

Short Communication
Characterization of carboxymethyl cellulase produced by Trichoderma asperellum
Nguyen Hoang Loc1,*, Nguyen Hong Van1, Hoang Tan Quang2
Adv. life sci., vol. 8, no. 1, pp. 42-46, November 2020
*– Corresponding Author: Nguyen Hoang Loc (Email: nhloc@hueuni.edu.vn)
Authors' Affiliations
2. Institute of Biotechnology, Hue University, Thua Thien Hue – Vietnam
Note: This version is corrected after publishing.
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: The present study characterized extracellular carboxymethyl cellulase (CMCase), an enzyme catalyzes the hydrolysis of the cellulose, from Trichoderma asperellum PQ34 that was isolated from agricultural cultivation soil in Thua Thien Hue, Vietnam.
Methods: CMCase was produced by culturing T. asperellum PQ34 on Czapek-Dox medium supplemented with carboxymethyl cellulose (CMC) at 28ºC for 96 h at a shaking speed of 150 rpm. Enzyme activity was spectrophotometerically determined with CMC used as a substrate and the absorbance was measured at 540 nm. The molecular weight of CMCase was determined by zymogram based on sodium dodecyl sulfate-polyacrylamide gel electrophoresis with 0.2% CMC.
Results: CMCase achieved the highest activity after 4 days of culture with the optimal pH and temperature at 4 and 55°C. The enzyme maintains pH and thermal stability in the range of 4-5 and 10-40°C and the relative activity of both is more than 90%. The presence of metal ions at 5 mM such as Ca2+, Al3+, or Co2+ increased the enzyme activity up to 133%, 124%, and 217%, respectively. CMCase was stable with 1 mM ethylenediaminetetraacetic acid and 1 M urea, but inhibited by 5% sodium dodecyl sulfate (SDS) and 5% Triton-X100. Zymographic analysis showed that CMCase from T. asperellum PQ34 consists of two enzymes with a molecular weight of about 31 and 66 kDa.
Conclusion: CMCases from T. asperellum PQ34 exhibited characteristics of an acidic and thermostable enzyme. The interaction of Co2+ and CMCase resulted in a significant increase in enzyme activity, however they were strongly inhibited by 5% SDS.
Keywords: Carboxymethyl cellulase; Cellulase; Endo-β-1,4-glucanase, Trichoderma asperellum
Introduction![]()
Cellulose is an important structural component of the primary cell wall of green plants, many forms of algae and the oomycetes fungi [1]. Cellulose is made up of glucose monomers that are linked by β-1-4 glycosidic bonds. Carboxymethyl cellulase (CMCase), also known as endo-β-1,4-glucanase or endo-β-glucanase (EC. 3.2.1.4), is one of three enzymes of the cellulose hydrolytic enzyme system (cellulase) including exo-β-glucanase (EC 3.2.1.91) and β-glucosidase (EC 3.2.1.21) [2, 3]. Cellulases break down the β-1,4-glycosidic bonds of the cellulose molecule into monosaccharides such as β-glucose, or shorter polysaccharides and oligosaccharides [4]. Different source of cellulases usually affect to their substrate specificity, optimal pH and temperature [5]. Many fungal strains can secrete cellulase in larger amounts than bacterial strains and Trichoderma is known as the fungal genus with the most ability to secrete cellulase. Most commercial cellulases are produced by two genera Trichoderma and Aspergillus. This reflects the fact that mold is a powerful natural source of enzyme production and can be studied for applications on industrial scale [5, 6]. To date, there have been many reports on isolation and characterization of CMCase from Trichoderma species such as T. reesei [7-9], T. viride [5, 10] or T. harzianum [11]. In last few years, the study of cellulase production from T. asperellum has also become area of interest [12-17]. The present study conducted characterization of CMCase produced by T. asperellum PQ34 strain. The results of this study promise potential applications for large-scale in the future.
Methods![]()
Fungal strain
T. asperellum PQ34 strain (GenBank accession number: HM545081.1) was isolated from the surface soil layer (15-20 cm in depth) under different agricultural lands in Thua Thien Hue province, Vietnam [18, 19].
Production of extracellular CMCase
T. asperellum PQ34 strain was cultured on Czapek-Dox medium at 28ºC for 48 h to induce spore germination. Fungal spores were then harvested in sterile distilled water and diluted to a density of 107 spores/mL. 2 mL of spore solution were subcultured in 100 mL of Czapek-Dox medium and incubated at 28ºC for 72 h at a shaking speed of 180 rpm for mycelial production. After washing with MgCl2, mycelium biomass was cultured in the same medium, but glucose was changed by 1% (w/v) carboxymethyl cellulose (CMC) to induce CMCase production, at 28ºC for 96 h with a shaking speed of 150 rpm. The culture was centrifuged at 12,000 ×g for 10 min at 4ºC to discard biomass and obtain culture filtrate (crude enzyme). Crude enzyme was partially purified by 70% saturation ammonium sulfate at 4°C for 2 h and then centrifuged at 24,000 ×g for 10 min at 4ºC. The pellet was resuspended and dialyzed in 10 mM citrate buffer at an appropriate pH for further use.
The diffusion method in agar plate was used to evaluate extracellular CMCase production of T. asperellum PQ34 strain. 100 µL of the partial purified filtrates were loaded into pre-punched holes of equal sizes on the assay plate containing 1.5% agar and 0.5% CMC. The plate was then incubated at 50oC for 24 h and subsequently stained with 1% Lugol solution for detection of substrate hydrolysis [20].
CMCase assay
CMCase activity was assayed according to Iqbal et al [5] with CMC was used as substrate. Reaction mixture consist of 300 µL of 1% CMC (in 0.05 M sodium acetate buffer, pH 5) and 150 µL of partial purified enzyme was incubated at 50oC for 30 min. Reaction was then stopped by adding 600 µL of 3,5-dinitrosalicylic acid and boiling for 5 min to develop the colour. The absorbance was measured at a wavelength of 540 nm.
One unit of enzyme activity was defined as the amount of enzyme required to form 1 μmol of glucose per one minute under the assay conditions [9]. Total protein concentration was determined by the method of Bradford [21] with bovine serum albumin (Sigma-Aldrich) as the standard. The samples were read at a wavelength of 595 nm against the blank. The specific activity of CMCase was obtained by dividing units of the enzyme (U/mL) with the total protein content (mg/mL) in the sample.
Characterization of CMCase
Effects of pH and temperature on CMCase action were tested within the range of 3-8 and 10-90°C. 20 mM citrate buffer was used for pH 3-5, and 20 mM phosphate buffer for pH 6-8. Thermal and pH stabilities were determined by pre-incubating the enzyme at pH 3-8 and 10-90ºC for 30 min without substrate. The enzyme activity was measured at the optimal pH and temperature. CMCase were also incubated with 5 mM of metal ion (Al3+, Cu2+, Co2+, Ca2+, Zn2+, Mn2+, Fe3+, or Hg2+) or reagents (5% SDS, 5% Triton X-100, 1 mM ethylenediaminetetraacetic acid (EDTA), or 1 M urea) at room temperature and optimal pH for 30 min. Enzyme relative activity was then determined at the optimal temperature.
Zymography
The molecular weight of CMCase was determined by sodium dodecyl sulfate (SDS)-12% polyacrylamide gel electrophoresis (PAGE) with 0.2% CMC used as substrate. Electrophoresis was conducted at 4ºC for 3 h, the gel was then incubated in 20 mM citrate buffer at 50°C overnight with 1% (v/v) Triton X-100 to remove SDS. Finally, the gel was stained with Lugol’s solution.
Statistics
Each experiment was repeated 3 times. The data were expressed as the means ± standard errors, followed by one-way ANOVA analysis with Duncan’s test at p=0.05.
Results![]()
Cellulase activity
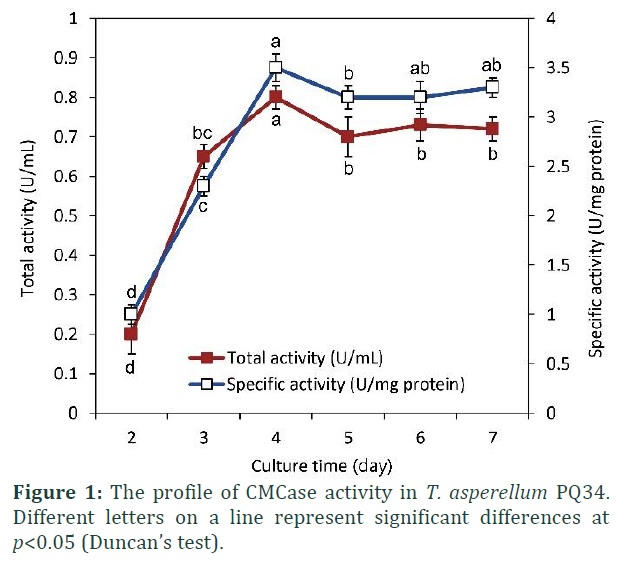
Extracellular cellulase activity of different T. asperellum strains including PQ34 was tested by agar plate with CMC substrate. The results indicated that the D–d value of T. asperellum PQ34 was the highest (about 1.4 cm) compared to that of other strains (CH2, SH16, and TN42) which were also isolated from the same place. Where, D is the diameter of the clear zone and d is the diameter of the pre-punched hole for enzyme loading (data not shown). After 4 days of culture, cellulase activity from PQ34 strain had peaked about 0.83 U/mL (equivalent to 3.62 U/mg protein) (Fig. 1). This strain was, therefore, used for characterizing CMCase.
Effect of temperature on CMCase activity
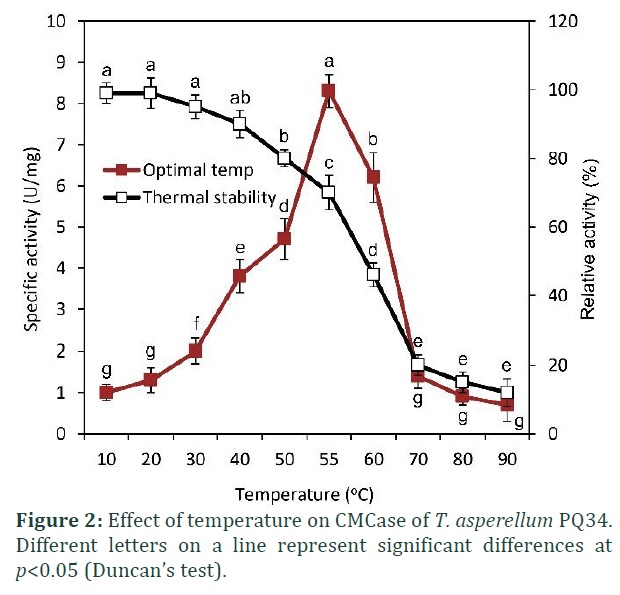
Data from Fig. 2 show that CMCase from T. asperellum PQ34 peaked at a relatively high temperature of 55°C with total activity of 1.7 U/mL (8.1 U/mg protein) and then drastically decreased at higher temperatures (60-90ºC). The activity of CMCase remained more than 90% when it was treated at temperatures below 40ºC. However, the enzyme activity decreased significantly at higher temperatures, especially it was only approximately 10% at temperatures from 70-90ºC (Fig. 2).
Effect of pH on CMCase activity
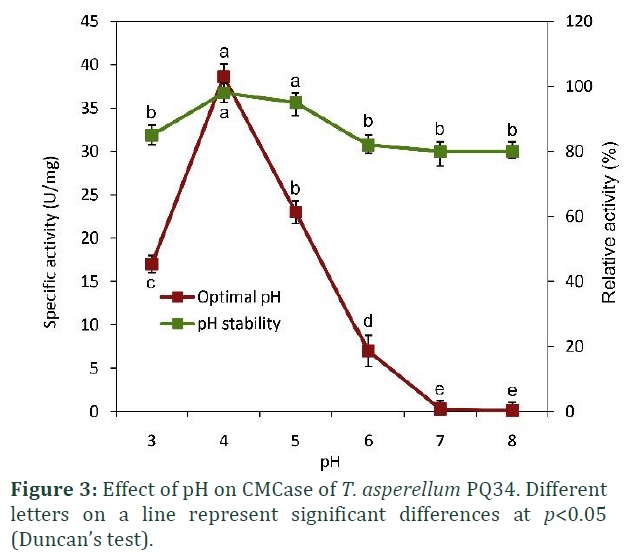
The present work on T. asperellum PQ34 strain has shown that CMCase has an optimal pH of 4 with a specific activity of approximately 40 U/mg, enzyme activity was completely lost at pH greater than 7. The pH stability of enzyme ranged from 4-5 with the relative activity of over 90%. Generally, the CMCase from T. asperellum PQ34 only works within a very narrow pH range in the acidic environment (Fig. 3).
Similar to CMCase from PQ34, the pH stability of CMCase from T. asperellum UC1 is also at 4 [17].
Effect of metal ions and reagents on CMCase activity
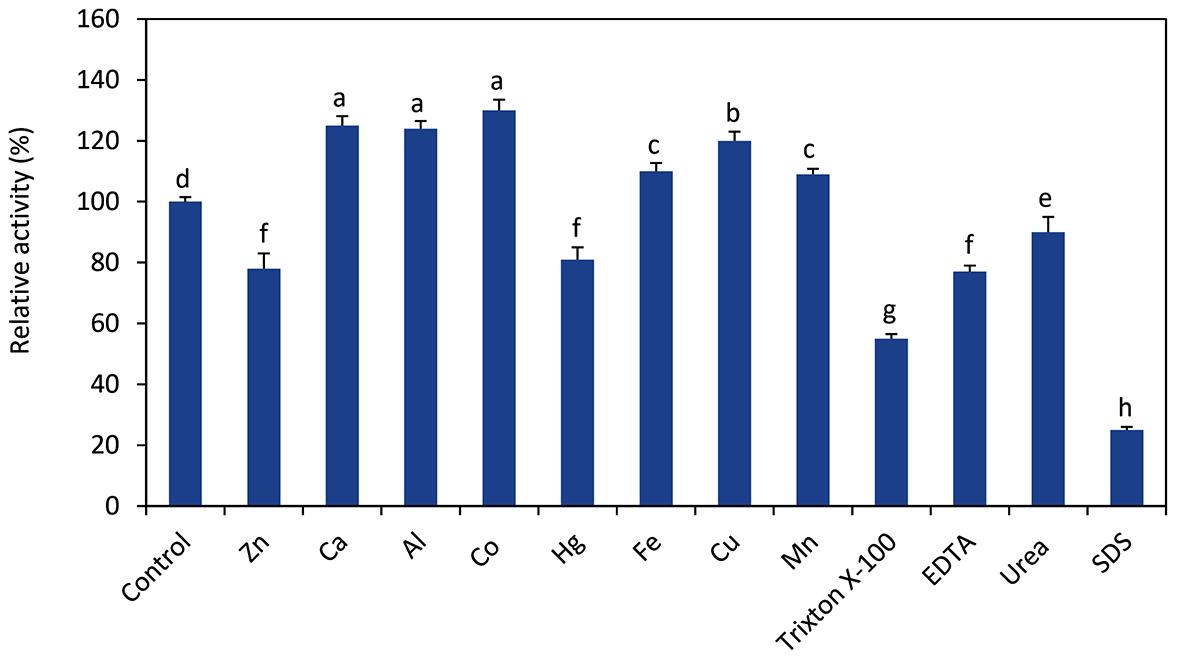
Investigation showed that some metal ions such as Zn2+ and Hg2+ slightly inhibited PQ34 CMCase, the relative activity was approximately 80%. While the other tested ions increased the enzyme activity such as Ca2+ (125%), Al3+ (124%) and Co2+ (128%). In the present work, all of the tested reagents inhibited enzyme activity, especially SDS (relative activity was about 25%) and Triton X-100 (relative activity was about 55%) (Fig. 4).
Zymogram
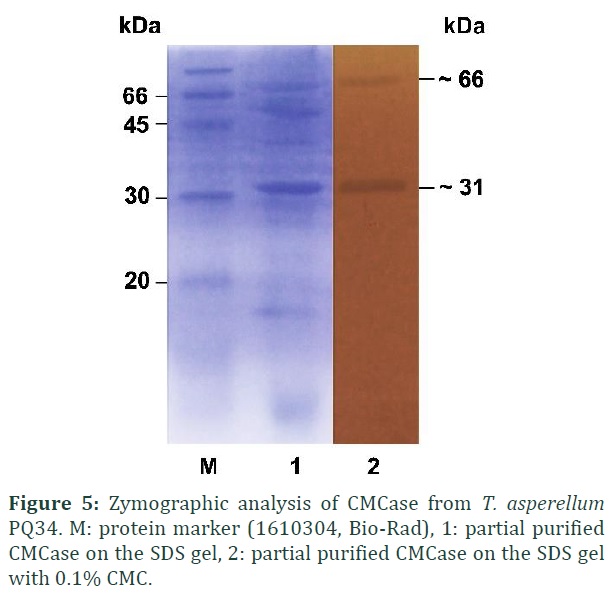
Zymographic analysis of CMCase by SDS-PAGE with CMC used as the substrate has detected enzymatic activity on the gel. Two clear zones were found on the gel which have the molecular weight of about 31 and 66 kDa (Fig. 5).
Up to now, there are no any reports related to the molecular weight of CMCases in T. asperellum. However, previous studies in Trichoderma species have shown that this enzyme has many types with different molecular weights. For example, cellulase III from T. viride has a molecular weight of 45 kDa [10], endoglucanase III from T. reesei QM 9414 is 48 kDa [7], CMCase from T. viride is 55 kDa [22], and cellulase from T. viride is 58 kDa [5].
Figures & Tables
Discussion![]()
Baig [23] obtained CMCase from T. lignorum with total activity of 0.25 U/mL on medium supplemented with CMC as carbon sources after 8 days of culture. Recently, solid state fermentations were performed for the production of cellulase and endoglucanase from T. asperellum, the enzyme activity reached maximum values of 5.6 and 4.1 U/g with paper used as substrate, respectively [16].
Some other studies in Trichoderma genus also showed the enzyme activity reached maximum values at the 55ºC likes CMCase from PQ34 strain. For example, endoglucanse III from T. reesei QM 9414 [7], cellulase from T. viride [5], or CMCase from T. reesei NRRL 3652 [9]. However, several other cellulases had maximum activity at lower temperatures such as 50ºC for cellulase III from T. viride [10, 22], 45°C for cellulase from Trichoderma sp. [24], 40-50°C for cellulase from T. reesei [25], 30-40°C for CMCase from T. harzianum [26, 27], or 30ºC for CMCase from T. reesei and T. harzianum [8, 11]. According to Guoweia et al [28] the optimal temperature for production of CMCase from T. reesei HY07 was 30°C. Endoglucanase III from T. reesei QM 9414 can withstand the activity at 65ºC for 30 minutes [7]. CMCase from T. asperellum UC1 retained 50% activitiy at 60°C [17]. Zhekova et al [9] obtained CMCase from T. reesei NRRL 3652 with thermal stability at 50°C within 160 minutes.
Trichoderma asperellumRCK2011 strains including wild-type and mutant can induce cellulases in the range of pH 4-10 [13]. Some CMCases only work in the acidic environment such as endoglucanase III from T. reesei QM 9414 at pH 4-5 [7], CMCase from T. reesei NRRL 3652 at pH 5 [9], cellulase III from T. viride at pH 4.5-5 [10], CMCase from T. viride at pH 5 [22] , cellulase from a mutation of T. reesei 2414 at pH 4.5 [25], even at pH 3 as CMCase from T. reesei [8]. Iqbal et al [5] and Nawaz et al [11] also found cellulases from T. viride and T. harzianum had higher optimal pH of 6.5 and 6, respectively. Li et al [29] showed CMCase from T. viride reached a maximum activity after 60 h of solid-state fermentation at 50ºC and pH 5. Attitalla and Salleh [26] found the optimum initial pH was 5-7 for CMCase from T. harzianum. Whereas cellulase from T. hazianum and Trichoderma sp. have the optimal pH at 4-6 and 6.5, respectively [24, 27].
The use of metal ions to enhance the cellulase activity has also been studied [30]. For example, Co2+ and Mn2+ enhance the activity of cellulase from T. viride [5], Ca2+ and Co2+ enhance CMCase activity from T. harzianum nearly two-fold [11]. Okada [10] found that cellulase III from T. viride was inhibited by Hg2+ but not affected by EDTA. While Iqbal et al [5] reported Hg2+, SDS and EDTA inhibited cellulase activity from T. viride. Nawaz et al [11] also showed that Hg2+ inhibited CMCase activity from T. harzianum.
CMCase of T. asperellum PQ34 strain has the optimum pH and temperature of 4 and 55°C, respectively. The pH and thermal stability of CMCase are in the range of 4-5 and 10-40°C with relative activities in both are more than 90%. The presence of metal ions at 5 mM concentration such as Ca2+, Al3+, or Co2+ increased the enzyme activity up to 133%, 124%, and 217%, respectively. Enzyme is stable with 1 mM EDTA and 1 M urea, but inactivated by 5% SDS and 5% Triton-X100. Zymographic analysis showed that CMCase from T. asperellum PQ34 strain consists of two enzymes which have the molecular weight of about 31 and 66 kDa.
Author Contributions
Loc NH: Designed study, processed experimental data and analyzed the results, wrote the manuscript.
Van NH and Quang HT: Conducted the research work.
The authors declare that there is no conflict of interest regarding the publication of this paper.
Acknowledgment
The authors would like to thank Hue University, Hue, Vietnam for their support for this study.
References![]()
- Cellulose. URL: https://en.wikipedia.org/wiki/Cellulose. Date accessed: 24 Oct. 2020.
- Kaur J, Chadha BS, Kumar BA, Saini HS. Purification and characterization of two endoglucanases from Melanocarpus sp. MTCC 3922. Bioresource Technology, (2007); 1: 74-81.
- Thongekkaew J, Ikeda H, Masaki K, Iefuji H. An acidic and thermostable carboxymethyl cellulase from the yeast Cryptococcus sp. S-2: purification, characterization and improvement of its recombinant enzyme production by high cell-density fermentation of Pichia pastoris. Protein Expression and Purification, (2008); 2: 140-146.
- Sani JK, Saini R, Tewari L. Lignocellulosic agriculture wastes as biomass feedstocks for second-generation bioethanol production: concepts and recent developments. 3 Biotech, (2015); 4: 337-353.
- Iqbal HMN, Ahmed I, Zia MA, Irfan M. Purification and characterization of the kinetic parameters of cellulase produced from wheat straw by Trichoderma viride under SSF and its detergent compatibility. Advances in Bioscience and Biotechnology, (2011); 3: 149-156.
- Jourdier E, Cohen C, Poughon L, Larroche C, Monot F, Chaabane FB. Cellulase activity mapping of Trichoderma reesei cultivated in sugar mixtures under fed-batch conditions. Biotechnology for Biofuels, (2013); 79.
- Macarrón R, Acebal C, Castiilón MP, Domínguez JM, Manta IDL. Mode of action of endoglucanase III from Trichoderma reesei. Biochemical Journal, (1993); Pt 3: 867-873.
- Shafaq A, Malana MA, Ikram N, Ghori MI, Butt KY, Ahmed S. Kinetic study of carboxymethylcellulase from Trichoderma reesei. Pakistan Journal of Life and Social Sciences, (2004); 2: 1-4.
- Zhekova B, Dobrev G, Dobreva V, Hadjikinova M. Characterization of enzyme with carboxymethyl cellulase activity mproduced by Trichoderma reesei NRRL 3652. Agricultural Science and Technology, (2012); 3: 311-314.
- Okada G. Enzymatic studies on a cellulase system of Trichoderma viride. The Journal of Biochemistry, (1976); 5: 913-922.
- Nawaz S, Malana MA, Ikram N, Hafeez S, Ghori MI, Jamil A. Kinetic study of carboxymethyl cellulase from Trichoderma harzianum. Pakistan Journal of Life and Social Sciences, (2006); 1-2: 15-19.
- Bech L, Busk PK, Lange L. Cell wall degrading enzymes in Trichoderma asperellum grown on wheat bran. Fungal Genomics and Biology, (2014); 1.
- Raghuwanshi S, Deswal D, Karp M, Kuhad RC. Bioprocessing of enhanced cellulase production from a mutant of Trichoderma asperellum RCK2011 and its application in hydrolysis of cellulose. Fuel, (2014); 183-189.
- Wang Q, Lin H, Shen Q, Fan X, Bai N, Zhao Y. Characterization of cellulase secretion and cre1-mediated carbon source repression in the potential lignocellulose-degrading strain Trichoderma asperellum T-1. PLoS One, (2015); 10:e0119237
- Inoue H, Kitao C, Yano S, Sawayama S. Production of β-xylosidase from Trichoderma asperellum KIF125 and its application in efficient hydrolysis of pretreated rice straw with fungal cellulase. World Journal of Microbiology and Biotechnology, (2016); 11:186.
- Zapata YM, Galviz-Quezada A, Osorio-Echeverri VM. Cellulases production on paper and sawdust using native Trichoderma asperellum. Universitas Scientiarum, (2018); 3: 419-436
- Ezeilo UR, Lee CT, Huyop F, Zakaria II, Wahab RA. Raw oil palm frond leaves as cost-effective substrate for cellulase and xylanase productions by Trichoderma asperellum UC1 under solid-state fermentation. Journal of Environmental Management, (2019); 206-217.
- Loc NH, Quang HT, Hung NB, Huy ND, Phuong TTB, Ha TTT. Trichoderma asperellum Chi42 genes encode chitinase. Mycobiology, (2011); 3: 182-186.
- Loc NH, Huy ND, Quang HT, Lan TT, Ha TTT. Characterisation and antifungal activity of extracellular chitinase from a biocontrol fungus, Trichoderma asperellum PQ34. Mycology: An International Journal on Fungal Biology, (2020); 1: 38-48.
- Calissendorff J, Falhammar H. Lugol’s solution and other iodide preparations: perspectives and research directions in Graves’ disease. Endocrine, (2017); 3: 467-473.
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry, (1976); 1-2: 248-254.
- Baig MMV. Cellulolytic enzymes of Trichoderma lignorum produced on banana agro-waste: Optimisation of culture medium and conditions. Journal of Scientific & Industrial Research, (2005); 64: 57-60
- Bhattacharya S, Das A, Patnaik A, Bokade P, Rajan SS. Submerged fermentation and characterization of carboxymethyl cellulase from a rhizospheric isolate of Trichoderma viride associated with Azadirachta indica. Journal of Scientific & Industrial Research, (2014); 73: 225-230.
- Gautam SP, Bundela PS, Pandey AK, Khan J, Awasthi MK, Sarsaiya S. Optimization for the product ion of cellulase enzyme from municipal solid waste residue by two novel cellulolytic fungi. Biotechnology Research International, (2011); Article ID 810425.
- Darabzadeh N, Hamidi-Esfahani Z, Hejazi P. Improvement of cellulase production and its characteristics by inducing mutation on Trichoderma reesei 2414 under solid state fermentation on rice by-products. Applied Food Biotechnology, (2018); 1: 11-18.
- Attitalla IH, Salleh B. Improvement of carboxymethyl cellulase and xylanase production by alginate immobilized Trichoderma harzianum. Biotechnology, (2010); 4: 529-532
- Pandey S, Shrivastava M, Shahid M, Kumar V, Singh A, Trivedi S, Srivastava YK. Trichoderma species cellulases produced by solid state fermentation. Journal of Data Mining in Genomics & Proteomics, (2015); 2: 1000170.
- Guowei S, Man H, Shikai W, He C. Effect of some factors on production of cellulase by Trichoderma reesei HY07. Procedia Environmental Sciences, (2011); 8: 357-361.
- Li XH, Yang HJ, Roy B, Park EY, Jiang LJ, Wang D, Miao YG. Enhanced cellulase production of the Trichoderma viride mutated by microwave and ultraviolet. Microbiological Research, (2010); 3: 190-198.
- Andreini C, Bertini I, Cavallaro G, Holliday GL, Thornton JM. Metal ions in biological catalysis: from enzyme databases to general principles. Journal of Biological Inorganic Chemistry, (2008); 8: 1205-1218.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0
![]()