

Review Article
Narrative Review: Use of Competent Stimulating Peptide in Gene Transfer Via Suicide Plasmid in Streptococcus pneumoniae
Atif Amin Baig1*, Nor Aina Shafiqah Binti Zulkiflee1, Muhammad Hassan1, Mohd Adzim Khalili Bin Rohin2, Mohd Khairi Bin Zahri Johari2, Ahmad Zubaidi Bin A. Latif1, Muhammad Umer Khan3, Nordin Bin Simbak1
Adv. life sci., vol. 8, no. 2, pp. 211-216, February 2021
*– Corresponding Authors: Atif Amin Baig (Email: atifamin@unisza.edu.my)
Authors' Affiliations
2. Faculty of Health Sciences, Universiti Sultan Zainal Abidin, Gong Badak Campus, Hafsah Block, 21300 Kuala Nerus, Terengganu Darul Iman – Malaysia
3. Faculty of Allied Health Sciences, The University of Lahore – Pakistan
Abstract![]()
Introduction
Methods
Discussion
Conclusion
References
Abstract
Natural competency for genetic transformation of Streptococcus pneumoniae causes the emergence of novel or non-vaccine preventable pneumococcal serotypes. This phenomenon has become a global concern as it can spread quickly in the population through inhalation and close contact. The colonisation of S. pneumoniae at the upper respiratory tract can either become commensal or pathogenic. Once the bacterium invades into the body system, it will secrete its toxin and virulence protein to facilitate the invasion. Besides, S. pneumoniae can undergo natural biological transformation via uptake of exogenous DNA by horizontal gene transfer for integration and recombination of the genome. S. pneumoniae natural transformation is aided by competence-stimulating peptide (CSP) that induces the competence of bacteria. Natural transformation cascade of S. pneumoniae via CSP is triggered in the presence of conserved 17-amino acids peptide which is regulated and encoded by comC, comD and comE operon, where comC is responsible in secreting precursor CSP. Nowadays, researchers transforming S. pneumoniae by inserting the mutated S. pneumoniae gene through a vector, suicide plasmid. Suicide plasmids such as pID701, pAUL-A and pVA891 can be transferred but cannot replicate in the bacteria. Homologous recombination process occurs once the mutated gene of suicide plasmid is integrated with wild-type S. pneumoniae. Previous studies had used the transformation of suicide plasmid into S. pneumoniae as it can integrate with host DNA at specific insert for gene transfer. But there is no evidence on the role of CSP in horizontal/gene replacement via suicide plasmid in Streptococcus pneumoniae. This narrative review's scope as per defined purpose statement is to relate and recommend the use of competent stimulating peptide in efficient horizontal gene transfer via suicide plasmids in Streptococcus pneumoniae.
Keywords: Streptococcus pneumoniae transformation; CSP-based transformation; Gene transfer via suicide plasmid; Genetic exchange; Competence-stimulating peptide
Introduction![]()
Streptococcus pneumoniae (S. pneumoniae) comprises an extensive capsule and underneath of capsule is a cell wall that comprises of polysaccharides, choline-binding protein, teichoic acid and sortase processed protein [1,2]. The variability in the capsular region is the result of serotypes differentiation and identification of S. pneumoniae [3]. S. pneumoniae is a gram-positive diplococcus that grows in chains [2] and grouped as α-hemolytic viridans streptococci [1]. Pneumococcal identification can be done through Gram staining, optochin susceptibility test, catalase test and followed by bile solubility test for further confirmation [4,5]. Based on phylogenetic reconstruction of 16S-rRNA-sequence, S. pneumoniae is one of the streptococci Mitis group that has different structure of cell wall carbohydrate as well as sequence of competence pheromone [6]. Several members of Mitis group naturally produce specialized competence-stimulating peptide (CSP) and its cognate receptors, which are responsible for genetic transformation at population level to accomplish goals which are impossible by individual cells [7]. Competence was an early example of bacterial response organized by scattered peptide signals, termed quorum-sensing (QS) and found in both gram-positive and gram-negative bacteria [8,9]. The virulence of bacterium is associated with genetic competence, biofilm production and acid tolerance which are controlled by CSP based QS system [10]. Quorum sensing in S. pneumoniae is caused by reserved 17-aminoacid peptide pheromone, which is the product of comC gene and later processed by ABC transporter (ComAB). Regulatory system depends on two components, histidine kinase receptor (ComD) and its associated response regulator (ComE), responsible to recognize the concentration of CSP in medium and initiate the response [1,2].
Among strains of S. pneumoniae, six well-defined CSPs have been recognized due to variation in their sequence, but majority of strains secrete either one of the two pheromones: CSP1 and CSP2 [10]. Each distinct CSP variant or group of strains recognized as a pherotype and pherotype genetically carrying one of two variants are not able to respond in the presence of other pheromones because of its receptor specificity, i.e. strain carrying comC1 (CSP1) is unable to respond to extracellular peptide (CSP2) product of comC2 [13]. Competent bacteria bind double-stranded DNA available in environment and transfer these fragments within cytoplasm in single-stranded form. A foreign DNA fragment can integrate within bacterial chromosome and change genotype and phenotype of the cell [12]. Isolates sharing similar antibiotic resistance have similar pherotypes, e.g. penicillin-resistant isolates have CSP-1. In pneumococcal population, pherotype vary independently of serotype and has clonal property. More divergence in genetic material have more probability to show different pherotype [3]. Competent cells can lyse the non-competent pneumococci, through fratricide mechanism. In pneumococci, fratricide plays a key role in lateral transformation by lysing non-competent cells and providing naked DNA in environment [14,15]. In natural population, genetic exchange between Intra pherotype prevails than inter pherotype [3]. This review study aims to discuss the comparative efficiency of gene transformation in the presence and absence of CSP.
Methods![]()
Literature Search strategy and selection criteria
Data mining concerned to the literature of the present study was sorted out by using key words including suicide plasmid in Streptococcus pneumoniae, homologous recombination in Streptococcus pneumoniae, Competence-stimulating peptide in Streptococcus pneumoniae, pneumococcal disease in google scholar and PubMed database. Articles retrieved under key words limitation were narrow down based on the study’s content. Where feasible, literature was limited to those articles which were recently issued in last decade, unless the essence was pivotal to flourish the article. We elected a total of 59 peer-reviewed articles, 55 original research articles and 4 review articles, and summarized into this conceptual and comprehensive review.
Discussion![]()
S. pneumoniae invades the body system via inhalation of small water droplet and colonize at the upper respiratory tract [1]. The bacterium can be either non-invasive or invasive [16]. Non-invasive bacteria reside at non-sterile parts inclusive of ears, otitis media, bronchitis and sinusitis. However, once be invasive and pathogenic, it can invade sterile parts including blood, cerebrospinal fluid, synovial fluids, pleural fluid and pericardial fluid causing invasive pneumococcal disease (IPD) [16,17].
Based on phylogenetic reconstruction of 16S-rRNA-sequence, S. pneumoniae is one of the streptococci Mitis group that has different structure of cell wall carbohydrate as well as sequence of competence pheromone [6]. S. pneumoniae had undergone many evolutionary lineages resulting in cluster separation with S. oralis and S. infantis [6]. Specifically, this bacterium consists of many serotypes under few serogroups [18]. The previous study stated the isolation and identification of 93 pneumococcal serotypes among 46 serogroups. However, only 10 serogroups were only related to pneumococcal disease [18].
Pneumococcal disease, public concern has been classified to be an airborne disease and to be spread by direct contact. Children and elderly are more prone to get infected by this bacterium, since, they are immunocompromised. In immunocompetent person, S. pneumoniae colonise asymptomatically at upper respiratory tract [17]. However, the inflammatory response is activated when the virulent genes of cell wall such as hyaluronidase [1], pneumococcal surface protein A [19], pneumococcal surface protein C [20] and α-enolase adhere to the epithelial cells of the upper respiratory tract for progression of disease [21]. Eventually, the inflammatory response becomes intense leading to the development of pneumococcal pathogenesis [1].
Antibiotics are used as a treatment of ailments to treat pneumococcal infection. In 1970s all pneumococcal isolates were susceptible towards antibiotics prescribed such as penicillin, clindamycin, rifampin, trimethoprim-sulfamethoxazole, macrolides, cephalosporin and vancomycin [22]. However, in 1990s most of pneumococcal clinical isolate in the United States showed resistance towards the antibiotics. S. pneumoniae had to go through genetic diversity by altering penicillin-binding protein receptor on its cell wall [6]. Due to inefficiency of treatment, another strategy had been done to improve public health status and prevention of S. pneumoniae is by implementing vaccination.
Vaccination is a preventive measure to counter pneumococcal disease among population. This prevention strategy was considered successful as the vaccination can target 75% of pneumococcal disease in children and after one year, the rate of infected children is reduced to more than half. However, in years later, the vaccination fails to overcome the disease due to emergence of non-vaccine type pneumococcal strains [18,23].
S. pneumoniae can evolve into different types of strains by genetic recombination. This highly recombinogenic bacterium will possess natural transformation mediated by horizontal gene transfer [24], resulting in high genetic diversity of S. pneumoniae that is difficult to be covered by monovalent vaccine [25]. Nowadays, scientists aim to construct new vaccines such as life attenuated vaccine that can yield wide coverage of S. pneumoniae serotypes.
S. pneumoniae natural transformation discovered by Griffth in 1928 proved to be a crucial finding in research area [26]. This transformable bacterium can take up naked extracellular DNA from the environment and integrating it with its own genome [26]. S. pneumoniae takes up double-stranded linear DNA by using different methods, including direct transformation of a DNA based product [27], direct transformation of plasmid DNA [28] and conjugation [29]. These modes of transfers can be with or without exogenous CSP. Natural transformation is one of quorum sensing system of S. pneumoniae that plays a role in communication and interaction of bacteria to manage and regulate gene expression as per environmental changes and signals [30].
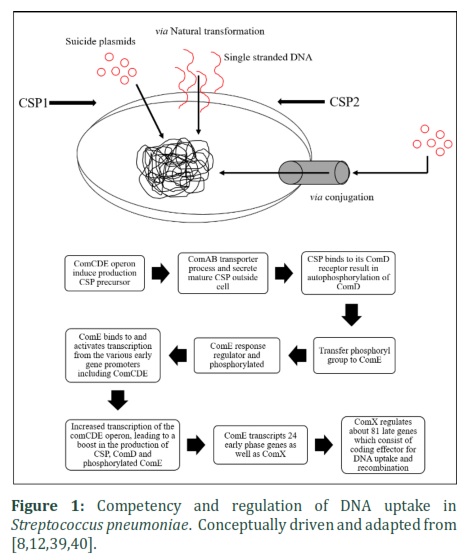
The transformation cascade is triggered by the presence of CSP which constitutes of 17-amino acids to induce the competency of bacteria [31]. Competence regulation of S. pneumoniae (figure 1) is regulated by its core protein encoded by comC, comD and comE operon, where comC is responsible in secreting precursor CSP [32,33]. The maturation and exportation of CSP outside cell are in charge by ATP-binding cassette (ABC) transporter, membrane-associated peptide permease, comA and accessory gene; comB [34-36]. CSP binding with histidine kinase receptor, comD autophosphorylation [37] which concurrently phosphorylates comE [38]. Transcriptional regulator, comE transcribed 24 early phase genes as well as comX and the activated ComX factor regulates about 81 late genes which consist of coding effector for DNA uptake and recombination [39]. Competence induction for genetic transformation of S. pneumoniae consists of pherotypes which are CSP1 and CSP2 [41]. These two variants have similar primary structures but they are highly specific to their receptors, comD1 and comD2. Therefore the specificity of phenotype depends on the type of pneumococcal strain [3]. In contrast with natural regulation of CSP, extracellular CSP is exogenously targets membrane-embedded histidine kinase receptor comD before initiates the downstream cascade events to achieve competence state [41]. However, when S. pneumoniae is induced to competent, fratricide factor encoded late genes; CbpD and CibAB are transcribed to upregulate LytA synthesis in the non-competent cell [26]. The stimulation of LytA, a murein hydrolase triggers pneumococcal autolysin that releases intracellular materials including chromosomal DNA [42]. The DNA uptake and release are in maximum level showing that the processes are simultaneously occurring [42,43]. The presence of exogenous CSP indicates that competent cell takes up extracellular DNA by killing neighbouring non-competent cell results in cell population reduction [43].
Process of DNA uptake occurs in two parts which are at outer membrane binding site and admission of DNA into the cell without being affected by DNase [44]. The binding of donor DNA to outer membrane of S. pneumoniae result in single-strand breaks before moving into the cell. Upon entry, one strand of DNA is degraded to oligonucleotides and another strand is introduced into the cell in single-stranded form [44]. The previous study stated that the incoming donor DNA strands move into
the cells in a 3’to 5’ direction [45]. Throughout the entry, the single-stranded DNA is coated with protein secreted by competent cells [46] and concurrently integrate with recipient chromosome complementary strand [47]. However, the transforming action cannot be performed due to lacking DNA processing factors such as comC encoded in production of competence stimulating peptide (CSP) and comD encoded for CSP receptor to phosphorylate comE [32,48].
Therefore, another approach had been used by cloning the gene of S. pneumoniae is into plasmid before introduced into the bacteria for gene recombination via transformation [29]. Homologous recombination will take place when the nucleotide sequences of two similar molecules of DNA are exchanged [49]. Throughout this process, Holliday junction takes place in which two
double-stranded of DNA molecules are separated into four strands and covalently linked to exchange part of genetic material and to make the genetic crossing over more specific, researchers are using suicide plasmid or shuttle plasmid [50,51]. Besides able to execute multiple mutations, the suicide plasmid can integrate with DNA by homologous recombination inside the chromosome without damages [51]. Suicide plasmid moves into the competent cell and integrates with target sequence by homologous recombination before eliminating itself through second cross-over event result in allelic exchange [51,52].
Transformation of suicide plasmid into S. pneumoniae through exogenous CSP had been reported [51] but the comparative transformant efficiency between with and without CSP induction have not been done yet.
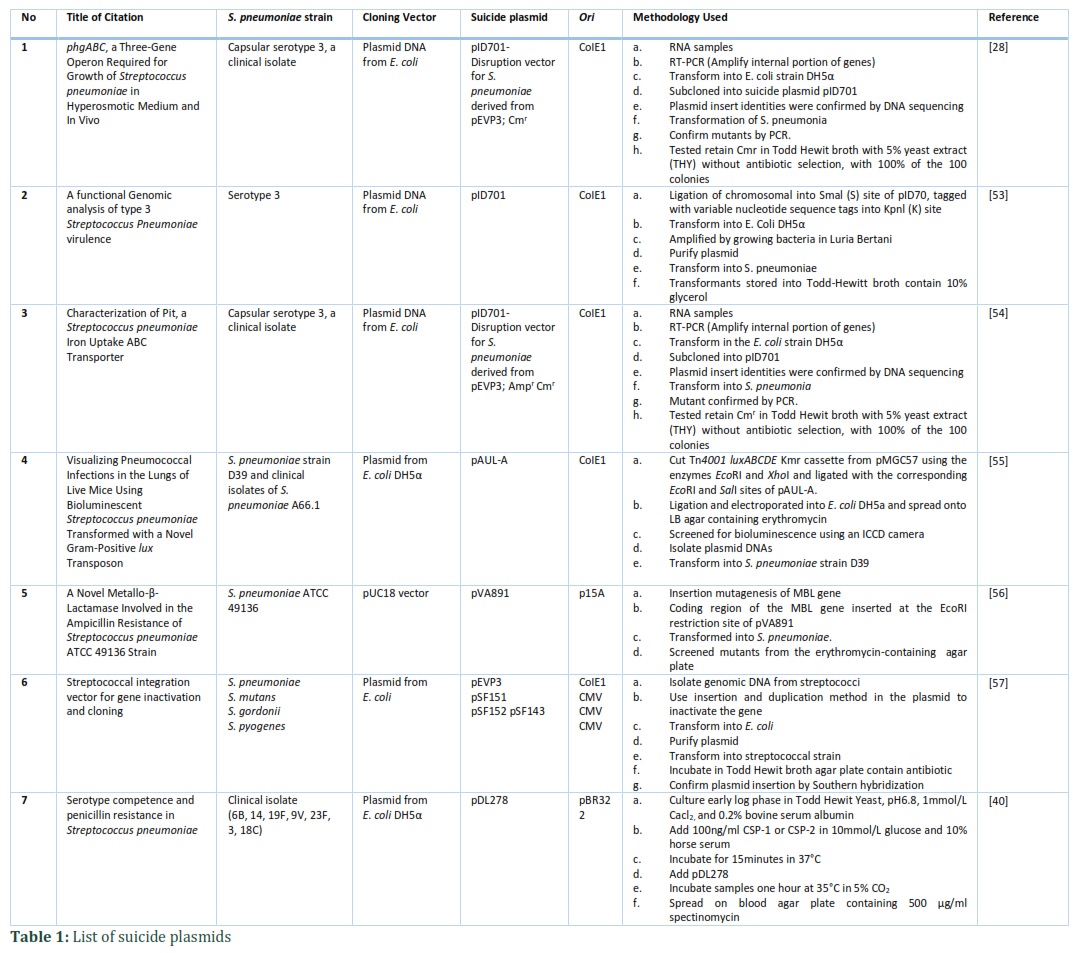
Therefore, the transformant efficiency of S. pneumoniae is low with positive selection of using competence-stimulating peptide due to autolysis as compare to the use of suicide plasmid for gene replacement in S. pneumonia [26,51]. Besides, there is a study that previously had used conjugation method by transferring plasmid pIP501 consist of DNAase resistant into S. pneumoniae through nitrocellulose filter [29]. The process is also known as filter mating that resembles conjugation mechanism. Along the process, pIP501 showed capability to mobilize a non-transmittable plasmid, pMV158 into clinical isolate group B streptococcus [58]. As stated (Table 1), some researchers used suicide plasmids that contain mutated gene of interest for replacement, to be transformed into S. pneumoniae such as pID701 [28,54], pAUL-A [55], pVA891 [56], pEVP3 [57], pSF151, pSF152, pSF143, pSF151 [57], pSF152 [57], pSF143 [57] and pDL278 [40].
- There is a need to study the comparative transformant efficiency between; with 2nd method. CSP indication in different serotypes of S. pneumonia.
- Transformation of S. pneumoniae by using suicide plasmid as a vector is an alternative to genetic recombination. Besides using the proper protocol, choosing suitable types of plasmid for S. pneumoniae transformation is necessary as it is a key for successful experiment.
- Transforming bacteria as mentioned in this study needs an alternative and more efficient method.
Furthermore, the mechanism of biological properties of S. pneumoniae competence membrane needs to be studied so that the suicide plasmid used for transformation is compatible with the bacteria used. It is undeniable that with the combination of comprehensive theory and protocol, genetic recombination of S. pneumoniae will be succeeded and be chosen as one of an alternative way to improve the prevention strategy against pneumococcal disease under the umbrella of CSP.
The research is supported by FRGS/1/2017/SKK06/UNISZA/02/1 under Kementerian Pendidikan Malaysia (KPM) and Universiti Sultan Zainal Abidin.
N.A.S., M.H.: Data mining and write manuscript; A.A.B., M.A.K., M.K.Z: Study design; A.Z.L., M.U.K., N.B.S, A.A.B: review manuscript and supervised
The authors declare that they have no conflict of interest.
References ![]()
- Bogaert D. de Groot R, Hermans PW. Streptococcus pneumoniae colonisation: the key to pneumococcal disease. The Lancet Infectious diseases, (2004); 4(3): 144-154.
- Dzaraly ND, Muthanna A, Mohd Desa MN, Taib NM, Masri SN, et al. Pilus islets and the clonal spread of piliated Streptococcus pneumoniae: A review. International Journal of Medical Microbiology, (2020); 310(7): 151449.
- Carrolo M, Pinto FR, Melo-Cristino J, Ramirez M. Pherotypes are driving genetic differentiation within Streptococcus pneumoniae. BMC microbiology, (2009); 9(1): 191.
- Organization WH (2003) Manual for the laboratory identification and antimicrobial susceptibility testing of bacterial pathogens of public health importance in the developing world: Haemophilus influenzae, Neisseria meningitidis, Streptococcus pneumoniae, Neisseria gonorrhoea, Salmonella serotype Typhi, Shigella, and Vibrio cholerae. World Health Organization.
- Perilla MJ. Manual for the laboratory identification and antimicrobial susceptibility testing of bacterial pathogens of public health importance in the developing world; Haemophilus influenzae, Neisseria meningitidis, Streptococcus pneumoniae, Neisseria gonorrhoeae, Salmonella serotype Typhi, Shigella, and Vibrio cholerae. (2003).
- Kilian M, Poulsen K, Blomqvist T, Håvarstein LS, Bek-Thomsen M, et al. Evolution of Streptococcus pneumoniae and its close commensal relatives. PloS one, (2008); 3(7): e2683.
- Håvarstein LS, Gaustad P, Nes IF, Morrison DA. Identification of the streptococcal competence‐pheromone receptor. Molecular microbiology, (1996); 21(4): 863-869.
- Mukherjee S, Bassler BL. Bacterial quorum sensing in complex and dynamically changing environments. Nature Reviews Microbiology, (2019); 17(6): 371-382.
- Waters CM, Bassler BL. Quorum sensing: cell-to-cell communication in bacteria. Annu Rev Cell Dev Biol, (2005); 21319-346.
- Allan E, Hussain HA, Crawford KR, Miah S, Ascott ZK, et al. Genetic variation in comC, the gene encoding competence-stimulating peptide (CSP) in Streptococcus mutans. FEMS microbiology letters, (2007); 268(1): 47-51.
- Claverys J-P, Havarstein LS. Extracellular-peptide control of competence for genetic transformation in Streptococcus pneumoniae. Front Biosci, (2002); 7(d1798-1814).
- Shanker E, Federle MJ. Quorum sensing regulation of competence and bacteriocins in Streptococcus pneumoniae and mutans. Genes, (2017); 8(1): 15.
- RAMIREZ M, MORRISON DA, TOMASZ A. Ubiquitous distribution of the competence related genes comA and comC among isolates of Streptococcus pneumoniae. Microbial Drug Resistance, (1997); 3(1): 39-52.
- Hong W, Khampang P, Erbe C, Kumar S, Taylor SR, et al. Nontypeable Haemophilus influenzae inhibits autolysis and fratricide of Streptococcus pneumoniae in vitro. Microbes and infection, (2014); 16(3): 203-213.
- Håvarstein LS, Martin B, Johnsborg O, Granadel C, Claverys JP. New insights into the pneumococcal fratricide: relationship to clumping and identification of a novel immunity factor. Molecular microbiology, (2006); 59(4): 1297-1037.
- Randle E, Ninis N, Inwald D. Invasive pneumococcal disease. Archives of Disease in Childhood-Education and Practice, (2011); 96(5): 183-190.
- Weiser JN, Ferreira DM, Paton JC. Streptococcus pneumoniae: transmission, colonization and invasion. Nature Reviews Microbiology, (2018); 16(6): 355-367.
- Tocheva AS, Jefferies JM, Christodoulides M, Faust SN, Clarke SC. Distribution of carried pneumococcal clones in UK children following the introduction of the 7-valent pneumococcal conjugate vaccine: a 3-year cross-sectional population based analysis. Vaccine, (2013); 31(31): 3187-3190.
- Amonov M, Simbak N, Wan Hassan WMR, Ismail S, A. Rahman NI, et al. Disruption of the cpsE and endA Genes Attenuates Streptococcus pneumoniae Virulence: Towards the Development of a Live Attenuated Vaccine Candidate. Vaccines, (2020); 8(2): 187.
- Quin LR, Onwubiko C, Moore QC, Mills MF, McDaniel LS, et al. Factor H binding to PspC of Streptococcus pneumoniae increases adherence to human cell lines in vitro and enhances invasion of mouse lungs in vivo. Infection and immunity, (2007); 75(8): 4082-4087.
- Hassan M, Baig AA, Attique SA, Abbas S, Khan F, et al. Molecular docking of alpha-enolase to elucidate the promising candidates against Streptococcus pneumoniae infection. DARU Journal of Pharmaceutical Sciences, (2021); 1-12.
- Cybulska J, Jeljaszewicz J, Lund E, Munksgaard A. Prevalence of types of Diplococcus pneumoniae and their susceptibility to 30 antibiotics. Chemotherapy, (1970); 15(5): 304-316.
- Brandileone M-CC, Almeida SC, Minamisava R, Andrade A-L. Distribution of invasive Streptococcus pneumoniae serotypes before and 5 years after the introduction of 10-valent pneumococcal conjugate vaccine in Brazil. Vaccine, (2018); 36(19): 2559-2566.
- Chaguza C, Cornick JE, Everett DB. Mechanisms and impact of genetic recombination in the evolution of Streptococcus pneumoniae. Computational and structural biotechnology journal, (2015); 13241-247.
- Khan MN, Pryharski K, Pichichero ME. Immunogenicity and protective efficacy of monovalent PCVs containing 22F and 33F polysaccharides in mouse models of colonization and co-infection. Vaccine, (2018); 36(38): 5701-5708.
- Johnsborg O, Håvarstein LS. Regulation of natural genetic transformation and acquisition of transforming DNA in Streptococcus pneumoniae. FEMS microbiology reviews, (2009); 33(3): 627-642.
- LefranGois J, Samrakandi MM, Sicard AM. Electrotransformation and natural transformation of Streptococcus pneumoniae: requirement of DNA processing for recombination. Microbiology, (1998); 144(11): 3061-3068.
- Brown JS, Gilliland SM, Basavanna S, Holden DW. phgABC, a three-gene operon required for growth of Streptococcus pneumoniae in hyperosmotic medium and in vivo. Infection and immunity, (2004); 72(8): 4579-4588.
- Heap JT, Ehsaan M, Cooksley CM, Ng Y-K, Cartman ST, et al. Integration of DNA into bacterial chromosomes from plasmids without a counter-selection marker. Nucleic acids research, (2012); 40(8): e59-e59.
- Zhu L, Lau GW. Therapeutic potential of the Streptococcus pneumoniae competence regulon. Expert Review of Anti-infective Therapy, (2013); 11(3): 227-229.
- Bergé M, Langen H, Claverys J-P, Martin B. Identification of a protein that inactivates the competence-stimulating peptide of Streptococcus pneumoniae. Journal of bacteriology, (2002); 184(2): 610-613.
- Håvarstein LS, Coomaraswamy G, Morrison DA. An unmodified heptadecapeptide pheromone induces competence for genetic transformation in Streptococcus pneumoniae. Proceedings of the National Academy of Sciences, (1995); 92(24): 11140-11144.
- Pestova E, Håvarstein L, Morrison D. Regulation of competence for genetic transformation in Streptococcus pneumoniae by an auto‐induced peptide pheromone and a two‐component regulatory system. Molecular microbiology, (1996); 21(4): 853-862.
- Hui FM, Morrison DA. Genetic transformation in Streptococcus pneumoniae: nucleotide sequence analysis shows comA, a gene required for competence induction, to be a member of the bacterial ATP-dependent transport protein family. Journal of bacteriology, (1991); 173(1): 372-381.
- Chandler MS, Morrison DA. Identification of two proteins encoded by com, a competence control locus of Streptococcus pneumoniae. Journal of bacteriology, (1988); 170(7): 3136-3141.
- Hui FM, Zhou L, Morrison DA. Competence for genetic transformation in Streptococcus pneumoniae: organization of a regulatory locus with homology to two lactococcin A secretion genes. Gene, (1995); 153(1): 25-31.
- Havarstein L. Molecular Microbiology, vol. 21,". Identification of the streptococcal competence-pheromone receptor, (1996); 863-869.
- Martin B, Soulet al, Mirouze N, Prudhomme M, Mortier‐Barrière I, et al. ComE/ComE∼ P interplay dictates activation or extinction status of pneumococcal X‐state (competence). Molecular microbiology, (2013); 87(2): 394-411.
- Peterson SN, Sung CK, Cline R, Desai BV, Snesrud EC, et al. Identification of competence pheromone responsive genes in Streptococcus pneumoniae by use of DNA microarrays. Molecular microbiology, (2004); 51(4): 1051-1070.
- Hsieh Y-C, Wang J-T, Lee W-S, Hsueh P-R, Shao P-L, et al. Serotype competence and penicillin resistance in Streptococcus pneumoniae. Emerging infectious diseases, (2006); 12(11): 1709.
- Johnsborg O, Kristiansen PE, Blomqvist T, Håvarstein LS. A hydrophobic patch in the competence-stimulating peptide, a pneumococcal competence pheromone, is essential for specificity and biological activity. Journal of bacteriology, (2006); 188(5): 1744-1749.
- Herta T, Bhattacharyya A, Bollensdorf C, Kabus C, García P, et al. DNA-release by Streptococcus pneumoniae autolysin LytA induced Krueppel-like factor 4 expression in macrophages. Scientific reports, (2018); 8(1): 1-14.
- Steinmoen H, Teigen A, Håvarstein LS. Competence-induced cells of Streptococcus pneumoniae lyse competence-deficient cells of the same strain during cocultivation. Journal of bacteriology, (2003); 185(24): 7176-7183.
- Lacks S, Greenberg B, Neuberger M. Role of a deoxyribonuclease in the genetic transformation of Diplococcus pneumoniae. Proceedings of the National Academy of Sciences, (1974); 71(6): 2305-2309.
- Méjean V, Claverys J-P. Polarity of DNA entry in transformation of Streptococcus pneumoniae. Molecular and General Genetics MGG, (1988); 213(2-3): 444-448.
- Morrison D. Transformation in pneumococcus: protein content of eclipse complex. Journal of bacteriology, (1978); 136(2): 548-557.
- Fox MS. Fate of Transforming Deoxyribonucleate following Fixation by Transformable Bacteria: II. Nature, (1960); 187(4742): 1004-1006.
- Koirala B, Phillips NR, Tal-Gan Y. Unveiling the importance of amide protons in CSP: ComD interactions in Streptococcus pneumoniae. ACS Medicinal Chemistry Letters, (2019); 10(6): 880-886.
- Mostowy R, Croucher NJ, Hanage WP, Harris SR, Bentley S, et al. Heterogeneity in the frequency and characteristics of homologous recombination in pneumococcal evolution. PLoS Genet, (2014); 10(5): e1004300.
- Punatar RS, Martin MJ, Wyatt HD, Chan YW, West SC. Resolution of single and double Holliday junction recombination intermediates by GEN1. Proceedings of the National Academy of Sciences, (2017); 114(3): 443-450.
- Latif AZA, Rohin MAK, Yeo CC, Simbak N, Baig AA. Suicide Plasmids for Gene Replacement in Streptococcus pneumoniae; Requirement for Natural Transformation against Positive Selection Using Competent Stimulating Peptide. Asian Journal of Medicine and Biomedicine, (2018); 17-18.
- Philippe N, Alcaraz J-P, Coursange E, Geiselmann J, Schneider D. Improvement of pCVD442, a suicide plasmid for gene allele exchange in bacteria. Plasmid, (2004); 51(3): 246-255.
- Lau GW, Haataja S, Lonetto M, Kensit SE, Marra A, et al. A functional genomic analysis of type 3 Streptococcus pneumoniae virulence. Molecular microbiology, (2001); 40(3): 555-571.
- Brown JS, Gilliland SM, Ruiz-Albert J, Holden DW. Characterization of pit, a Streptococcus pneumoniae iron uptake ABC transporter. Infection and immunity, (2002); 70(8): 4389-4398.
- Francis KP, Yu J, Bellinger-Kawahara C, Joh D, Hawkinson MJ, et al. Visualizing Pneumococcal Infections in the Lungs of Live Mice Using Bioluminescent Streptococcus pneumoniaeTransformed with a Novel Gram-Positive luxTransposon. Infection and immunity, (2001); 69(5): 3350-3358.
- Chang C-Y, Lin H-J, Li B-R, Li Y-K. A Novel metallo-β-lactamase involved in the ampicillin resistance of Streptococcus pneumoniae ATCC 49136 strain. PloS one, (2016); 11(5): e0155905.
- Tao L. Streptococcal integration vectors for gene inactivation and cloning. Methods in cell science, (1998); 20(1-4): 59-64.
- Smith MD, Shoemaker NB, Burdett V, Guild WR. Transfer of plasmids by conjugation in Streptococcus pneumoniae. Plasmid, (1980); 3(1): 70-79.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0