

Full Length Research Article
A study of PTF1 interaction with phosphorus stress inducing genes and its influence on root architecture of transgenic Arabidopsis
Aqsa Akhtar, Muhammad Irfan*, Asma Maqbool, Kauser Abdulla Malik
Adv. life sci., vol. 9, no. 1, pp. 60-66, May 2022
*– Corresponding Author: Muhammad Irfan (Email: ranahaiderali94@gmail.com)
Authors' Affiliations
Publisher's Note: This article was published as online-first (abstract only) on 25/07/2021 at http://www.als-journal.com/proonlinefirst-1211/.
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Phosphorus; an essential macronutrient needed by the plant for its robust growth is inaccessible to the plant as required. Hence, a need arises to develop smart crops capable of utilizing maximum phosphorus from soil. PTF1(Inorganic Phosphorus Starvation Induced Transcription Factor 1) is overexpressed during phosphorus stress and regulates number of genes to combat this abiotic stress.
Methods: The current study is the first ever reported case of transforming Arabidopsis thaliana with plant expression binary vector pSB219 harboring PTF1 via floral dip method and analyzing phosphorus stress induced genes interaction, through yeast-one-hybrid. Yeast-one-hybrid analysis was performed on four selected genes namely LPR1 (Low Phosphate Response), PDR2 (Phosphate Deficiency Response 2), PHT1;2 (Phosphate Transporter) and RNS1 (Ribonuclease). The positive transformed lines were expression analyzed for PTF1 by real time PCR and further studied for their root morphology.
Results: The results clearly showed direct interaction of LPR1 with PTF1 while other genes, although being overexpressed, were indirectly regulated. Transformation efficiency of 1% was achieved and a maximum 2.5-fold increase in PTF1 expression was observed. Root morphological studies exhibited significantly enhanced root hair and lateral surface area when grown in phosphorus deficient MS medium.
Conclusion: The results of the current study may pave path for improved comprehension of gene interactions and root architecture modifications under phosphorus limiting conditions.
Keywords: Phosphorus; Arabidopsis; Yeast one hybrid; Roots
Introduction![]()
Phosphorus; one of the essential macronutrients required by the plant plays a crucial role in various diverse metabolic, energy and signal transduction processes, thus, being imperative for robust plant growth [1-3]. However, its inaccessibility mainly due to plant’s inability of maximum absorption makes Pi (Inorganic Phosphorus) a limiting factor in plant growth resulting in stunted growth and limited crop yield [4,5]. Thus, developing smart crops, able to utilize maximum phosphorus seems an apt and eco-friendly solution. Numerous approaches are employed for smart crop development; transformation being the most prevalent one. Transformation requires genetic modulations and various plants like wheat, Arabidopsis and tobacco have been developed with different enhanced traits [6-8].
PTF1 (Inorganic Phosphorus Starvation Induced Transcription Factor 1) is a member of bHLH (basic Helix Loop Helix) family of wheat regulating several genes during phosphorus starvation. It encodes for a 480 amino acid long protein weighing 51.32 kDa. So far, PTF1 from maize, soybean and rice [9-11] has been characterized regulating almost 450 genes involved in secondary root development and enhanced root hair surface area for improved phosphorus absorption [11,12]. The primary function of the TFs (Transcription Factors) is to regulate gene expression by binding to cis-element. Several techniques are used to evaluate the number, position and interaction of cis-element with TF like yeast-two-hybrid, microarray etc. However, yeast-one-hybrid, a relatively advance technique derived from yeast-two-hybrid is highly endorsed due to its varying advantages, reaction simplicity, efficiency and sensitivity [13-16]. Owing to the aforementioned advantages, the current study employed yeast-one-hybrid to study interactions between PTF1 and its regulated genes. The genes were selected on thorough literature review and on the basis of presence of more than one G-box (Binding motif of PTF1) in their promoter regions. The analysis was performed on four selected genes namely LPR1 (Low Phosphate Response), PDR2 (Phosphate Deficiency Response 2), PHT1;2 (Phosphate Transporter) and RNS1 (Ribonuclease) [17-20]. Root development: a post embryonic process is remarkably malleable and relies upon the nutrients present in the soil. Under phosphorus stress, plants undergo a large number of root adaptations for maximum phosphorus absorption [21]. These root system modifications transpire due to root hairs, that perceive low phosphorus levels and initiate diverse genetic pathways which ultimately result in increasing lateral root surface area for enhanced phosphorus absorption and exposure to significantly greater soil volume [22].
The current study is the first reported experiment of wheat PTF1 transformed into Arabidopsis to analyze its effects on root architecture and study its physical interaction with phosphorus stress inducing genes through yeast-one-hybrid. Although, large number of abiotic stress related studies have been done on Arabidopsis, however, up till now, it has not been transformed with plant expression binary vector harboring PTF1.
Methods![]()
YEAST-ONE-HYBRID
Gene selection
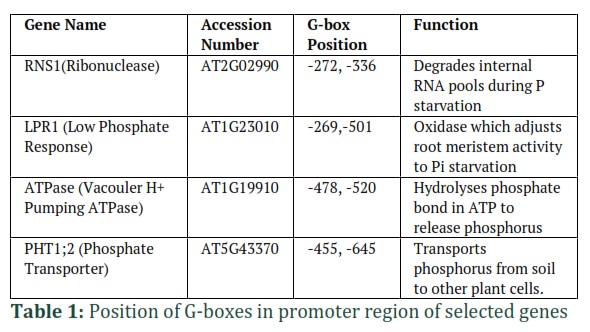
Several genes were identified based on thorough literature review. Motif analysis was done on these genes by Motif finder software and only four genes with palindromic bHLH motif G-Box CTCGTG in their promoter regions were selected. The genes nucleotide sequences were retrieved from TAIR and a region of almost 500 to 600 bp up-stream to the gene was analyzed for G-box presence. The accession number of selected genes along with their functions and the position of G-boxes in their promoter regions are given in table 1.
Promoter cloning in pTUY1H vector
The promoters were cloned in pTUY1H vector following conventional restriction digestion and ligation method. The restriction enzymes Xma1 (NEB catalog # R0180S) with restriction site CCCGGG and Xba1(NEB catalog #R0145S) which cuts at TCTAGA were selected for cloning and added in primers for amplification (Table S1). The digestion reactions were incubated at 37°C for two hours while ligation was done by means of T4 DNA ligase (NEB catalog # M0202S) and incubated at 16°C overnight. Promoters cloning in pTUY1H vector was confirmed through conventional PCR using vector specific primers given in Table S1.
Gateway cloning of PTF1 in pDEST22 vector
Full length PTF1 (sequence retrieved from NCBI accession number DQ979392) was cloned in pDEST22 vector through an intermediate pDONR221 vector following manufacturer’s instructions given in Gateway cloning manual (Invitrogen cat #12535-019). AttB primers were designed by SnapGene software to clone PTF1 in pDONR221 through BP cloning. The confirmed sequenced pDONR221 with attB flanked PTF1 was then cloned in pDEST22 through LR cloning. Vector specific primers given in table S1 were used to confirm PTF1 cloning in pDNOR221 and pDEST22.
Transformation in yeast strains
The positively sequenced plasmids namely RNS1-pTUY1H, LPR1-pTUY1H, ATP-pTUY1H, PHT1;2-pTUY1H were transformed in Y187α yeast strain while PTF1-pDEST22 was transformed in YM4271a strain through heat shock method by giving five minutes’ heat shock at 95°C and then cold shock at -4°C for 2 minutes. Chemi-competent cells prepared from 0.1M calcium chloride solution were used for yeast transformation.
Mating of yeast strains
The yeast strains were grown in full YPD medium (Sigma lot SLBP8224V) at 28°C for two days and a few well-defined colonies were subjected to PCR using vector specific primers described in table S1. The sequenced confirmed yeast strains harboring PTF1, and the four gene promoters were allowed to mate for three days at 28°C by mixing 100µl of PTF1-pDEST22 with 100µl of each pTUY1H vectors.
Growth of mated cultures
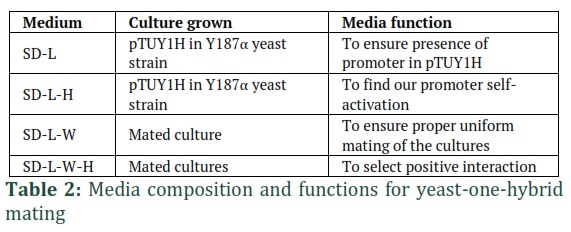
Four different kinds of media namely SD-L (Synthetic defined –Leucine medium), SD-L-W (Synthetic defined -leucine- tryptophan medium), SD-L-H (synthetic defined –leucine –histidine medium) and SD-L-W-H (Synthetic defined –leucine –tryptophan –histidine medium) were prepared to observe the growth of mated cultures and infer results from them. About 7µl of each mated culture was dropped out on each medium and the results were observed after 5 days incubation at 28°C in dark. A histidine inhibitor 3-AT (3 Amino, 1,2,4, trizole) was used in varying concentrations in SD-L-H an SD-L-W-H media for screening and selection. The media composition, the cultures grown on them, and their functions are described in table 2.
TRANSFORMATION
Transformation of wild type Arabidopsis thaliana Col-0 (Columbia-0) variety with plant expression binary vector pSB219 harboring PTF1 transcription factor under ubiquitous CaMV 2X35S promoter and Nos terminator was done by floral-dip method [23]. The putative transformed plants were allowed to mature, and their seeds were properly harvested, dried and stored.
T1 plant growth and screening
The T1 seeds were placed on half MS media and kept at 4°C for two days for stratification. The plates were then placed in light chamber following 16 hours light and 8 hours dark cycle at 22°C for maturation. One-week old plantlet with fibrous shoots were then shifted to moist sand pots for two weeks till they reach 4 rosette stage and were then subjected to PCR screening using optimized PTF1 specific primers (table S1). Phire plant direct PCR master mix kit (Thermo Scientific lot# 00558704) was used for conventional PCR screening following manufacturer’s instructions. The positive plants were further selected, and expression analyzed for PTF1 by real time PCR.
PTF1 Expression Analysis
cDNA synthesis and end point PCR
Total RNA extracted from the positive plants by following the instructions given in InviTrap spin plant RNA mini kit (Stratec molecular lot#MQ180012). Any remaining contamination of DNA was removed by DNAse treatment using TURBO-DNA free kit Ambion (Cat# AM1907). 0.1ng of purified DNAse treated RNA was then used to synthesize cDNA by following manufacturer’s instructions given in RevertAid H Minus first strand cDNA synthesis kit (thermo scientific, USA). End point PCR was conducted to check for the quality of synthesized cDNA by using full length PTF1 primers for template and GADPH primers as internal control using 1µl of cDNA as template in both reactions (table S1).
Real time PCR
Quantitative real time PCR was performed using Maxima SYBR green/ ROX qPCR master mix (cat # K0221). All the reactions were run in triplicates and housekeeping gene GAPDH was used as internal control. The results were analyzed using the built in Bio-Rad CFX manager. The plants with higher expression analysis of PTF1 were allowed to mature and their seeds were properly harvested, dried and stored for further use.
ROOT MORPHOLOGICAL ANALYSIS
A few seeds of selected positive T1 PTF1 over expressing (PTF1-OX) lines were harvested, surface sterilized and then germinated to study root morphology. The dried sterilized seeds were allowed to germinate after stratification on full MS medium for 2 days to grow a single fibrous root. Half of the control and PTF-OX seeds were placed equidistant in vertically standing square petri plates with full MS medium and half to MS-P i.e. without phosphorus supplemented with BASTA herbicide (3mg/ml) for screening, in climate control room at 16°C for two weeks. All the plants in both the media were then subjected to conventional PCR screening and their root morphological analysis was done.
Results![]()
Yeast-one-hybrid
Promoters and PTF1 cloning confirmation
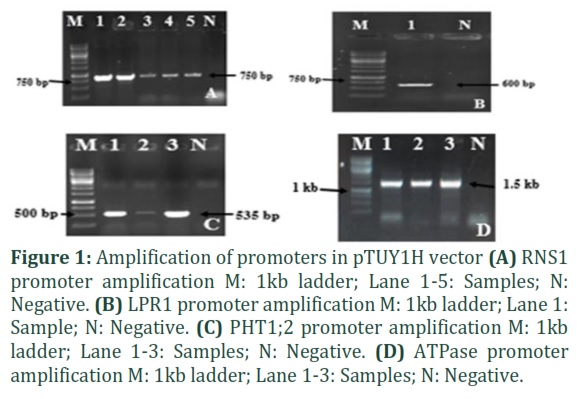
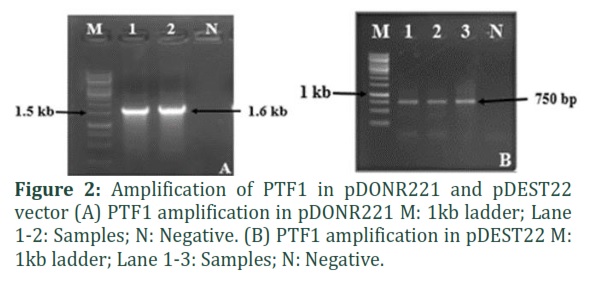
The promoters for the four selected genes were amplified from control Arabidopsis Col-0 DNA and transformed in pTUY1H. Vector specific primers flanking the cloned promoter region were selected for transformant screening. Figure 1 A, B, C, D show the amplification of RNS1, LPR1, PHT1;2 and ATPase in pTUY1H vector respectively. PTF1 transcription factor was cloned in vector pDEST22 through an intermediate vector pDNOR221 via gateway cloning. Figure 2 A,B show the amplification of PTF1 in pDONR221 and pDEST22 respectively.
Yeast Mating Results
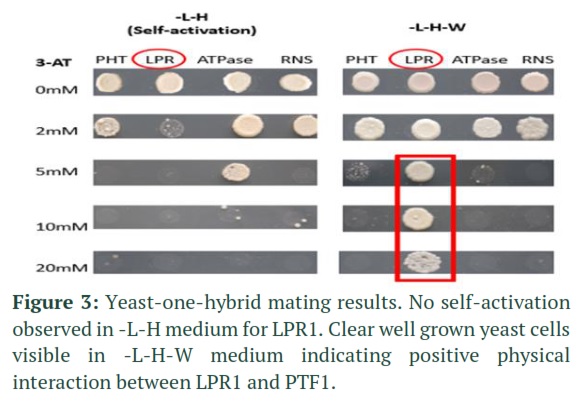
The positively sequenced plasmids harboring gene promoters namely RNS1-pTUY1H, LPR1-pTUY1H, ATP-pTUY1H, PHT1;2-pTUY1H and PTF1-pDEST222 were transformed into yeast strain Y187α and YM4271a respectively. The two yeast strains were allowed to mate and the results were observed after 5 days (figure 3). The results clearly indicated a strong physical interaction between PTF1 and LPR1. Absence of colonies in self-activation (-L-H) medium further validated the finding that this gene is activated only by PTF1. The rest three genes did not give any significant results which may imply some intermediate cross talk or involvement of other genes for their activation under phosphorus stress.
Arabidopsis Screening
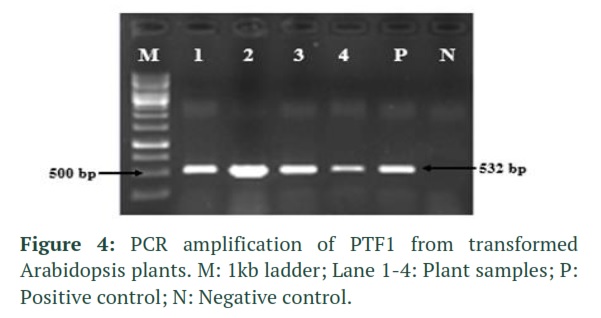
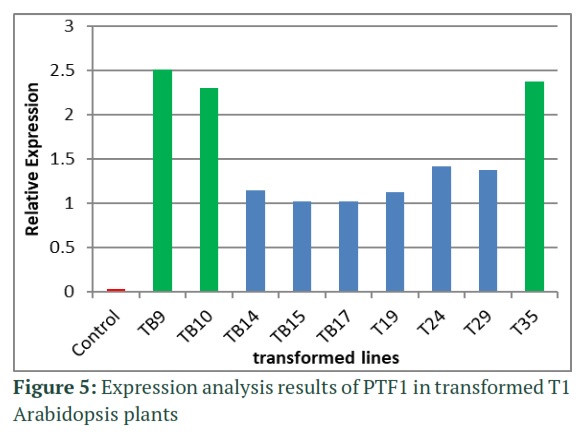
The T1 plants grown in moist sand pots were PCR screened using gene specific PTF1 primers which gave an amplicon of 532bp as shown in figure 4. The positive plants were subjected to RNA extraction and then cDNA synthesis. Quality of cDNA was analyzed by end point PCR using full length primers for PTF1 and GAPDH which was used as internal control. The samples with good quality cDNA were then analyzed for expression analysis by qRT-PCR. The control Columbia-0 variety was used as standard and relative expression of all other lines was measured in comparison to it. Out of 10 lines analyzed, only three lines gave over expression with TB9 giving highest overexpression of 2.5 folds while TB10 and T35 showed 2.2 and 2.3 times more relative expression as compared with GAPDH. (Figure 5).
Root Morphological Analysis
The roots of both PTF-OX lines and control Col-0 lines were analyzed for their length and surface area. A significant increase in root surface area was observed in PTF-OX lines when grown in P-stress medium as compared to their control counterparts. However, in full MS medium, there was no difference observed in roots of either plants. Figure 6 depicts the pictorial evidence of root growth difference.
Figures & Tables

Discussion![]()
DNA protein interactions play an integral part in regulating diverse vital biological processes including transcription, translation, DNA repair etc. Functionally, the most significant element for DNA protein interaction is the conserved motif i.e. specific DNA sequence discernable by the protein as its binding site, usually 5-6 base pairs long [24, 25]. Identification of this conserved motif facilitates in understanding and manipulation of the DNA protein interactions. In the current study, a conserved palindromic motif named as G-box (CTCGTG) for PTF1 was identified (Table 1) using motif finder software [26,27]. Large number of studies employed this software with varying algorithms and programs to find and analyze conserved motifs in genome of different species [28-30].
A large number of systems have been developed to study DNA protein interactions like nanofluidic tools, atomic force microscopy, yeast hybrids etc. [31,32]. Yeast-one-hybrid; a relatively new technique, derived from yeast-two-hybrid, can be applied for rapid and large scale interaction study of TF with hundreds of cis element including promoter, enhancer, silencer, non coding regions and motifs in a single experiment [33]. Other reasons for use of this technique include simple yeast growth conditions along with well optimized protocols and reaction simplicity [13]. Owing to these advantages, the current study employed yeast-one-hybrid study to study interactions between PTF1 and its regulated genes.
The present study is the first of its kind to report physical interaction between phosphorus stress induced genes and PTF1. Various studies have been conducted on phosphorus stress in Arabidopsis focused on its metabolism [34], its leaf and stem anatomical responses [35], its sulpho lipids and galactolipids profile [36] and its fungal microbiota regulation [37]. Of the four selected genes, LPR1 (Low Phosphate Repressor) is the only gene which directly interacts with PTF1 while other genes, although being overexpressed didn’t show direct physical interaction pointing towards some crosstalk or involvement of other genes (Figure 3).
Phosphorus stress prompts the plant to undergo several biochemical and genetic modifications which include phosphorus remobilization, release of organic acids from root tips etc. [38-40]. However, roots of the plant are the most transformed and altered structure during phosphorus stress. Root hair architecture is modified significantly with most obvious difference being observed in increased root hair density and lateral root surface area [41]; [12,42]. Likewise, in the present study a momentous variation was observed in root hair and lateral surface area of Arabidopsis when grown vertically under phosphorus deficient MS medium (Figure 6).
Funding Agency
The project has been funded by Pakistan Agriculture Research Council (PARC) under the Agricultural Linkages Program (ALP) Project number 313.
Author Contributions
Aqsa Akhtar conducted the experiments and structured the manuscript. Dr Muhammad Irfan and Dr Asma Maqbool helped in research design and data analysis. Dr. Kauser Abdulla Malik designed the research and also helped in refining the manuscript.
The authors declare that there is no conflict of interest regarding the publication of this paper.
References
- Baker A, Ceasar SA, Palmer AJ, Paterson JB, Qi W, et al. Replace, reuse, recycle: improving the sustainable use of phosphorus by plants. Journal of experimental botany, (2015); 66(12): 3523-3540.
- Tam THY, Catarino B, Dolan L. Conserved regulatory mechanism controls the development of cells with rooting functions in land plants. Proceedings of the National Academy of Sciences, (2015); 112(29): E3959-E3968.
- Taliman NA, Dong Q, Echigo K, Raboy V, Saneoka H. Effect of phosphorus fertilization on the growth, photosynthesis, nitrogen fixation, mineral accumulation, seed yield, and seed quality of a soybean low-phytate line. Plants, (2019); 8(5): 119.
- Sun B, Gao Y, Wu X, Ma H, Zheng C, et al. The relative contributions of pH, organic anions, and phosphatase to rhizosphere soil phosphorus mobilization and crop phosphorus uptake in maize/alfalfa polyculture. Plant and Soil, (2020); 447(1): 117-133.
- Antoniadis V, Koliniati R, Efstratiou E, Golia E, Petropoulos S. Effect of soils with varying degree of weathering and pH values on phosphorus sorption. Catena, (2016); 139214-219.
- Abid N, Khatoon A, Maqbool A, Irfan M, Bashir A, et al. Transgenic expression of phytase in wheat endosperm increases bioavailability of iron and zinc in grains. Transgenic research, (2017); 26(1): 109-122.
- Lu M, Ying S, Zhang D-F, Shi Y-S, Song Y-C, et al. A maize stress-responsive NAC transcription factor, ZmSNAC1, confers enhanced tolerance to dehydration in transgenic Arabidopsis. Plant Cell Reports, (2012); 31(9): 1701-1711.
- Yan H, Jia H, Chen X, Hao L, An H, et al. The Cotton WRKY Transcription Factor GhWRKY17 Functions in Drought and Salt Stress in Transgenic Nicotiana benthamiana Through ABA Signaling and the Modulation of Reactive Oxygen Species Production. Plant and Cell Physiology, (2014); 55(12): 2060-2076.
- Li Z, Liu C, Zhang Y, Wang B, Ran Q, et al. The bHLH family member ZmPTF1 regulates drought tolerance in maize by promoting root development and abscisic acid synthesis. Journal of experimental botany, (2019); 70(19): 5471-5486.
- Li X, Wu B, Kong Y, Zhang C. GmPTF1, a novel transcription factor gene, is involved in conferring soybean tolerance to phosphate starvation. Genetics and Molecular Research, (2014); 13(1): 926-937.
- Yi K, Wu Z, Zhou J, Du L, Guo L, et al. OsPTF1, a novel transcription factor involved in tolerance to phosphate starvation in rice. Plant physiology, (2005); 138(4): 2087-2096.
- Janes G, von Wangenheim D, Cowling S, Kerr I, Band L, et al. Cellular patterning of Arabidopsis roots under low phosphate conditions. Frontiers in plant science, (2018); 9735.
- Lopato S, Bazanova N, Morran S, Milligan AS, Shirley N, et al. Isolation of plant transcription factors using a modified yeast one-hybrid system. Plant Methods, (2006); 2(1): 3.
- Breton G, Kay SA, Pruneda-Paz JL (2016) Identification of Arabidopsis transcriptional regulators by yeast one-hybrid screens using a transcription factor ORFeome. Environmental Responses in Plants: Springer. pp. 107-118.
- Reece-Hoyes JS, Walhout AM. Yeast one-hybrid assays: a historical and technical perspective. Methods, (2012); 57(4): 441-447.
- Castrillo G, Turck F, Leveugle M, Lecharny A, Carbonero P, et al. Speeding cis-trans regulation discovery by phylogenomic analyses coupled with screenings of an arrayed library of Arabidopsis transcription factors. PloS one, (2011); 6(6): e21524.
- Xu JM, Wang ZQ, Wang JY, Li PF, Jin JF, et al. Low phosphate represses histone deacetylase complex1 to regulate root system architecture remodeling in Arabidopsis. New Phytologist, (2020); 225(4): 1732-1745.
- Abel S. Phosphate sensing in root development. Current opinion in plant biology, (2011); 14(3): 303-309.
- Liu X, Zhao X, Zhang L, Lu W, Li X, et al. TaPht1; 4, a high-affinity phosphate transporter gene in wheat (Triticum aestivum), plays an important role in plant phosphate acquisition under phosphorus deprivation. Functional Plant Biology, (2013); 40(4): 329-341.
- Jost R, Pharmawati M, Lapis-Gaza HR, Rossig C, Berkowitz O, et al. Differentiating phosphate-dependent and phosphate-independent systemic phosphate-starvation response networks in Arabidopsis thaliana through the application of phosphite. Journal of experimental botany, (2015); 66(9): 2501-2514.
- Dong J, Piñeros MA, Li X, Yang H, Liu Y, et al. An Arabidopsis ABC transporter mediates phosphate deficiency-induced remodeling of root architecture by modulating iron homeostasis in roots. Molecular plant, (2017); 10(2): 244-259.
- Ma Z, Bielenberg D, Brown KM, Lynch JP. Regulation of root hair density by phosphorus availability in Arabidopsis thaliana. Plant, Cell & Environment, (2001); 24(4): 459-467.
- Clough SJ, Bent AF. Floral dip: a simplified method for Agrobacterium -mediated transformation of Arabidopsis thaliana. The Plant Journal, (1998); 16(6): 735-743.
- Li C, Tao Y, Yang Y, Xiang Y, Li G. In Vitro Analysis of DNA–Protein Interactions in Gene Transcription Using DNAzyme-Based Electrochemical Assay. Analytical chemistry, (2017); 89(9): 5003-5007.
- Niranjani G, Murugan R. Generalized theory on the mechanism of site-specific DNA–protein interactions. Journal of Statistical Mechanics: Theory and Experiment, (2016); 2016(5): 053501.
- Gordân R, Shen N, Dror I, Zhou T, Horton J, et al. Genomic regions flanking E-box binding sites influence DNA binding specificity of bHLH transcription factors through DNA shape. Cell reports, (2013); 3(4): 1093-1104.
- Sun X, Wang Y, Sui N. Transcriptional regulation of bHLH during plant response to stress. Biochemical and biophysical research communications, (2018); 503(2): 397-401.
- Kang K, Chung JH, Kim J. Evolutionary Conserved Motif Finder (ECMFinder) for genome-wide identification of clustered YY1-and CTCF-binding sites. Nucleic acids research, (2009); 37(6): 2003-2013.
- Wang T, Kettenbach AN, Gerber SA, Bailey-Kellogg C. MMFPh: a maximal motif finder for phosphoproteomics datasets. Bioinformatics, (2012); 28(12): 1562-1570.
- Schmidt MC, Rocha AM, Padmanabhan K, Chen Z, Scott K, et al. Efficient α, β-motif finder for identification of phenotype-related functional modules. BMC Bioinformatics, (2011); 12(1): 440.
- Kasas S, Dietler G. DNA-protein interactions explored by atomic force microscopy; 2018. Elsevier. pp. 231-239.
- Frykholm K, Nyberg LK, Westerlund F. Exploring DNA–protein interactions on the single DNA molecule level using nanofluidic tools. Integrative Biology, (2017); 9(8): 650-661.
- Sewell JA, Bass JIF (2018) Options and considerations when using a yeast one-hybrid system. Two-Hybrid Systems: Springer. pp. 119-130.
- Fukushima A, Iwasa M, Nakabayashi R, Kobayashi M, Nishizawa T, et al. Effects of combined low glutathione with mild oxidative and low phosphorus stress on the metabolism of Arabidopsis thaliana. Frontiers in plant science, (2017); 81464.
- Cai Q, Ji C, Yan Z, Jiang X, Fang J. Anatomical responses of leaf and stem of Arabidopsis thaliana to nitrogen and phosphorus addition. Journal of Plant Research, (2017); 130(6): 1035-1045.
- Wang F, Ding D, Li J, He L, Xu X, et al. Characterisation of genes involved in galactolipids and sulfolipids metabolism in maize and <i>Arabidopsis</i> and their differential responses to phosphate deficiency. Functional Plant Biology, (2020); 47(4): 279-292.
- Fabiańska I, Gerlach N, Almario J, Bucher M. Plant-mediated effects of soil phosphorus on the root-associated fungal microbiota in Arabidopsis thaliana. New Phytologist, (2019); 221(4): 2123-2137.
- Gutiérrez-Alanís D, Ojeda-Rivera JO, Yong-Villalobos L, Cárdenas-Torres L, Herrera-Estrella L. Adaptation to phosphate scarcity: tips from Arabidopsis roots. Trends in plant science, (2018); 23(8): 721-730.
- Kisko M, Shukla V, Kaur M, Bouain N, Chaiwong N, et al. Phosphorus transport in Arabidopsis and wheat: Emerging strategies to improve P pool in seeds. Agriculture, (2018); 8(2): 27.
- Wu L, Kobayashi Y, Wasaki J, Koyama H. Organic acid excretion from roots: a plant mechanism for enhancing phosphorus acquisition, enhancing aluminum tolerance, and recruiting beneficial rhizobacteria. Soil Science and Plant Nutrition, (2018); 64(6): 697-704.
- Kong Y, Wang B, Du H, Li W, Li X, et al. GmEXLB1, a soybean expansin-like B gene, alters root architecture to improve phosphorus acquisition in Arabidopsis. Frontiers in Plant Science, (2019); 10.
- Janes G (2018) Factors regulating cortex cell file proliferation under low phosphorus stress in Arabidopsis thaliana roots: University of Nottingham.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0