

Full Length Research Article
Biosynthesis of poly-3-hydroxybutyrate by Rhodococcus pyridinivorans using unrelated carbon sources
Naima Khan*, Nazia Jamil
Adv. life sci., vol. 8, no. 2, pp. 128-132, February 2021
*– Corresponding Author: Naima Khan (Email: naima.phd.mmg@pu.edu.pk)
Authors' Affiliations
Editorial Expression of Concern
08 July 2025: Following publication of this paper, the internal audit (consequent to concerns on quality raised by Web of Science) notified Advancements in Life Sciences about errors in metadata. By this Editorial Expression of Concern, we alert the scientific community of this incidence as we do the required corrections.
Editorial Note
26 July 2025: You are viewing the latest version of this article having correction in article info (metadata) section. Expression of concern is hereby revoked.
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Polyhydroxyalkanoates are intracellular polymers comparable to synthetic plastic in their thermostable and elastomeric properties. PHAs are produced by bacteria under various nutrient – nitrogen – stress conditions.
Methods: Bacteria were isolated from hot water springs of Tatta Pani Kashmir and Karachi Mangrove forest Pakistan. Polyhydroxyalkanoate detection agar was used to isolates PHA producing bacteria and several carbon sources such as glucose, glycerol and palmitic acid were used for relative biomass and biopolymer productions. PHA was extracted by solvent extraction method using sodium hypochlorite and chloroform. Extracted polymer was characterized by Fourier transfer infrared spectroscopy (FTIR).
Results: Rhodococcus pyridinivorans NK19 (KY703220) produced up to 60% PHA with glucose, 40% with palmitic acid and 58% with glycerol as carbon sources. FTIR spectrum confirmed the polymer produced as poly -3, hydroxybutyrate. A peak at 1720 cm -1 of FTIR confirmed the presence of PHB monomers in the polymer extracted.
Conclusion: Rhodococcus pyridinivorans NK19 produced short chain length PHA interchangeably known as P3HB while utilizing unrelated carbon sources up to 60%.
Keywords: Glycerol; Fatty acids degrading bacteria; FTIR; Palmitic acid
Introduction![]()
Solid waste management has been a major problem since the industrial revolution in the 1950s. Incineration of plastics produces toxic waste, solid as well as gases, and results in incomplete degradation. Approximately 187 million tons of plastic waste are introduced into the environment annually [1]. Bioplastics can be used as alternatives of synthetic plastics. Polyhydroxyalkanoates (PHAs) are thermostable and elastomeric and degrade completely under favorable environmental conditions [2]. They are produced by bacteria as intracellular inclusion bodies. Bacteria accumulate PHAs and related compounds under various environmental stresses including limiting nutrients such as nitrogen thus making them better candidate to survive and withstand natural selection pressures. Polyhydroxyalkanoates are biodegradable, biocompatible polyesters with various industrial applications that almost parallel synthetic polyesters [3]. Most common PHAs are polyhydroxybutyrates (P3HB) and has wide number of applications due to its polystyrene like properties [4]. PHAs are cytoplasmic inclusions may range in size from 0.2 to 0.5 µm [5]. The molecular weight of PHB depends upon its source, growth situations and extraction method. Its molecular weight ranges from 50,000 to a million Da. Organically created, PHB is a semi crystalline isotactic stereo systematic polymer with 100% R configuration, which permits enhanced degradability [5].
Polyhydroxyalkanoates have wide range of applications in medical as well as industrial fields due to their living tissue compatibility, zero toxicity and biodegradability [6]. Their wide range of physical and chemical properties enable them to mimic extracellular matrix thus making in-vitro studies more reliable and natural. They have been used as sutures, adhesion barriers and valves to guide tissue repair and in regeneration devices such as cardiovascular patches, articular cartilage repair scaffolds, bone graft substitutes, and nerve guides [7,8].
Polyhydroxybutyrate is the PHA that has been most frequently studied and characterized to date. It is a natural and biodegradable aliphatic homopolymer that is composed of monomers of four carbon atoms. It is soluble in some organic solvents and insoluble in water. It has the chemical formula(C4H6O2)n, which corresponds to 55.81% carbon, 7.03% hydrogen, and 37.16% oxygen by weight. The industrial interest in the use of P(3HB) emerged during the1960s when its thermoplastic properties were first described. Its synthesis was first conducted in 1971 via the polymerization of a racemic mixture of b-butyrolactone using a catalyst system of triethyl aluminum and water, which resulted in a stereo regular polymer that is partially optically inactive. In additional studies, P(3HB) was biotechnologically obtained from bacteria with a low molar mass (1×104 Da) and crystallinity (29%). P(3HB) can be processed as a classic thermoplastic as it can resist wide temperature range (from 30 ºC to120 ºC). P(3HB) is non-toxic and its degradation produces 3-hydroxybutyric acid, a normal constituent of human blood. Thus, it can be used both in products that assimilate with human or animal tissue and for human consumption [9]. In this study we isolated several bacterial strains with ability to produce PHAs under salt stress. Ability to grow optimally in presence of salt makes bacterial isolates more desirable [10]. Salt tolerant bacteria minimize the growth of non-tolerant bacteria thus reducing the cost of production as there will be no need to sterilize the salt containing growth media. We used glycerol, glucose, palmitic acid and propionic acid as carbon sources for relative biomass and polymer production.
Methods![]()
Bacterial strain isolation and screening for salt tolerant PHA producers
Different bacterial strains were isolated from hot water springs of Kashmir, Pakistan and mangrove forest of Karachi, Pakistan. We collected rhizosphere soil of mangroves in sterile containers, similarly fine stones and water samples were also collected from hot water springs and surrounding area. Samples were serially diluted and bacterial colonies were obtained by spread plate method. Isolated bacterial strains were then subjected to screening based on salt tolerance and PHA production. PHA detection agar was supplemented with 0.5 M NaCl for screening purpose. Bacterial strains that were able to tolerate salt and produce PHA were identified using 16S ribosomal RNA gene. Deoxyribonucleic acid was extracted from bacteria and the conserved 16S rRNA gene was amplified and sequenced. Sequence homology was done using NCBI-BLAST to determine the similarity of isolated bacterial strain with already submitted sequences at the NCBI GenBank. For screening of PHA producing strains, Nile red and Nile blue A stains were used to stain the colonies. After 24 hours of incubation plates were checked for fluorescence. For further detection of PHA, Sudan Black B staining was done to visualize intracellular PHA granules.
Growth kinetics of isolated strain for pH, temperature and salt stress
Various pH (4, 6.8, 8, and 10), temperature (10°C, 25°C, 37°C and 45°C), nitrogen concentration (0.2, 0.5, 1 and 2%) and salt concentrations (0.1M, 0.5M, 1M, 2M, and 3 M) were used to optimize growth of isolated bacterial strains[11]. Bacterial growth was monitored at two-hour interval by measuring optical density of the broth culture at A660 nm. PHA production was also estimated at an interval of six hours. The experiment was conducted in shake flask culture and PHA was isolated using a modified method using [12] ultrasound assisted solvent extraction with chloroform and sodium hypochlorite.
Relative production od biomass and PHA using different carbon sources
Glucose – 2%, glycerol – 2% and palmitic acid – 0.5% was used as carbon sources for optimal PHA production. PHA detection agar (PDA) [13] was used to conduct shake flask experiment. All production experiments were conducted at optimal pH, temperature, and salt conditions. Growth and production of PHA was monitored for a period of 72 hours. After every two-hours, optical density of the culture was checked at A600 nm.
Characterization of extracted polymer
Fourier transfer infrared spectroscopy (FTIR) of the extracted polymer was done to check the presence of different PHA related functional groups. The samples were dissolved in chloroform and deposited on KBr disc and were scanned between 400 to 4000/cm wave number [14].
Results![]()
Bacterial strain identification and growth optimization
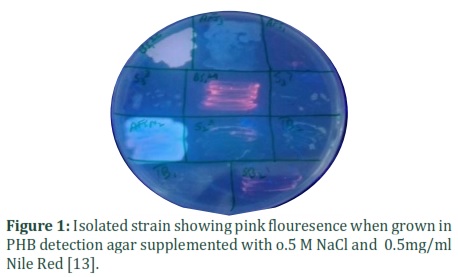
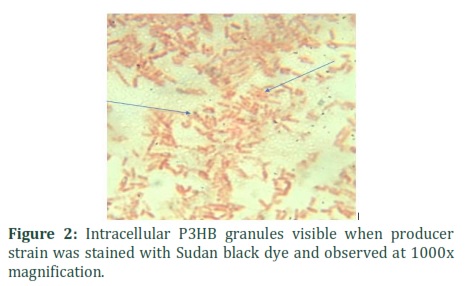
Bacterial colonies of Rhodococcus pyridinivorans (KY703220) stained with Nile blue and Nile red showed bright pink fluorescence when viewed under UV as shown in the figure 1. P3HB producing bacteria give fluorescence when grown in the presence of Nile red [15]. Sudan Black staining revealed purple-black granules inside rod shaped bacteria (figure 2). Sudan Black has high affinity for carbonyl functional group containing molecules that therefore, it is assumed that it must have attached with P3HB granules. Evolutionary history of selected strain, Rhodococcus pyridinivorans (KY703220) was detected using molecular evolutionary genetic analysis software (MEGA 7.2). Neighbor joining tree method gave optimal tree with the sum of branch length = 1.05268421 as shown in figure 3. The analysis involved nine nucleotide sequences selected from NCBI database having maximum similarity with the isolated strain, Rhodococcus pyridinivorans (KY703220).
Relative growth of Rhodococcus pyridinivorans under abiotic stresses
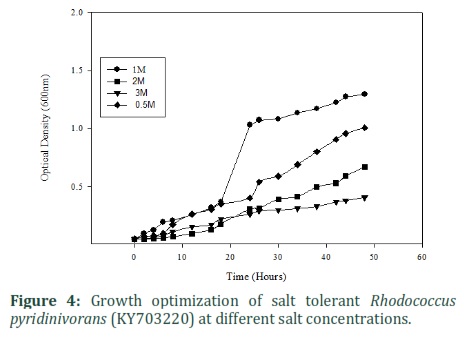
In this study we isolated fifty bacterial strains from extreme environments of Pakistan. After screening, several isolates were found to be PHA producers. Thus, our focus was to isolate salt tolerant PHA producing strains. Rhodococcus pyridinivorans (KY703220) was able to produce P3HB in as much as 3M salt media (figure 4). The isolated strain was able to grow and produce P3HB at different NaCl concentrations. However, at concentrations above 1M its growth rate reduced. Salt tolerant bacteria are an excellent candidate for the production of PHA as they might reduce sterility cost and will be able to grow in extreme conditions [16].
PHA production
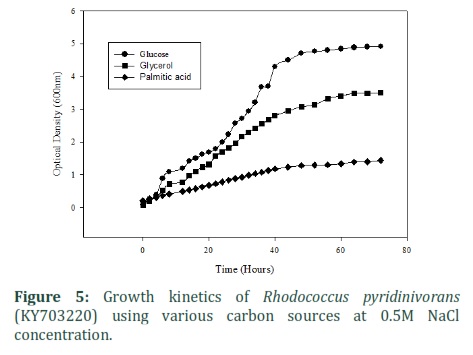
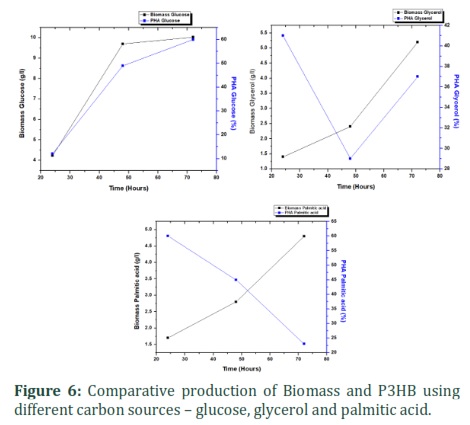
Glucose, Palmitic acid and Crude glycerol was used as carbon sources for the production of P3HB. Growth kinetics observed showed that strain was growing best with glucose giving optical density of 4.9, with glycerol OD recorded was 3.5 while with palmitic acid 1.435 after 72 hours of incubation at 37ºC and 0.5 M salt (⁓ 3%) as shown in figure 5. Rhodococcus pyridinivorans (KY703220) showed different trends for carbon sources as shown in the figure 6. When glucose was used as carbon source Rhodococcus pyridinivorans (KY703220) was able to produce 60 % PHB of dry cell weight. While with other carbon sources – glycerol and palmitic acid – the production was relatively low 40% and 58% respectively (figure 6). In this study we isolated bacterial strain that was not only able to produce P3HB using different types of carbon sources but can also tolerate high salt concentrations thus making it suitable for further large-scale studies. P3HB produced by Rhodococcus pyridinivorans (KY703220) can be used as it in various commercial application or modified by various metabolic as well as physical methods [17-19].
Characterization of extracted polymer
Fourier transfer infrared spectroscopy detected the presence of carbonyl functional group at 1720cm-1. This functional group is particularly associated with short chain length PHA.
Figures & Tables

Discussion![]()
Bio-plastic production has faced many setbacks due to the fact that its production is rather costly as compared to synthetic polymers. However, its importance cannot be denied and this problem can be fixed by discovering bacterial species that can survive under severe abiotic stress and utilize a number of unrelated carbon sources. Rhodococcus pyridinivorans (KY703220), although not a typical halophile, was able to tolerate NaCl concentrations as much as 1M (5.8%). Most other environmental isolates need vigorous aseptic conditions as other microbial contaminants can also compete with them under same growth conditions. Halophiles do not require such conditions as other mesophile are not able to grow under high salt stresses. This is an advantage that can be used in biotechnology to reduce production cost of various products as important as bioplastics [22]. Cost effective production of PHB can also be ensured by using cheap carbon sources. Carbon sources of organic and inorganic nature originated as waste from various capacities either domestic or commercial are being investigated to produce PHB [23]. Production of PHA, as much as 0.14g/l, has been previously reported by Rhodococcus opacus B4 using glucose, acetate and hexadecane and Rhodococcus equi produced 38% poly-3(hydroxybutyrate) using crude palm kernel oil [16,17]. In this study, isolated bacterial strain was able to accumulate 40% PHB using palmitic acid and 58 % with glycerol.
FTIR spectrum of the extracted PHA showed a strong peek at 1720.39 cm−1 which was reported as bond stretching vibrations of carbonyl (C O), this region is supposedly a polyhydroxybutyrate marker peek and is associated with amorphous region of the biopolymer. This absorption region also indicate that the polymer has maximum crystallinity another attribute of the short chain length PHA [24, 25]. Thus, the polymer extracted from Rhodococcus pyridinivorans (KY703220) is confirmed to be PHB. Due to high crystallinity of the scl-PHA it has a limited number of applications, however the polymer can be modified by increasing content of 3-hydroxyvelarate or 4-hydroxybutyrate and can be used in various fields including biomedical and cosmetics industry.
Author Contributions
Naima Khan conducted experimental work and constructed the initial draft of the manuscript.
Nazia Jamil designed experimental work as well as provided guidelines for the manuscript preparation.
All authors declare no conflicts of interest in this paper.
References![]()
- Ray S, Prajapati V, Patel K, Trivedi U. Optimization and characterization of PHA from isolate Pannonibacter phragmitetus ERC8 using glycerol waste. International journal of biological macromolecules, (2016); 86741-749.
- Teeka J, Imai T, Reungsang A, Cheng X, Yuliani E, et al. Characterization of polyhydroxyalkanoates (PHAs) biosynthesis by isolated Novosphingobium sp. THA_AIK7 using crude glycerol. Journal of industrial microbiology & biotechnology, (2012); 39(5): 749-758.
- Gumel AM, Annuar MSM, Heidelberg T. Biosynthesis and characterization of polyhydroxyalkanoates copolymers produced by Pseudomonas putida Bet001 isolated from palm oil mill effluent. PLoS One, (2012); 7(9): e45214.
- Sangkharak K, Prasertsan P. Screening and identification of polyhydroxyalkanoates producing bacteria and biochemical characterization of their possible application. The Journal of general and applied microbiology, (2012); 58(3): 173-182.
- Raza ZA, Abid S, Banat IM. Polyhydroxyalkanoates: Characteristics, production, recent developments and applications. International Biodeterioration & Biodegradation, (2018); 12645-56.
- Ali I, Jamil NJFib. Polyhydroxyalkanoates: current applications in the medical field. Frontiers in Biology, (2016); 11(1): 19-27.
- Cazzato RL, Garnon J, Ramamurthy N, Koch G, Tsoumakidou G, et al. Percutaneous image-guided cryoablation: current applications and results in the oncologic field. Medical oncology, (2016); 33(12): 140.
- Choy G, Khalilzadeh O, Michalski M, Do S, Samir AE, et al. Current applications and future impact of machine learning in radiology. Radiology, (2018); 288(2): 318-328.
- Alves MI, Macagnan KL, Rodrigues AA, de Assis DA, Torres MM, et al. Poly (3-hydroxybutyrate)-P (3HB): Review of production process technology. Industrial Biotechnology, (2017); 13(4): 192-208.
- Hong J-W, Song H-S, Moon Y-M, Hong Y-G, Bhatia SK, et al. Polyhydroxybutyrate production in halophilic marine bacteria Vibrio proteolyticus isolated from the Korean peninsula. Bioprocess and biosystems engineering, (2019); 42(4): 603-610.
- Abd El-malek F, Farag A, Omar S, Khairy HJIJoBM. Polyhydroxyalkanoates (PHA) from Halomonas pacifica ASL10 and Halomonas salifodiane ASL11 isolated from Mariout salt lakes. International Journal of Biological Macromolecules, (2020); 1611318-1328.
- Martínez-Herrera RE, Alemán-Huerta ME, Almaguer-Cantú V, Rosas-Flores W, Martínez-Gómez VJ, et al. Efficient recovery of thermostable polyhydroxybutyrate (PHB) by a rapid and solvent-free extraction protocol assisted by ultrasound. International Journal of Biological Macromolecules, (2020); 164: 771-782.
- Munir S, Iqbal S, Jamil NJJPAM. Polyhydroxyalkanoates (PHA) production using paper mill wastewater as carbon source in comparison with glucose. Journal of Pure and Applied Microbiology, (2015); 9453-460.
- Gomaa EZJBAoB, Technology. Production of polyhydroxyalkanoates (PHAs) by Bacillus subtilis and Escherichia coli grown on cane molasses fortified with ethanol. Brazilian Archives of Biology and Technology, (2014); 57(1): 145-154.
- Al-Kaddo KB, Sudesh K, Samian MRJMJoM. Screening of bacteria for PHA production using waste glycerol as carbon source and the ability of new strain to produce P (3HB-co-3HV) copolymer. Malaysian Journal of Microbiology, (2016); 12(3): 245-253.
- Zhang X, Lin Y, Chen GQJAB. Halophiles as chassis for bioproduction. Advanced Biology, (2018); 2(11): 1800088.
- Khan N, Jamil N (2019) Bacterial Conversion of Waste Products into Degradable Plastics: An Inexpensive Yet Eco-friendly Approach. Environmental Contaminants: Ecological Implications and Management: Springer. pp. 297-309.
- Altaee N, El-Hiti GA, Fahdil A, Sudesh K, Yousif EJAJfS, et al. Screening and evaluation of poly (3-hydroxybutyrate) with Rhodococcus equi using different carbon sources. Arabian Journal for Science and Engineering ,(2017); 42(6): 2371-2379.
- Castro AR, Rocha I, Alves MM, Pereira MAJAE. Rhodococcus opacus B4: a promising bacterium for production of biofuels and biobased chemicals. AMB Express, (2016); 6(1): 35.
- Biradar GG, Shivasharana C, Kaliwal BBJEJEB. Isolation and characterization of polyhydroxybutyrate (PHB) producing Bacillus species from agricultural soil. European Journal of Experimental Biology, (2015); 5(3): 58-65.
- Gao Z, Su T, Li P, Wang ZJB. Biodegradation of P (3HB-co-4HB) powder by Pseudomonas mendocina for preparation low-molecular-mass P (3HB-co-4HB). 3 Biotech, (2017); 7(5): 281.
- Kucera D, Pernicová I, Kovalcik A, Koller M, Mullerova L, et al. Characterization of the promising poly (3-hydroxybutyrate) producing halophilic bacterium Halomonas halophila. Bioresource Technology, (2018); 256552-556.
- Zhan J, Tian X, Zhu Y, Wang L, Ren LJAss. Oxygen plasma modified P (3HB-4HB) used as anticoagulant materials. ACS Applied Materials & Interfaces, (2013); 280564-571.
- Ribeiro PLL, Figueiredo TVB, Moura LE, Druzian JIJPfAT. Chemical modification of cellulose nanocrystals and their application in thermoplastic starch (TPS) and poly (3‐hydroxybutyrate) (P3HB) nanocomposites. Polymers for Advanced Technologies, (2019); 30(3): 573-583.
- Vigneswari S, Murugaiyah V, Kaur G, Khalil HA, Amirul AJBM. Corrigendum: Biomacromolecule immobilization: grafting offish-scale collagen peptides onto qaminolyzed P (3HB-co-4HB) scaffolds as potential wound dressing (2016Biomed. Mater. 11 055009). (2018); 13069502.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0