

Full Length Research Article
The effect of wildfires on wood-decay fungi in the Great Smoky Mountains National Park
Nawaf I. Ibrahim1, Abdel Moneim E. Sulieman1*, Sami Hannachi1, Steven L. Stevenson2
Adv. life sci., vol. 8, no. 1, pp. 73-77, November 2020
*– Corresponding Author: Abdel Moneim E. Sulieman (Email: abuelhadi@hotmail.com)
Authors' Affiliations
2. Department of Biological Sciences, University of Arkansas, Fayetteville, Arkansas – USA
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: In late November and early December of 2019, wildfires occurred over portions of the Great Smoky Mountains National Park. More than 4,000 ha were affected by the wildfires. The objective of the research reported herein was to access the impact of these wildfires on the assemblages of wood-decay fungi associated with the study areas in the park.
Methods: The investigated fungi were confined to two study areas; the first subjected to a relatively high intensity burn and the second subjected to a relatively low intensity burn. In addition to specimens of fungi obtained in the field, small pieces of coarse woody debris were assembled, brought back to the laboratory and placed in plastic chambers for incubation and kept moist. Over the course of two months, fruiting bodies appearing in these incubation chambers were observed and collected. All specimens from both the field and incubation chambers were identified from sequences of the internal transcribed spacer (ITS) region of the ribosomal DNA.
Results: A total of 31 different taxa were identified along with nine taxa reported previously as unknown and uncultured. However, almost all of these records were from the study area subjected to the relatively low intensity burn.
Conclusion: The relatively high intensity burn site was almost completely devoid of wood-decay fungi.
Keywords: Great Smoky Mountains National Park; ITS ribosomal DNA region; Wood-decay fungi
Introduction![]()
The woody decay matter is vital to the general timberland biological system. By this process, the arrival of minerals and carbon counterbalances the opposite activities of carbon assimilation and nutrient absorption by living trees. For a proper understanding of the ecology of an area, the two perspectives need to be considered. Adjusting the carbon budget involves quantifying carbon fixation, yet in addition of its getaway through deterioration [1, 2]. The bound minerals that are additionally liberated as the wood decays are fundamental supplements the nutrient cycle of the forest [3-5]. Therefore, the species responsible for the breakdown of wood are an essential factor of the total forest biota.
Decay fungi of living trees can be classified and named utilizing various techniques. Accurate identification provides proof gives significant data about the effect of decay on the tree, mode of action and significance to hazard investigation. The presence of any fruiting body on a tree necessitates that the tree be examined all the more intently for decay [6].
Throughout forest ecosystems, Wood-decay fungi have a significant role in breaking down coarse woody debris (CWD) resulting from trees and other woody plants [7]. The Great Smoky Mountains National Park encompasses a total area of 2080 km2 in western North Carolina and eastern Tennessee between 35° 28’ and 35° 28’ N latitude. Elevations in the park range from approximately 270 to 2000 m above sea level. In late November and early December of 2016, wildfires occurred over portions of the Great Smoky Mountains National Park. More than 4,000 ha were affected by the wildfires. Some areas were subjected to intense fires that consumed most of the vegetation, whereas the fires in other areas were less intense. The objective of the project reported herein was to access the impact of fire on the assemblages of wood-decay fungi in two study areas, the first subjected to a relatively high intensity burn and the second subjected to a relatively low intensity burn.
Methods![]()
Study area: The study was conducted during the period of November to December 2019. The relatively higher intensity burn study area was along the Baskins Creek Trail (35° 40’ 45” N, 83° 28’ 38” W). A second growth mixed oak-hardwood forest was present prior to the burn. This study area is located on a slope with a west-southwest aspect at an elevation of approximately 865 m. The relatively lower intensity burn study area was the Twin Creeks All Taxa Biodiversity Inventory site (35° 41’ 11” N, 83° 29’ 60” W). A second growth mixed hardwood cove forest with a great laurel (Rhododendron maximum L.) understory was present prior to the burn. This study area is located on a slope with a west-northwest aspect at an elevation of approximately 595 m.
Collection of samples
Collecting trips were made to both study areas in late November 2019. An opportunistic method of search, as Cannon & Sutton described [8] was used to locate the fruiting bodies of wood-decay fungi. Once located, fruiting bodies in the field were photographed and then removed from the substrate upon which they occurred using a knife. These fruiting bodies were loosely wrapped in aluminum foil or placed in a plastic collecting box with several compartments and then returned the laboratory. In addition, a series of samples of CWD was collected from each study area. These were divided into two portions and placed in plastic chambers measuring (each 30 cm x 12 cm x 6 cm) in the manner described by Alshammari et al. [9]. A small amount of water was added to each chamber (Figs. 1A and B). These samples were observed over a period six months and watered when necessary to keep them moist. The fruiting bodies were photographed, removed from the CWD and then placed in one compartment of a plastic collecting box with numerous compartments.
All specimens were dried in the laboratory on a food dehydrator at a temperature between 42 °C and 50 °C, put in plastic bags, then labeled with a special collection number in the herbarium at the University of Arkansas (UARK).
Sampling and morphological observations
To determine morphological characteristics of the specimens, an AmScope stereomicroscope (10, 11) was used. Based upon these features (e.g., fruiting body size, shape, color and the presence or absence of such structures as a distinct cap or stipe) a tentative identification was reached by using sources of information such as [11-16].
Extraction of DNA, PCR and Sequencing
DNA was extracted from one or more representative specimens for each different taxon tentatively identified based on morphological characteristics of the fruiting body. The DNA extraction was performed using a genomic purification kit from Wizard ® (Promega Corporation, Madison, WI). Genomic DNA amplification was achieved using universal primers ITS4 (5’-TCCTCCGCTTATTGATATGC-3’) and ITS1 (5’-TCCGTAGGTGAACCTGCGG-3’) (17). In a 5 min thermocycler optimized for initial denaturation at 95 °C, PCR amplifications were carried out, 35 denaturation cycles followed at 95 °C for 45 sec, annealing at 50 °C for 45 sec, And extension for 1.5 min at 72 °C, and final extension for 10 min at 72 °C. The length of amplified products has been verified by 1% agarose gel electrophoresis gel using 0.5 TAE buffer, SYBR safe staining dye and 1 kb DNA ladder (New England Biolabs, Ipswich, MI). Amplicons for Sanger sequencing were sent to GeneWiz (South Plainfield, NJ). The sequences obtained from the latter company were cleaned up and then classified through searches of nucleotide blast against the NCBI database.
Results![]()
Sequences were considered as identified to the level of species at 95% sequence similarity. When there was a lower % sequence identity match for a particular sequence, it was considered as identified only to the level of genus. A 95% of other studies used sequence similitude, although there no single cutoff value has been universally established for species identification across the kingdom Fungi. As described above, these taxa were taxonomically identified through both methods from morphological characters followed by genetic identification using the ITS sequence data.
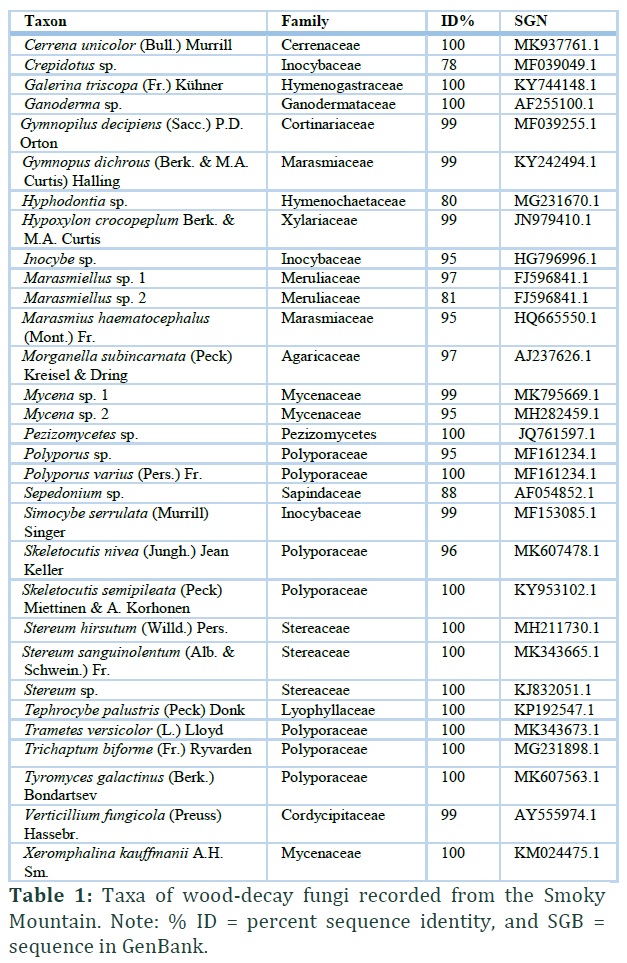
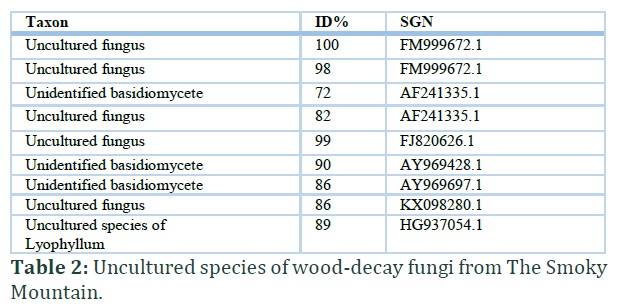
Forty-eight specimens of macro- fungi were sequenced. Out of the 48, sequences from eight specimens were of poor quality and thus not included in the analysis. Out of 40 good quality sequences, a total of 31 different taxa (Table 1) were identified, along with nine sequences that have been reported previously as uncultured species (Table 2).
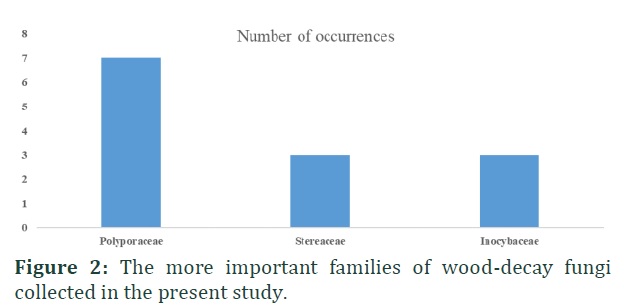
At least 18 different families were identified, with representatives of the Polyporaceae, Stereaceae and Inocybaceae the most common (Fig. 2 and 3). Also, 25 different genera were identified (Fig. 2 and 3). While the emphasis of this study was focused to wood-decay fungi, some of the taxa identified from the specimens associated with decaying wood have different ecological roles not related to wood decomposition.
In the more intensively burned study area, only a single species (Skeletocutis nivea) was collected from CWD. All of the other species reported herein were collected from CWD in the less intensively burned study area. Moreover, only a few of the pieces of CWD placed in incubation chambers produced any evidence of wood-decay fungi, and the latter was in the form of white mycelia that never yielded any fruiting bodies.
Figures & Tables


Discussion![]()
Few studies have examined the effects of burning on wood-decay fungi, and most of those of which we are aware [19] have considered prescribing burning and not wildfires. Soil microbiota, which is the primary agent responsible for organic matter deterioration and is associated with the macronutrient cycles [20, 21], is adversely influenced by fire. The significance of bacteria and fungi in in normal soil environments is all around perceived and they assume a significant role in the recovery of upset biological systems [22], adding to microbial biomass, the development of stable aggregates [23] and the degradation of headstrong substances [24].
Based on studies [25] of the effects of fire on the microbiology of soil, an intense fire accompanied by high temperatures can result in complete sterilization. There is no reason to suggest that the same would not be the case for CWD. Even at lower temperatures, mycelia already present in the wood would be reduced in extent even if they were not killed outright. The data obtained in the present study strongly suggest that small pieces of charred CWD do not provide an appropriate substrate for most species of wood-decay fungi but larger pieces (present at Twin Creeks but mostly absent at Baskins Creek Trail) can support the growth and development of at least some species of these fungi.
In conclusion, the data obtained in the present study, although limited in extent, do suggest that wildfires have a dramatic effect on the assemblages of wood-decay fungi associated with forests in the Great Smoky Mountains National Park. The information announced in this help the idea that prescribed burning has a recognizable impact upon wood-rot fungi, bringing about an appreciable decrease in the quantities of both fruiting bodies and taxa. Furthermore, the one of a kind part of the present effort was the utilization of incubation chambers to describe the taxa of fungi related with coarse woody debris. Clearly, there is a need for more comprehensive investigations of this aspect of forest ecology.
Author Contributions
Dr. Nawaf Alshammari, visited the location, samples collection, Morphology work, Molecular work, and wrote manuscript. Dr.Sami Hannachi, designed tables and figures, discussed results. Prof. Abdel Moneim Elhadi Sulieman, rewrote, edited, added some information in the introduction, and discussed the results. And prof. Stephenson, supervision all this work, facilitated forests work, reviewed the paper and discussed the work, provided all materials work and payment for DNA Sequencing.
The authors declare that there is no conflict of interest regarding the publication of this paper.
Acknowledgment
The research reported herein was supported in part by the Slime Mold Project at the University of Arkansas and a grant from Discover Life in America. University of Hail was enhanced and contributed in financial support.
References![]()
- Beets PN., Hood IA, Kimberley MO, Oliver GR, Pearce SH, Gardner JF. Coarse woody debris decay rates for seven indigenous tree species in the central North Island of New Zealand. Forest Ecology and Management , (2008); 256: 548–557.
- Mason NW, Bellingham PJ. Carswell FE, Peltzer DA, Holdaway RJ, Allen RB. Wood decay resistance moderates the effects of tree mortality on carbon storage in the indigenous forests of New Zealand. Forest Ecology and Management , (2013); 305: 177–188.
- Clinton PW, Buchanan PK, Allen RB. Nutrient composition of epigeous fungal sporocarps growing on different substrates in a New Zealand mountain beech forest. New Zealand journal of botany, (1999); 37:149–153.
- Clinton PW, Buchanan PK, Wilkie JP, Smaill SJ, Kimberley MO. Decomposition of Nothofagus wood in vitro and nutrient mobilization by fungi. Candian Journal of Forest Research, (2009); 39 (11): 2193–2202.
- Buchanan PK, Clinton PW, Allen RB. Dead wood in the forest — alive and dynamic! Zealand journal of botany, (2001); 45(4): 40–42.
- Seweta S, Ravidra K, Vinit P. Wood Decaying Fungi.. (2013); 3-5. LAP Lambert Academic Publishing.
- David L, Marco P. Wood-decaying fungi in the forest: conservation needs and management options. European Journal of Forest Research, (2008); 127:1–22
- Cannon P, Sutton B. Microfungi on wood and plant debris. In: Mueller G, Bills G, Foster M, (eds.) Biodiversity of Fungi: Inventory and Monitoring Methods. (2004); 217–239. Burlington, Massachusetts, Elsevier Academic Press.
- Alshammari, N. I. (2019). The Effect of Prescribed Burning on Wood-Decay Fungi in the Forests of Northwest Arkansas. Theses and Dissertations Retrieved from https://scholarworks.uark.edu/etd/3352 .
- Gilbertson RL, Ryvarden L. 1986 – North American Polypores, .Fungiflora, (1986); Volume I. Oslo, Norway.
- Sotome K, Akagi Y, Lee SS, Ishikawa NK, Hattori T. Taxonomic study of Favolus and Neofavolus gen. nov. segregated from Polyporus (Basidiomycota, Polyporales). Fungal Diversity, (2013); 58 (1) , 245–266.
- Gilbertson RL, Ryvarden L.– North American Polypores,. Fungiflora, (1987); Volume II Oslo, Norway.
- Bessette AE, Bessette AR, Fischer DW. Mushrooms of Northeastern. North America. Syracuse, (1997); University Press, Syracuse, New York.
- Barron G. Mushrooms of Northeast North America. (1999); Lone Pine Publishing, Vancouver, British Columbia.
- Binion DE, Stephenson SL, Roody WC, Burdsall HH et al. Macrofungi Associated with Oaks of Eastern North America. (2008); West Virginia University Press, Morgantown, West Virginia.
- Elliott TF, Stephenson SL. Mushrooms of the Southeast. (2018); Timber Press, Portland, Oregon.
- Toju H, Tanabe AS, Yamamoto S, Sato H. High-coverage ITS primers for the DNA-based identification of ascomycetes and basidiomycetes in environmental samples. PloS One, (2012); 7(7), e40863.
- Mousa MA., Sulieman AE, Veettil VN, Snoussi M, Antagonistic, biofilm-forming rhizospheric Pseudomonas spp. isolated from Hail province. Advancements in Life Sciences | www.als-journal.com | May 2020 | Volume 7 | Issue 3 ( www.ncbi.nlm,nih.gov).
- Alshammari N, Stephenson S. A preliminary study of wood-decay fungi in forests of northwest Arkansas. Current Research in Environmental & Applied Mycology, (2018); 8 (5): 11.
- Hernández T, García C, Reinhardt I. Short-term effect of wildfire on the chemical, biochemical and microbiological properties of Mediterranean pine forest soils. Biology and Fertility of Soils, (1997); 25 (1): 109-116.
- Acea MJ. Carballas, T. Microbial fluctuations after soil heating and organic amendment. Bioresource Technology, (1999); 67 (1): 65-71.
- Forster SM. The role of microorganisms in aggregate formation and soil stabilization: Types of aggregation. Arid Soil Research and Rehabilitation, (1999); 4 (2): 85-98.
- Dinel H, Levesque M, Jambu P, Righi D. Microbial and longchain aliphatics in the formation of stable aggregates. Soil Science Society of America Journal, (1992); 56 (1): 1250- 125.
- Prieto-Fernández A, Acea MJ, Carballas T. Soil microbial and extractable C and N after wildfire. Biology and Fertility of Soils, (1998); 27: 132-142.
- Mataix-Solera JC. Guerreno F, Garcia-Orenes G, Barcenas-Moreno, Torres MP. Forest Fire Effects on Soil Microbiology. Chapter in Fire Effects on Soils and Restoration Strategies, (2009); Science Publishers, Inc., Boca Raton, Florida.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0