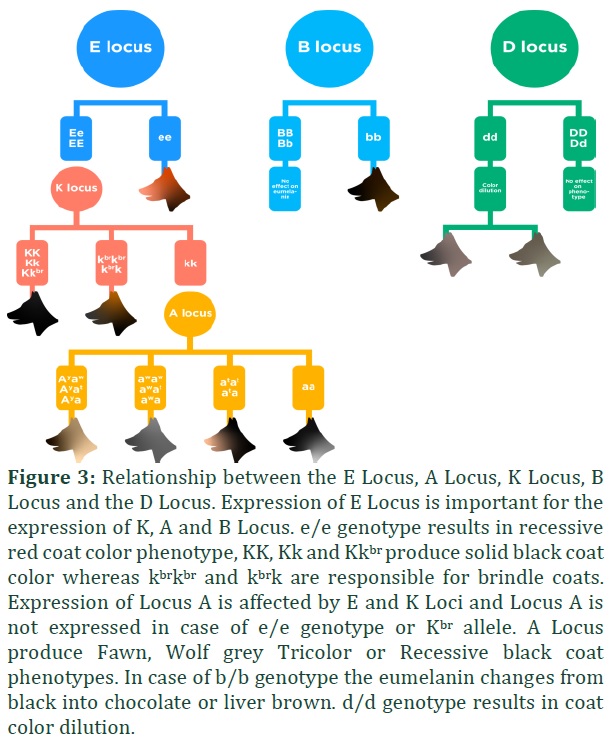
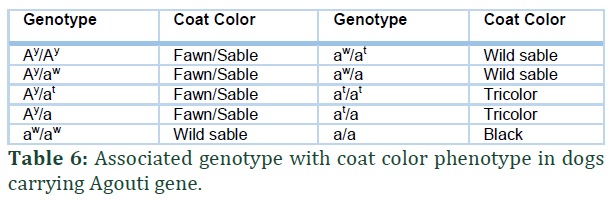
ASIP (Agouti signaling protein) gene is found in chromosome 24 of dogs. There are four known alleles of this gene and in the following hierarchy Ay > aw > at > a [25]. Locus E (MC1R) and Locus K (CBD103) affect the expression of locus A (Figure 2). In the presence of e/e genotype and KB allele of aforementioned Locus/genes respectively, A Locus is not expressed [25-28].
If allele ay is present in dog then the dog will have fawn coat color and the presence of other alleles does not matter. This suggests that the allele ay is dominant over other alleles of the gene. If aw allele is present in combination with aw/aw, aw/at or aw/a the dog will have a wild sable coat color suggesting that the allele aw is dominant in this case over a and at. In the case of at/a or at/at the dog will have a tan or black or tricolor phenotype. The allele a is recessive and dogs with genotype a/a have black coat color (Table 4).
The aw allele is the wild type allele and can be traced in wild wolfs and coyote [25,28] and causes the eumelanin to change in phaeomelanin which is responsible for agouti (like the color of grey wolf) color. A SINE insertion has been identified to be present in intron 1 of ASIP gene at allele and the a allele and absent in aw and ay alleles. The ay allele developed independently and two mutations c.246G>T (A82S) a c.250G>A (R83H) in this allele differentiate it from aw allele [25]. The at allele is only different from the aw allele because of the presence of the SINE insertion and the recessive allele a is caused as a result of SINE insertion as well as c.286C>T (p.R96C) in exon 4 [27] (Figure 3; G-J).
TYRP1 gene (B or brown locus)
TYRP1 gene (Tyrosine related protein 1) is mapped in the chromosome 11 of the canine genome and it is involved in the production of brown or black eumelanin. Locus B is also referred to as Live gene because of liver brown color phenotypic characteristics. Locus B has 2 known alleles: allele B and allele b which controls the black color of coat and nose and the brown color of coat and nose respectively (Figure 3; K-L). The color mutation of Locus B is only effective on dogs that carry E/E or E/e on Locus E (Extension) [29]. In some dog breeds, like the French bulldog, the exact mutation responsible for chocolate brown color is not exactly known yet.
The wild type allele B is responsible for the black coat color and it is dominant over allele b (Figure 2). Dogs with allele B will have black coat color and black nose. The brown eumelanin is only produced when two b alleles are present (Figure 3; K-L). For brown coat color three variants of the TYRP1 gene are discovered: bs (c.991C>T), bc (c.121T>A) and bd (c.1033_1035del) and the fourth variant baus c.555T>G.c is only found in Australian Shepherd breeds.
MLPH gene (D or dilution locus)
MLPH gene (Melanophilin protein) is mapped in the chromosome 25 of the dogs and the mutation is responsible for lighter or diluted coat colors (Figure 3; M-N). The recessive alleles of this gene are represented with d. Variations in MLPH gene affects the melanosomes transport in melanocytes resulting in a lighter coat color or dilution (Figure 3; M,N) [29].
There are two allelic variants of D locus: d1 (c.-22G>A) and a recently discovered allele d2 ( c.705G>C) [30]. Diluted color is inherited as an autosomal recessive trait. The d1 variant is seen in the following breeds: Labrador Retriever, Australian shepherd and German dog etc. and the variant of d2 is studied in Chow-Chow, Sloughi, French bulldog, Papillon and Thai Ridgeback. The non-diluted allele, in case of dogs with no dilution, is the dominant D allele. The genotype of dog carrying D locus allele could be homozygous as d1/d1 or d2/d2 or it can be compound heterozygous d1/d2 [3].
MC1R gene (E or extension locus)
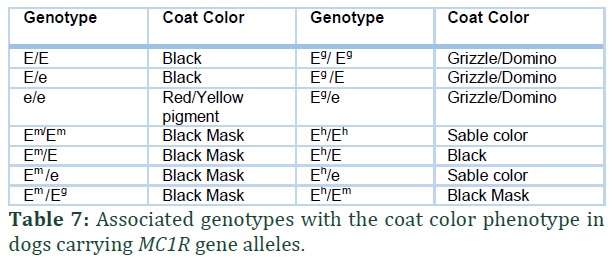
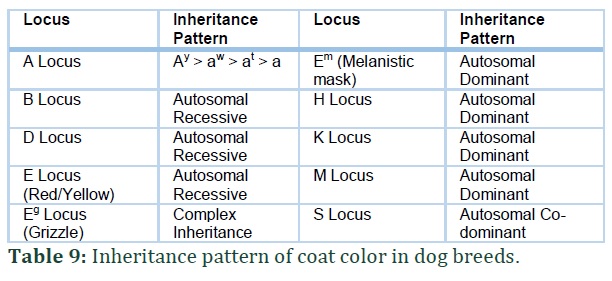
MC1R gene (Melanocortin 1 receptor) is mapped in the chromosome 5 of canine genome [29,31] and controls the production of pigment in melanocytes (Figure 3; O-S). Melanocytes control dog coat color. The wild type dominant allele of the gene is E which produces eumelanin (black pigment). Other alleles in the E locus series include Em (black mask on face), e (recessive allele) and the newly discovered alleles Eg and Eh.
The allele e is created by a mutation in the gene c.916C>T that turns the eumelanin to phaeomelanin in the coat [31,32] this recessive form of allele results in bright red or yellow coat color in many dog breeds. The Em allele is a result of point mutation c.790A>G and it is responsible for melanistic mask. The expression of this allele depends on the alleles on Locus A and Locus K. Dogs that are black or any dark colored dogs have a melanistic mask that is hard to distinguish and the melanistic mask may not always be black as seen in many dog breeds. Dog breeds showing Em allele characteristics include: German Shepherd, Afghan and Staffordshire Bull Terrier. EG allele can result in grizzle coloration in Saluki dog breed as well as domino color in Afghan hounds. This allele results as a mutation c.233G>T in the gene MC1R and this mutation is seen in both the dogs. The EG allele grizzle and domino coloration only appear if the dog is carrying two at alleles and stays hidden if the dog carries ay allele. The Eh allele results from c.250G>A mutation and inhibits eumelanin and it is studied in the dogs with sable phenotype. The hierarchy of Locus E is: Em > Eg > E > Eh > e (Table 5).
CBD103 gene (K or dominant black locus)
The CBD103 (Beta-defensin gene) is mapped in the chromosome 16 of canine genome and is responsible for black coat color [33]. Three alleles in the K locus are discovered: KB (Dominant Black), Kbr (Brindle) and Ky (Recessive yellow or non-solid black). The K Locus is linked with the E Locus and is not expressed if the E Locus genotype is e/e and requires at least one wild type E allele (Figure 2) [34,35]. KB is a dominant allele and it inhibits the Agouti (A Locus) and results in solid black coat color phenotype. Kbr is another mutation in the K locus gene, however, it allows the expression of Agouti (A Locus) and results in brindling. This allele is dominant over Ky allele but recessive with KB. Ky allele is the third allele of K Locus and allows the expression of Agouti locus too but without brindling. In homozygous state i.e. Ky/Ky locus A is responsible for dog coat color but since this allele is recessive over other two alleles, in any other genotypic form the A locus is not fully expressed as it is.
TYR and SLC45A2 genes (C or colored/chinchilla locus)
The TYR (Tyrosinase) gene mapped in chromosome. 21 of canine causes Albinism in many animal species and is thought to be responsible for white coat phenotype in dogs. The mutations in this gene is not completely studied yet. Five alleles of C locus are theorized: C (normal wild type allele expressing phaeomelanin, results in deep intense coat), cch allele causes partial inhibition of phaeomelanin production in coat, ce completely inhibits phaeomelanin production and results in a white coat color, cb, cd and ca alleles results in albino like appearance in dogs. None of these alleles are completely studied yet and some experimental works (cDNA study of albino and white coated dogs) suggest no polymorphism in this gene [36].
A second gene SLC45A2 that is found to be involved in albinism in humans is thought to the gene resulting in C locus in dogs. Experimental works show a homozygous deletion in white Dobermans dogs in this gene [36]. Lhasa Apso breed shows the characteristics of Albinism but does not carry mutation in TYR and SLC45A2 genes but a second mutation in SLC45A2 gene leaving a room open for much experimental work to be done on C locus.
SILV or PMEL17 gene (M or merle locus)
SILV gene (Silver Locus protein) formerly known as the PMEL17 gene (Pre-melanosome protein) is linked with some health issues in dogs and is responsible for irregular diluted or solid colored patches on dog coat (Figure 3; T-W). Merle gene has been identified in many dog species but this specific coat pattern phenotype occurs in only few dog breeds like Australian Shepherd and Border collie etc. Merle gene only dilutes eumelanin pigment. A black coated dog showing merle patterns is known as a Blue Merle and brown dog having merle pattern is known as Red Merle [37].
There are two alleles of merle gene: M and m whereas, m is the normal wild type allele. Merle mutation results as a SINE insertion in the SILV gene this insertion is a PolyT insertion of a variable length and the length of this insertion determines the merle phenotype in dogs (Table 6). Merle is inherited as an autosomal, incomplete trait. In the case of Mx allele the x represents the length of PolyT sequence insertion. There are 25% chances of a M/M dog to be born when two M carries are bred, this is “Double Merle” and the dogs will have hearing or vision deficiencies. Hidden merle is when the merle is not recognizable phenotypically but the dog is genetically Mx.
PSMB7 gene (H or harlequin locus)
The PSMB7 gene is a modifier gene of merle SILV, together these two loci control the harlequin phenotype (white coats with black spots or patches all over) in some dog species i.e. Great Dane (Figure 3; X). H Locus gene has been mapped in the chromosome. 9 of canine. H locus results from a point mutation in the 20S proteasome β2 subunit at the H locus [37-39]. There are two alleles of H locus: H and h, H allele results from the point mutation and the h allele is the wild type allele. Dogs will not have harlequin coat phenotype if they do not carry the merle mutation. H/H genotype is lethal for dogs and the dog will die in embryonic stage.
MITF gene (S or white spotting locus)
The MITF gene (Microphthalmia Associated Transcription Factor) is responsible for irregular white patches or white spotting on dog body. White coloration in dogs is not caused by lack of eumelanin or phaeomelanin pigments but white dogs lack any pigment. The MITF gene is responsible for the development of melanocytes and the white pigmentation results from the lack of melanocytes. There are 4 known alleles of the S locus: S Locus (Solid color with no white patterns), si (Irish spotting, white spots often seen as white collar, belly and legs), sw (Extreme white spotting) and sp (Piebald). S allele is the wild type and results in no white pigmentation whereas a SINE insertion mutation is responsible for other allele variants. There are two MITF gene promoters that carry mutations, the MTIF-M promoter has three mutations (SINE insertion, SNP and LP-Variable Length Polymorphism) and the MITF-1B promoter has one mutation which is the 12bp deletion in exon 1B. Mutations in several MITF gene associated regions results in white spotting. Piebald means irregular spotting and the amount of white a dog expresses as phenotype depends on the number of allele variants a dog carry and Piebald allele is recessive to the S allele. g.21,836,232_218,364,27ins>del mutation in the MITF gene results in the insertion of SINE-Cf element that results in extremely white spots sw/sw or piebald sp/sp. This mutation is not seen in the case of S and si alleles [40].
Coat color and texture variability in Pakistani dog breeds
Pakistani dog breeds are found to be different from foreign dog breeds not only in their coat color but also in some other characteristics. Pakistani Mastiff is black, brown, fawn and brindle. It requires minimal grooming. Seasonal flea treatment is necessary. This is one of the superlative selections if there is no time, skill or money to take care of a high maintenance dog. CNVs and SNPs leads to the variety of coat color, patterns and texture in dog breeds of Pakistani and other countries due to the environmental and physiological conditions.
Bakharwal Dog, an indigenous Pakistani dog breed has dense straight hair and supposedly carries l/l genotype of FGF5 gene. Whereas Bully Kutta and Gaddi Dog, 2 Pakistani dog breeds with straight medium-length coat hair. Most of the dog breeds in Pakistan have straight coat hair as a result of natural breeding and since it is more suitable for Pakistani environmental conditions and it also saves grooming costs.
In Pakistan there is a large number of street dogs (free-ranging urban dogs) including Pakistani Mastiff, Gaddi Kutta and Indian Pariah dogs. The major reason for existence of huge number of stray dogs in Pakistan is unplanned breeding. While this leads to a huge variability in coat texture and color among dog breeds but mutations may lead to various other genetic defects. Large amount of garbage exposed on streets and different areas enable stray dogs to survive and reproduce. People would also feed stray dogs and keep them as “free roaming pets”. These factors contribute to huge coat variability among Pakistani dog breeds.
Adopting a dog in Pakistan depends on a large number of factors. Most often people would keep dogs to guard their livestock like the Gaddi Kutta. Temperature is a huge factor that is considered before adopting any dog breed. Dogs that are easy to maintain and don’t require grooming very often are preferred because of economic state of people and availability of services.




Breeding for a particular trait in dogs would be quite expensive and a tedious job. The progeny needs to reach their maturity level only after which the determination of the success of the cross could be carried out. If the trait is very complex, a lot of time and effort would be needed to achieve a desirable outcome. Marker-Assisted Selection (MAS) is used to help reduce the amount of time required to determine whether the progeny have the trait or not. Another advantage is that it’s a cost-effective technique with reference to screening of traits. By detecting the unique trait at the level of DNA, identification of positive selection can occur quite early. Another advantage of this technique is that these markers are not affected by environmental changes.
All authors contributed equally to this review.
The authors declare no conflict of interest relevant to this review article.
References ![]()
- Hédan B, Corre S, Hitte C, Dréano S, Vilboux T, et al. Coat colour in dogs: identification of the merle locus in the Australian shepherd breed. (2006); 2(1): 9.
- Ostrander EAJAS. Genetics and the Shape of Dogs: Studying the new sequence of the canine genome shows how tiny genetic changes can create enormous variation within a single species. (2007); 95(5): 406-413.
- Kaelin CB, Barsh GSJTGotD. Colour, Texture and Length in the Dog. (2012); 57.
- Bardeleben C, Moore RL, Wayne RKJMp, evolution. A molecular phylogeny of the Canidae based on six nuclear loci. (2005); 37(3): 815-831.
- Lindblad-Toh K, Wade CM, Mikkelsen TS, Karlsson EK, Jaffe DB, et al. Genome sequence, comparative analysis and haplotype structure of the domestic dog. (2005); 438(7069): 803.
- Savolainen P, Zhang Y-p, Luo J, Lundeberg J, Leitner TJS. Genetic evidence for an East Asian origin of domestic dogs. (2002); 298(5598): 1610-1613.
- Wayne RK, Leonard JA, Vilà CJDdNg, paradigms a. Genetic analysis of dog domestication. (2006); 279-293.
- Shearin AL, Ostrander EAJPb. Canine morphology: hunting for genes and tracking mutations. (2010); 8(3): e1000310.
- Vaysse A, Ratnakumar A, Derrien T, Axelsson E, Pielberg GR, et al. Identification of genomic regions associated with phenotypic variation between dog breeds using selection mapping. (2011); 7(10): e1002316.
- Vilà C, Savolainen P, Maldonado JE, Amorim IR, Rice JE, et al. Multiple and ancient origins of the domestic dog. (1997); 276(5319): 1687-1689.
- Minnick MF, Stillwell LC, Heineman JM, Stiegler GLJG. A highly repetitive DNA sequence possibly unique to canids. (1992); 110(2): 235-238.
- Bauer A, Hadji Rasouliha S, Brunner M, Jagannathan V, Bucher I, et al. A second KRT 71 allele in curly coated dogs. (2019); 50(1): 97-100.
- Salmela E, Niskanen J, Arumilli M, Donner J, Lohi H, et al. A novel KRT71 variant in curly‐coated dogs. (2019); 50(1): 101-104.
- Parker HG, Harris A, Dreger DL, Davis BW, Ostrander EAJPTotRSBBS. The bald and the beautiful: hairlessness in domestic dog breeds. (2017); 372(1713): 20150488.
- Parker HG, Whitaker DT, Harris AC, Ostrander EAJGG, Genomes, Genetics. Whole Genome Analysis of a Single Scottish Deerhound Dog Family Provides Independent Corroboration That a SGK3 Coding Variant Leads to Hairlessness. (2019).
- Cadieu E, Neff MW, Quignon P, Walsh K, Chase K, et al. Coat variation in the domestic dog is governed by variants in three genes. (2009); 326(5949): 150-153.
- Parker HG, Chase K, Cadieu E, Lark KG, Ostrander EAJJoh. An insertion in the RSPO2 gene correlates with improper coat in the Portuguese water dog. (2010); 101(5): 612-617.
- Housley D, Venta PJAg. The long and the short of it: evidence that FGF5 is a major determinant of canine ‘hair’‐itability. (2006); 37(4): 309-315.
- Drögemüller C, Karlsson EK, Hytönen MK, Perloski M, Dolf G, et al. A mutation in hairless dogs implicates FOXI3 in ectodermal development. (2008); 321(5895): 1462-1462.
- Dierks C, Mömke S, Philipp U, Distl OJAg. Allelic heterogeneity of FGF 5 mutations causes the long‐hair phenotype in dogs. (2013); 44(4): 425-431.
- Hayward JJ, Castelhano MG, Oliveira KC, Corey E, Balkman C, et al. Complex disease and phenotype mapping in the domestic dog. (2016); 710460.
- Buzhardt L. Genetics Basics – Coat Color Genetics in Dogs. (2016).
- Parker HG, Kim LV, Sutter NB, Carlson S, Lorentzen TD, et al. Genetic structure of the purebred domestic dog. (2004); 304(5674): 1160-1164.
- Caduff M, Bauer A, Jagannathan V, Leeb TJPo. OCA2 splice site variant in German Spitz dogs with oculocutaneous albinism. (2017); 12(10): e0185944.
- Berryere TG, Kerns JA, Barsh GS, Schmutz SMJMG. Association of an Agouti allele with fawn or sable coat color in domestic dogs. (2005); 16(4): 262-272.
- Dreger DL, Schmutz SMJJoH. A SINE insertion causes the black-and-tan and saddle tan phenotypes in domestic dogs. (2011); 102(Suppl_1): S11-S18.
- Kerns JA, Newton J, Berryere TG, Rubin EM, Cheng J-F, et al. Characterization of the dog Agouti gene and a nonagoutimutation in German Shepherd Dogs. (2004); 15(10): 798-808.
- Schmutz SM, Berryere TG, Barta JL, Reddick KD, Schmutz JKJJoh. Agouti sequence polymorphisms in coyotes, wolves and dogs suggest hybridization. (2007); 98(4): 351-355.
- Schmutz S, Berryere TJAg. Genes affecting coat colour and pattern in domestic dogs: a review. (2007); 38(6): 539-549.
- Bauer A, Kehl A, Jagannathan V, Leeb TJAg. A novel MLPH variant in dogs with coat colour dilution. (2018); 49(1): 94-97.
- Schmutz S, Berryere T, Ellinwood N, Kerns J, Barsh GJJoH. MC1R studies in dogs with melanistic mask or brindle patterns. (2003); 94(1): 69-73.
- Newton J, Wilkie AL, He L, Jordan SA, Metallinos DL, et al. Melanocortin 1 receptor variation in the domestic dog. (2000); 11(1): 24-30.
- Candille SI, Kaelin CB, Cattanach BM, Yu B, Thompson DA, et al. A β-defensin mutation causes black coat color in domestic dogs. (2007); 318(5855): 1418-1423.
- Kerns JA, Cargill EJ, Clark LA, Candille SI, Berryere TG, et al. Linkage and segregation analysis of black and brindle coat color in domestic dogs. (2007); 176(3): 1679-1689.
- Korec E, Hančl M, Bydžovská M, Chalupa O, Korcová JJBg. Inheritance of coat colour in the cane Corso Italiano dog. (2019); 20(1): 24.
- Winkler PA, Gornik KR, Ramsey DT, Dubielzig RR, Venta PJ, et al. A partial gene deletion of SLC45A2 causes oculocutaneous albinism in Doberman pinscher dogs. (2014); 9(3): e92127.
- Clark LA, Wahl JM, Rees CA, Murphy KEJPotNAoS. Retrotransposon insertion in SILV is responsible for merle patterning of the domestic dog. (2006); 103(5): 1376-1381.
- Clark LA, Starr AN, Tsai KL, Murphy KEJG. Genome-wide linkage scan localizes the harlequin locus in the Great Dane to chromosome 9. (2008); 418(1-2): 49-52.
- Clark LA, Tsai KL, Starr AN, Nowend KL, Murphy KEJG. A missense mutation in the 20S proteasome β2 subunit of Great Danes having harlequin coat patterning. (2011); 97(4): 244-248.
- Schmutz SM, Berryere TG, Dreger DLJJoH. MITF and white spotting in dogs: a population study. (2009); 100(suppl_1): S66-S74.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0