

Full Length Research Article
Systems genomics of nucleoporins provides prognostic insights into breast cancer
Rashid Mehmood1*, Kazuya Jibiki2, Zakeya J. Alsafwani1, Muhammad Naseem3, Noriko Yasuhara2*
Adv. life sci., vol. 9, no. 1, pp. 98-110, May 2022
*– Corresponding Author: Rashid Mehmood (Email: rmehmood@alfaisal.edu)
Noriko Yasuhara (Email: yasuhara@chs.nihon-u.ac.jp)
Authors' Affiliations
2. Graduate School of Integrated Basic Sciences, Nihon University, Setagaya-ku, Tokyo, Japan
3. Department of Life and environmental Sciences, College of Health and Natural Sciences, Zayed University, Abu Dhabi, UAE; Department of Bioinformatics, Biocenter, University of Würzburg, Am Hubland, Wuerzburg, Germany
Correction Notice 14/07/2021: This article contains non-technical corrections in the previous version published on 25/05/2022.
Editorial Expression of Concern
20 June 2025: Following publication of this paper, the internal audit (consequent to concerns on quality raised by Web of Science) notified Advancements in Life Sciences about suspected plagiarism. By this Editorial Expression of Concern, we alert the scientific community of the errors as we reconcile the records.
Editorial Note
25 June 2025: While rerunning the Turnitin originality analysis, similarity index of 8% from a single source was found for this article (out of a total of 17%). Editorial board of Advancements in Life Sciences has started the process of retracting this article due to the above post-publication findings. The process shall be concluded after registering responses from the authors. Meanwhile, full text of the article shall remain unavailable for citations (this notice has been updated following insights derived from relevant COPE cases and the industry standards). Show of cause notice has also been issued to the concerned editorial team member.
Rescinded: Editorial Expression of Concern
27 July 2025: Editorial expression of concern issued on 20 June 2025 is hereby rescinded on account of editorial review of the found similarity in light of COPE guidelines. Since the total similarity index is within the permissible limit i.e., <20%, and the source of the extraordinary similarity of 8% in Introduction and Methods sections is an article published previously by authors of this study. Therefore, in light of the Text Recycling guidelines of COPE, the Chief Editor is rescinding editorial expression of concern seeing details of the similarity between both articles (this one and the article published in 2021).
The board has decided to keep the contents of this article unchanged.
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Nucleoporins (Nups) constitute a large group of proteins that are structurally arranged at the nuclear envelope and facilitate the bidirectional movement of molecules across the nuclear membrane. In addition to regulating the shuttling of ribonucleoprotein complexes, RNAs and proteins, various Nups interact with chromatin either directly or indirectly, thus regulating gene expression. Any mutations or expression anomalies of Nups may lead to abnormal localization of critical molecules, or dysregulated expression of genes that they interact with. A comprehensive genomic study encompassing all Nup genes in relation to breast cancer is lacking.
Methods: We used genomic and transcriptomic datasets from Pan-Cancer TCGA (The Cancer Genome Atlas), Genotype-Tissue Expression (GTEx) and microarray platforms and conducted in silico analysis of all the genes encoding nucleoporins that are associated with the Nuclear Pore Complexes (NPCs). For mutation detection, we used cBioportal; for expression analysis, we used Xena and for patient survival plots, KMPlot was utilized.
Results: The genetic and molecular profile of nucleoporin genes identified multiple mutations and detected aberrant expression in breast cancer. Interestingly, NUP133, AHCTF1, TPR, Nup121L showed simultaneous gene amplification in nearly 10% of breast cancer patients. In addition, deregulated expression of some of the nucleoporins, namely, NUP62, NUP 93, NUP98, NUP155, POM121L12, RAE1, SEC13, TPR were correlated with patient prognosis.
Conclusion: The current study is the first one that unravels a comprehensive molecular and genetic profile of nucleoporins genes in breast cancer and underscores the critical roles of various nucleoporins in cancer progression. The identified molecules may advance our understanding of the etiology of the disease and serve as possible targets for novel therapeutic strategies in cancer.
Keywords: Nuclear Pore Complexes (NPCs); Nucleoporins; Breast cancer; Bioinformatics
Introduction![]()
Nuclear pore complexes (NPCs), embedded in the nuclear envelope, provide a gateway for the movement of molecules in and out of the nucleus. The barrier function of the nuclear envelope is quintessential not only for maintaining the distinct composition of the nucleus but also for the unique functions therein. Two types of movements take place through the NPCs; passive diffusion that involves movements of small sized proteins, metabolites, and ions, and facilitative transport wherein large molecules including ribonucleoprotein complexes, RNAs and proteins cross the nuclear envelope. The facilitative transport requires particular transport receptors, called importins and exportins. Both of these types of transport are mediated by the NPCs [1]. NPCs are megadalton complexes (approximately 60-125MDa) and are composed of ~30 distinct proteins called nucleoporins (Nups), which provide binding sites for transport receptors for the transport of molecules in and out of the nucleus [2].
The NPCs are remarkably complicated structures and are composed of three structurally distinct regions. 1) the cytoplasmic ring from which filaments extend into the cytoplasm, 2) the nuclear basket that extends to the nucleoplasmic faces of the NPCs and, 3) the central framework which makes up the pore [3]. The correct positioning of various Nups in the NPC warrants their structural and functional roles, facilitating the transport of selective molecules [4]. The NPC assembly requires coordination between NUPs. For instance, Nup133, a component of the NPC scaffold, is required for Tpr and Nup153 assembly into the nuclear basket. For the transport of molecules into the nucleus, the karyopherins that bind NLS (Nuclear Localization Signal) containing cargoes interact with FG (Phe-Gly) repeats containing Nups of the NPCs and mediate the translocation of the transport complex [5]. Once into the nucleus, the association between Nups and importins is disengaged by the action of RanGTP by virtue of its very high binding affinity to importins [6]. Conversely, the nuclear export of a number of proteins is mediated by interaction of Exportins with RanGTP, while RanGTP independent export also exist, as in case of mRNAs [7].
In addition to their well-known role in the transport of molecules across the nuclear membrane, some of the Nups have been shown to regulate a variety of functions, including RNA processing, transcriptional activation, and chromatin modulation [8-10]. Their widespread functions are strengthened by the fact that some NUPs (i.e. NUP98, NUP62, NUP50, NUP153) are not only localized to the NPCs; instead, they exhibit dynamic features by moving off and on of the NPCs. Their shuttling in and out of the nucleus and interaction with cytosolic and nuclear components point to their unorthodox functions. An interesting correlation was found between the expression levels of genes and their position in the vicinity of the NPCs [11]. The observation that the genomic regions nearby the NPCs are transcriptionally active led to the identification of their roles in transcriptional regulation [12, 13]. NUP53, for instance, was shown to not only bind DNA, but its binding pattern was co-enriched with other transcriptional activation markers, i.e., Histone H4K16 acetylation and RNA polymerase II [14]. Over the years, the roles of a battery of Nups were identified in regulating the chromatin landscape [15]. A subset of the Nups regulate expression of genes implicated in cellular identity thus regulating cellular differentiation and development [16, 17]. Moreover, recently, their roles in viral infection and innate immunity is also described [85, 86]. Therefore, it is not surprising that aberrant expression patterns of various Nups are associated with a wide range of medical conditions [18, 19].
Genomic fusions involving Nups with other partners have been found in hematopoietic and non-hematopoietic malignancies [20, 21]. Additionally, the deregulated expression and mutations of some Nups are also linked with various cancers [22, 23]. Their roles in cancer progression may be due to specifically expediting the nuclear translocation of oncogenes, e.g., E2F1, MYC, thus enhancing their oncogenic activities [24] or through interacting with chromatin, thus regulating chromatin remodeling, and regulating the transcription of genes [25]. In breast cancer, only few studies have looked into the individual Nups for their roles in cellular transformation. NUP88 mRNA overexpression, for instance, has been shown to be associated with high aggressiveness of breast cancer [26]. Inverse correlation of NUP93 expression with the survival of triple-negative breast cancer patients by modulating cytoskeleton remodeling has been recently shown [27]. However, to our knowledge, a comprehensive study of all the nucleoporins cataloging their mutational profile, differential expression compared with the normal tissue and their correlation with breast cancer patient survival is lacking. In the current study, using genomic and transcriptomic datasets from The Cancer Genomic Atlas (TCGA) and microarray platforms, we analyzed all the known genes encoding components of NPCs in breast cancer. Our data suggest that majority of NUPs are mutated/aberrantly expressed in breast cancer, albeit with varying degrees, and point to the implications of NUPS in patients’ prognosis.
Methods![]()
Genetic alterations in Nup genes
In order to identify mutations in various Nups, we utilized TCGA (The Cancer Genome Atlas) data using cBioportal [28 ,29]. At the cBioportal, breast cancer datasets are available from multiple sources. We selected the latest TCGA Breast Invasive Carcinoma (Firehouse legacy). In this category, there are 963 patients that had both mutation and CNA (Copy number alterations) data. Therefore, following genetic alterations were scanned in these 963 breast cancer patients' datasets; 1) Mutations, 2) CNA (copy number alterations). Ethical approval was not needed for the study as all datasets in this study were utilized from public databases (TCGA), and processing of data were according to the TCGA publication guidelines (https://cancergenome.nih.gov/publications/guidelines).
Nups and encoding genes nomenclature
Nucleoporins constitute all the proteins that make up the nuclear pore complexes. However, before their identification as a part of this huge nuclear pore complexes, they adapted different names based on interacting proteins and functions they were associated with. For the sake of consistency, we used Human Genome Organization (HUGO) gene nomenclature committee resource (https://www.genenames.org/). All the genes corresponding to nucleoporins were identified from this HUGO resource and published literature [30]. The list of Nups encoded by respective genes are listed in Table 1.
NUPs Interactome and network analysis
We mapped protein-protein interactions among NUPs specific proteins as high confidence interactome retrieved from STRING database (https://string-db.org/) [87]. The list of NUPs genes (tables 1) was given as input identifiers to the muti-proteins search option of the STRING database. A stringent criterion for the selection of interactions as combined edge score greater than 0.9 (P-values for Bayesian confidence score) resulted in a densely connected network, composed of 35 nodes and 175 edges. For network analysis we used STRING inbuilt analysis function for degree and clustering co-efficient. We only selected experimental interactions and visualized the network in evidence mode as STRING output. The edges denote both functional and physical protein associations among the nodes as opted in the basic settings options.
Gene expression changes compared with the normal tissue
For expression comparison between normal tissue and breast cancer samples, we used Xena [67] which incorporates data from TCGA datasets and GTEX for comparison between the two [68].
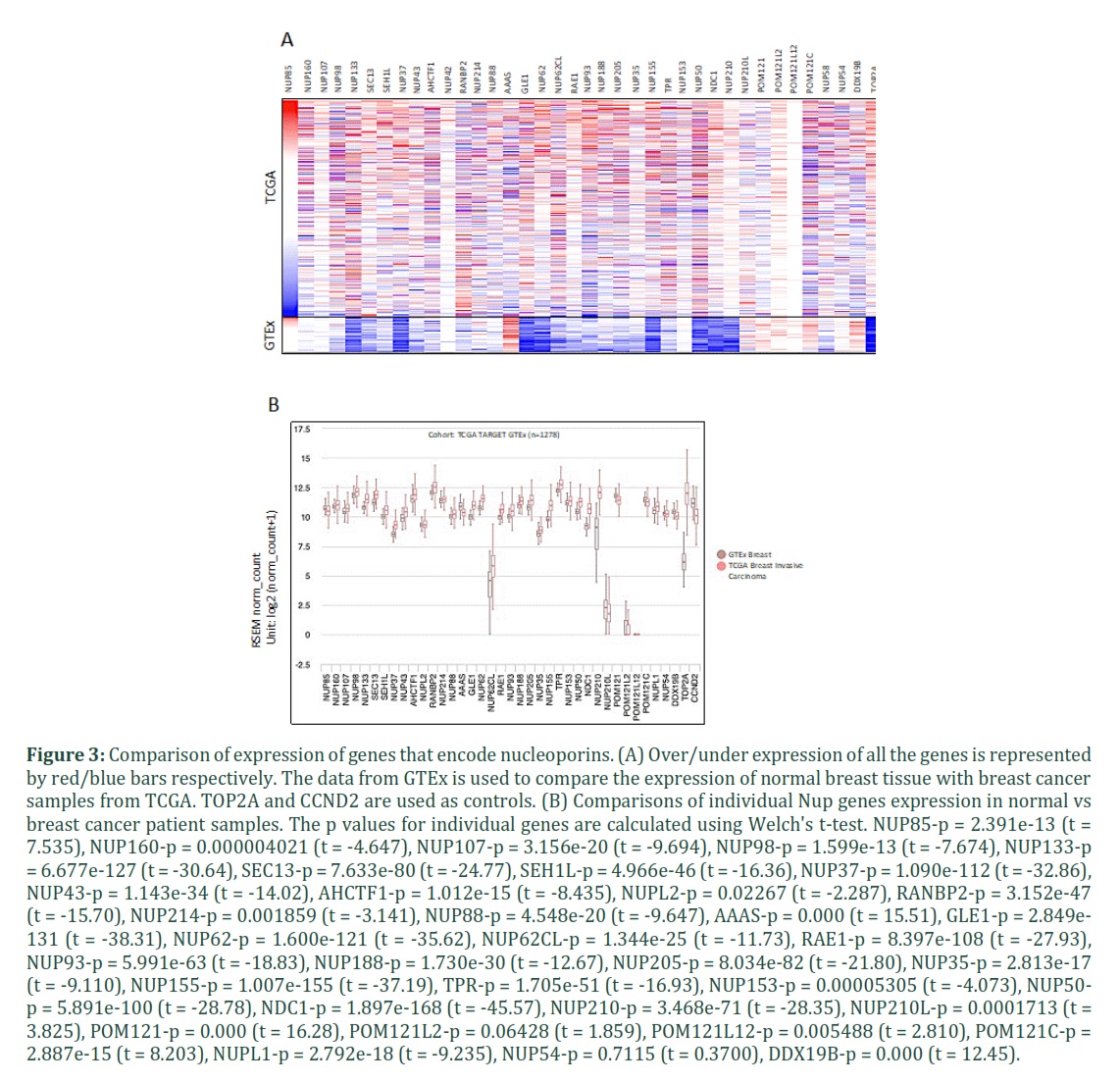
The gene list in the table 1 was used to generate heat map. All the genes were incorporated in the UCSC Xena. After launching Xena, the first variable phenotype “main category” was selected. In the second Genomic variable, Gene expression was chosen, followed by incorporating all the genes encoding Nups. TCGA GTEX datasets was used to generate heatmap. Additionally, box plots for the individual genes were fetched from the “view chart” to make gene expression comparisons between GTEX and TCGA breast cancer patients' datasets. As Xena utilizes the same pipeline for TCGA and GTEx samples, therefore, both datasets are re-analyzed (using UCSC Toil RNA-seq recompute compendium) to eliminate batch effects [67].
Patient survival plots
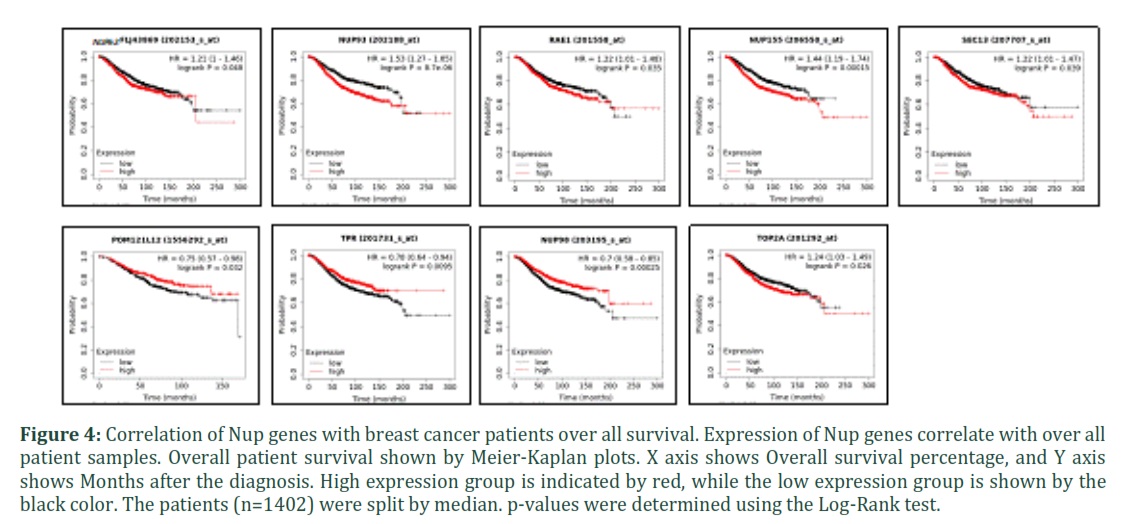
Kaplan-Meier Plotter was used to generate Meier-Kaplan plots wherein multiple microarray expression datasets are incorporated from 7462 breast cancer patients [69]. In order to generate overall survival plots, the patients were split by median. For the best probe selection, “jetset” was selected for the shown time periods. The indicated p value does not include correction for multiple hypothesis testing by default [69].
Results![]()
Mutational landscape of various Nups in breast cancer
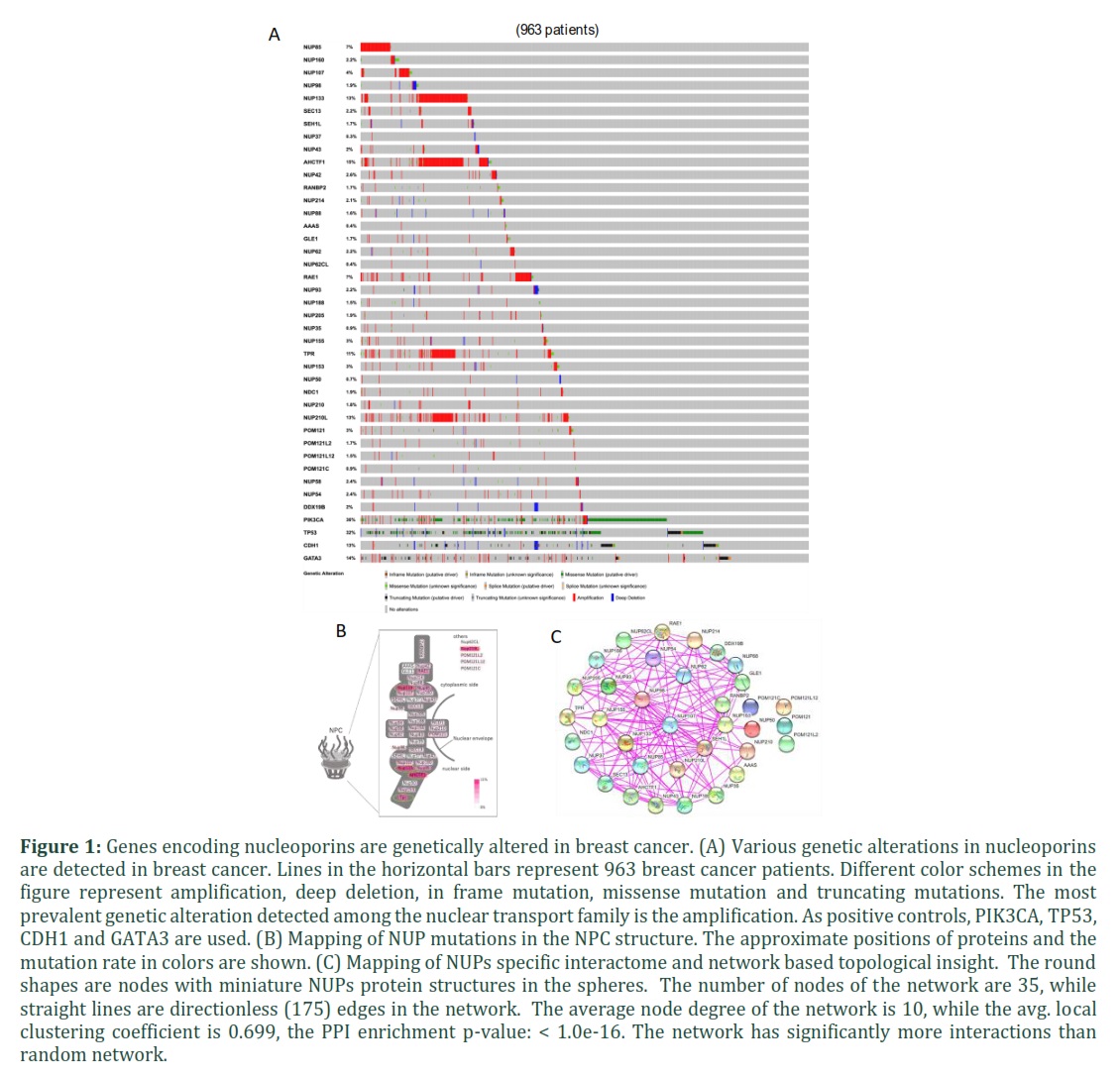
In order to delineate the roles of nuclear transport process in breast cancer progression, we recently carried out bioinformatic analysis of TCGA datasets and showed that various genes encoding factors related to nuclear transport are mutated [70]. An equally important aspect of the nuclear transport process is the interaction of transport factors with the components of the NPCs and subsequent translocation to the nucleus. Therefore, mutations in the genes coding for Nups, the proteins that constitute the NPCs, may disrupt/expedite the nuclear transport process. Thus, it is imperative to identify various mutations in this category of genes. Therefore, using TCGA datasets, we characterized mutations in Nup genes and found that majority of the Nup genes were genetically altered in breast cancer patients. The alteration frequency differed markedly (ranging from 0.4% in AAAS to 15% in AHCTF1) (Figure 1) amongst various Nups. Interestingly, gene amplification appeared to be the most prevalent genetic anomaly. Additional mutations were also detected, including deep deletions, base substitutions and multiple alterations. A synergistic gene amplification in AHCTF1, NUP133, NUP210L and TPR was found in over 10% of patients. Figure 1B shows the frequency of mutations in the components of nuclear envelope based on their location in the nuclear pore. To further expand on the possible consequences of genetic alterations on the nuclear pore architecture and to get functional insights into the NUPs genes, we mapped physical interactions among their encoded proteins. A densely connected network of NUPs proteins was derived from the publicly available STRING database [87]. As shown in the Figure 1C, there are 35 protein nodes in the network that constitute 175 edges with diverse functional implications. Interactions between Nup133, AHCTF1 and Nup210L, that are synergistically amplified, are clearly visible in the interactome. Interestingly, alteration frequency in all Nup genes was found to be in nearly 50% of the patients representing significance of this set of molecules (Supplementary figure 1A). Genetic alterations in Nups genes were detected in all breast cancer subtypes albeit with variable rates and types of genetic alterations. In order to validate our analysis, we used PIK3CA, TP53, CDH1, and GATA3. Consistent with literature, we found high prevalence of mutations in these control genes (Figure 1A).
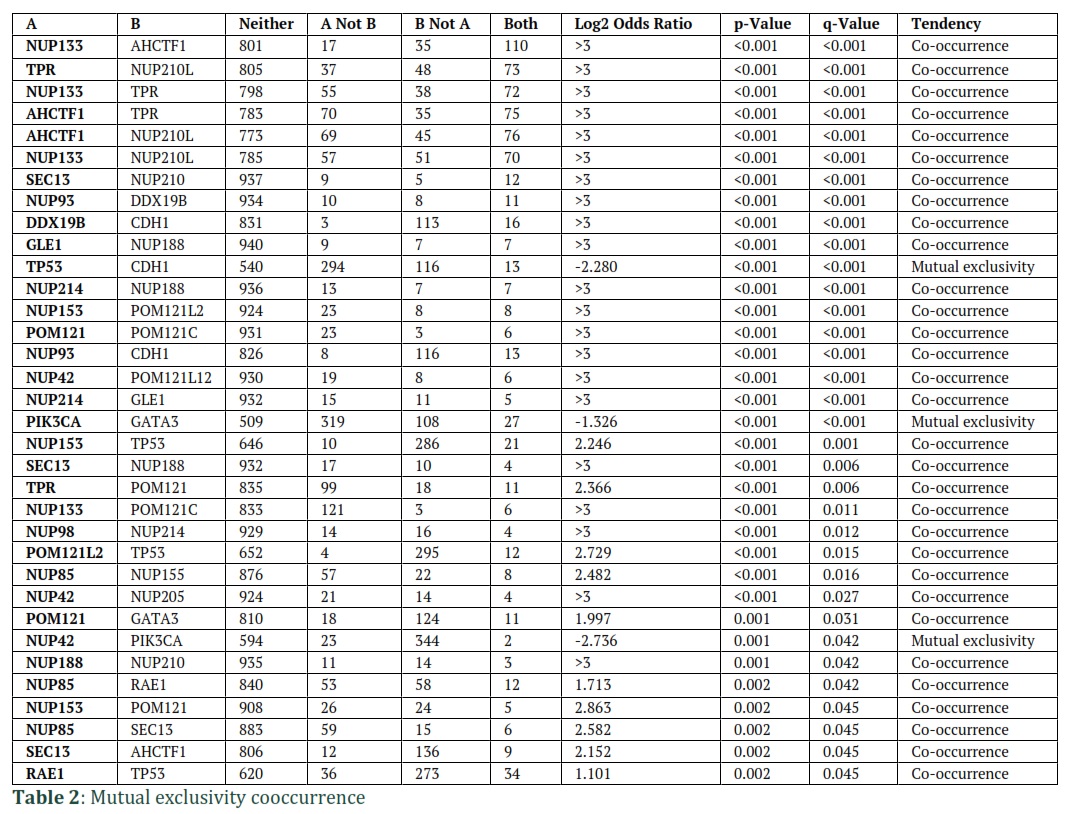
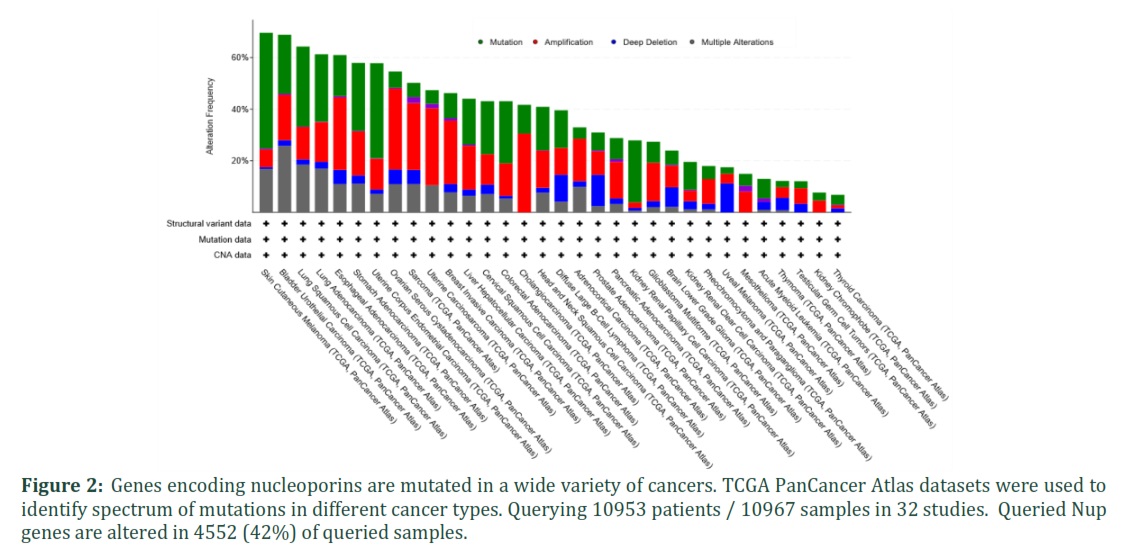
After establishing that Nup genes are mutated in breast cancer, we stratified breast cancer patients into metaplastic breast cancer, breast invasive ductal carcinoma, breast mixed ductal and lobular carcinoma, breast invasive locular carcinoma, and breast invasive mixed mucinous carcinoma and looked for mutations (Supplementary figure 1B). Additional analysis also identified co-occurrences and mutual exclusivities of these top mutated genes with several Nups (Table 2). The frequency of genetic alterations was not specific to breast cancer. We utilized TCGA PanCancer Atlas Studies that has datasets from a variety of cancers and identified similar trends in mutations in majority of the cancers. This shows that mutations in Nup genes span a spectrum of cancers (Figure 2).
Nups related genes undergo massive expression anomalies in breast cancer
Recent transcriptomic analysis showed massive transcriptomic deregulation is an important hallmark of cancer. A number of genes are not only aberrantly expressed in breast cancer, but may also serve as biomarkers. Therefore, we analyzed RNA levels of all Nup genes compared with the normal tissue. The cancer patient’s data was from TCGA, and for comparison GTEx datasets were used. As can be seen in Figure 3A, several Nup genes showed deregulated expression in breast cancer. In majority of the genes analyzed, overexpression was detected to be major anomaly in cancer patients compared with the samples from GTEx. However, a few genes including AAAS, POM121, and DDX19B showed reduced expression in cancer patients compared with the GTEx cohort. The high expression in TCGA datasets might be associated with high amplification rate, at least partially. Figure 3B shows comparisons of individual gene expression between TCGA and GTEx datasets. Collectively, majority of genes encoding Nups go through massive deregulation in expression in cancer patients.
Nups with correlation with patient overall survival.
Having established that genetic aberrations and expression changes in cancer patients prevail significantly, next was to determine if any of these changes in gene expression in Nups have a prognostic potential. Mining various microarray datasets using Meier-Kaplan (KM) Plotter [69], we calculated overall survival (OS) of breast cancer patients based on expression of Nup genes. Out of all the Nup genes tested, NUP62, NUP93, RAE1, NUP155, and SEC13 overexpression showed poor overall survival, while POM121L12, TPR, and NUP98 overexpression were correlated with better patient survival. TOP2A, a well-known marker for poor patient survival is also shown in the figure 4, which, consistent with its established role, showed poor patient OS. All other Nups that did not show any significant correlation with patient overall survival are shown in Supplementary figure 2. Overall, the analysis identifies several molecules that might have prognostic potential in breast cancer.
Figures & Tables
Discussion![]()
In this study, we describe a comprehensive genetic and molecular landscape of nucleoporins genes in breast cancer. Our analysis shows that a number of Nup genes are not only mutated but also undergo drastic expression changes in breast cancer.
Among the Nup genes, we found that AHCTF1, NUP133, NUP210L and TPR were synergistically altered in around 10% of patients. Nup133 is located in the NPC scaffold forming the Y-complex together with Nup85, SEC13, Nup160, Nup107 [88]. It has been shown that Nup133 is instrumental in formation of the nuclear basket with its role in the Tpr and Nup153 recruitment to the NPC basket [89]. Moreover, Tpr is necessary for the nuclear export of mRNAs and proteins. Thus, if the gene products of NUP133 and TPR genes are synergistically altered, the assembly and the functions of NPC could be significantly altered. AHCTF1, also known as ELYS, is not only necessary for post-mitotic NPC assembly through recruitment of chromatin but also regulates the nuclear size [90]. Therefore, simultaneous changes in the quality and quantity of these NUPs could have a significant impact on the function of the nuclear pores, which may define the behavior of cancer.
We recently catalogued genetic and molecular attributes of all the genes encoding nuclear transport factors in breast cancer [70]. A crucial part of the nuclear transport process is the interaction of Nups with the actual transport receptors. Nups constitute a group of proteins that constitute the nuclear pore complexes. Their critical role in regulating transport of molecules across the nuclear membrane is already established [71]. One of the categories of cargo that traverse through the nuclear membrane is the proteins related to regulation of cell cycle. Therefore, the functions of Nups in cell cycle regulation become pivotal. Mislocalization of cell cycle regulators has been linked to various cancers [24]. Our knowledge of individual Nups is rudimentary with limited evidence of NUP43, NUP88 and NUP98 to be drivers of breast cancer [72, 73]. However, an overarching molecular profiling of Nup genes in cancer was lacking. Our study fills this gap by providing comprehensive analysis of all Nup genes in breast cancer.
Most popular and known association of Nups with various malignancies including leukemiac and non leukemiac are through juxtaposition of nup genes with a wide range of other genes [20, 21]. Nup98, for instance, is frequently identified to be translocated with a number of factors and mediate transcription changes depending on its fusion partner [74]. However, there is little knowledge regarding the role of individual Nups in cancer progression. Their molecular analysis of individual role of Nups is critical as they perform diverse aspects of cell cycle either by regulating the nucleocytoplasmic shuttling of various molecules implicated in cancer or by direct or indirect interaction with chromatin., thus regulating expression of genes with possible implications with cancer. An evidence in support of the first aspect was recently reported in prostate cancer wherein overexpression of POM121 led to enhanced nuclear import of E2F1, MYC, and AR, eventually affecting tumorigenesis, proliferation, and survival in lethal prostate cancer [24]. We identified that, in breast cancer, several other Nups were either mutated or had deregulated expression. Interestingly, some of them are implicated in directing nuclear transport by interacting with transport receptors [75], implicating possible nuclear transport related functions.
In addition to above mentioned functions of Nups, evidence regarding their un-expected role in coordinating cell cycle regulation is being established [91]. A recent report highlighted the role of Nup155 in regulating translation of P21 which, a key downstream effector of P53 response in liver cancer, thus regulating P53 network [76]. Nup155 also emerged as an important modulator of chromatin remodeling by interaction with HDAC4 [77]. More recently, Nup155 has been shown to regulate non-coding RNA network mouse embryonic stem cells (mESCs). Partial loss of Nup155 led to reduction in abundance of miR290–295 cluster with concomitant loss of pluripotency factors in mES cells thus affecting pluripotency [78]. The involvement of a single nucleoporin in a wide variety of signaling networks including pluripotency underscores widespread functions. Our data show that NUP155 is not only significantly upregulated in breast cancer but also correlated with poor patient survival. It remains to be seen if and how individual pathways regulated by Nup155 converge onto the pathogenic phenotype in breast cancer.
Another novel molecule that was identified through our in silico analysis is SEC13. Although SEC13 is generally associated with COPII vesicle for the anterograde movement of molecules from ER to Golgi, its location in the nuclear pore is well established where it is a part of Nup107-160 subcomplex [37]. Additionally, its nucleocytoplasmic shuttling of SEC13 is also reported [79]. In the nucleus, it is also shown to bind kinetochores [37]. Its multi-compartmental distribution indicates a variety of possible functions. Our analysis indicates that it is not only significantly upregulated, but also correlates with poor patient survival, thus warranting deeper exploration.
It is pertinent to note that majority of Nups were upregulated in breast cancer patients (Figure 3). While expression of most of them is correlated with poor patient survival, thus pointing to their roles as oncogenes, overexpression of some Nups correlated with better over all patient survival. The Nups in this category included Nup98, TPR and POM121L12. The correlation between their overexpression and better patient survival might provide a hint for their possible tumor suppressor functions. Further studies are required to realize their role in cell survival and proliferation by using loss of function and gain of function assays in cellular models.
As nucleoporins feed multiple arms of the cellular physiology, it is challenging to identify what particular altered functions might lead to tumorigenesis. Finding the downstream effects is important as it provides an opportunity to target the specific pathway for therapeutic purpose. e.g. Importazole was used to reduce tumorogenic potential in POM121 overexpressed cells as it was mediated through interaction with importin b [80]. However, individual Nups might have role in the disease progression that is independent of nuclear transport functions, e.g the role of Nup62 in regulating gastric cancer metastasis by regulating Wnt/b-Catenin and TGF-b signaling pathways [81]. In such circumstances, individual Nups and/or the downstream pathways that are altered might have to be unraveled for eventual targeting.
Although inhibitors for nuclear transport receptors are relatively well studied in cancer, specific inhibitors for Nups are equally promising in various malignancies considering their massive deregulation, as exemplified by the current study in breast cancer. The mutations in Nup genes were represented in 50% of the breast cancer patients in addition to deregulated expression representing an important vulnerable point. The repertoire of small molecules perturbing the nuclear pore permeability by disrupting FG interactions is limited to only trans1-2 cyclohexanediol (CHD) and Pitstop2 [82, 83]. Despite their ability to compromise the NPC permeability barrier effectively, their systemic effects on the NPCs may hinder their use in therapeutics. Design and synthesis of small molecules targeting specific components of NPCs remains an arduous task, albeit with potential to pave way for novel therapeutic strategies in breast cancer.
Authors' Contribution
R.M. and N.Y. conceived and designed the study, interpreted data, and finalized the manuscript;
R.M., M. N., and Z. A. conducted Bioinformatics analysis;
R.M., K.J., M.N., and N.Y. were involved in manuscript writing;
All authors read and approved the manuscript.
The authors declare no competing interests.
Acknowledgment
A grant from Alfaisal University (IRG 20410) to RM is thankfully acknowledged.
References
- Paci G, Caria J, Lemke EA. Cargo transport through the nuclear pore complex at a glance. Journal of cell science, (2021); 134(2): jcs247874.
- Fernandez-Martinez J, Rout MP. One ring to rule them all? Structural and functional diversity in the nuclear pore complex. Trends in Biochemical Sciences, (2021); 46(7): 595-607.
- Hurt E, Beck M. Towards understanding nuclear pore complex architecture and dynamics in the age of integrative structural analysis. Current opinion in cell biology, (2015); 3431-38.
- Sakiyama Y, Panatala R, Lim RY. Structural dynamics of the nuclear pore complex; 2017. Elsevier. pp. 27-33.
- Strawn LA, Shen T, Shulga N, Goldfarb DS, Wente SR. Minimal nuclear pore complexes define FG repeat domains essential for transport. Nature cell biology, (2004); 6(3): 197-206.
- Cook A, Bono F, Jinek M, Conti E. Structural biology of nucleocytoplasmic transport. Annual review of biochemistry, (2007); 76(1): 647-671.
- Güttler T, Görlich D. Ran‐dependent nuclear export mediators: a structural perspective. The EMBO journal, (2011); 30(17): 3457-3474.
- Rougemaille M, Dieppois G, Kisseleva-Romanova E, Gudipati RK, Lemoine S, et al. THO/Sub2p functions to coordinate 3′-end processing with gene-nuclear pore association. Cell, (2008); 135(2): 308-321.
- Taddei A, Van Houwe G, Hediger F, Kalck V, Cubizolles F, et al. Nuclear pore association confers optimal expression levels for an inducible yeast gene. Nature, (2006); 441(7094): 774-778.
- Light WH, Brickner DG, Brand VR, Brickner JH. Interaction of a DNA zip code with the nuclear pore complex promotes H2A. Z incorporation and INO1 transcriptional memory. Molecular cell, (2010); 40(1): 112-125.
- Brickner DG, Brickner JH. Interchromosomal clustering of active genes at the nuclear pore complex. Nucleus, (2012); 3(6): 487-492.
- Casolari JM, Brown CR, Komili S, West J, Hieronymus H, et al. Genome-wide localization of the nuclear transport machinery couples transcriptional status and nuclear organization. Cell, (2004); 117(4): 427-439.
- Brown CR, Silver PA. Transcriptional regulation at the nuclear pore complex. Current opinion in genetics & development, (2007); 17(2): 100-106.
- Vaquerizas JM, Suyama R, Kind J, Miura K, Luscombe NM, et al. Nuclear pore proteins nup153 and megator define transcriptionally active regions in the Drosophila genome. PLoS genetics, (2010); 6(2): e1000846.
- Kuhn TM, Capelson M. Nuclear pore proteins in regulation of chromatin state. Cells, (2019); 8(11): 1414.
- Ibarra A, Benner C, Tyagi S, Cool J, Hetzer MW. Nucleoporin-mediated regulation of cell identity genes. Genes & development, (2016); 30(20): 2253-2258.
- Sun J, Shi Y, Yildirim E. The nuclear pore complex in cell type-specific chromatin structure and gene regulation. Trends in Genetics, (2019); 35(8): 579-588.
- Burdine RD, Preston CC, Leonard RJ, Bradley TA, Faustino RS. Nucleoporins in cardiovascular disease. Journal of molecular and cellular cardiology, (2020); 14143-52.
- Oka M, Mura S, Otani M, Miyamoto Y, Nogami J, et al. Chromatin-bound CRM1 recruits SET-Nup214 and NPM1c onto HOX clusters causing aberrant HOX expression in leukemia cells. Elife, (2019); 8.
- Fahrenkrog B. Nucleoporin gene fusions and hematopoietic malignancies. New Journal of Science, (2014); 2014.
- Nofrini V, Di Giacomo D, Mecucci C. Nucleoporin genes in human diseases. European Journal of Human Genetics, (2016); 24(10): 1388-1395.
- Chow K-H, Factor RE, Ullman KS. The nuclear envelope environment and its cancer connections. Nature Reviews Cancer, (2012); 12(3): 196-209.
- Simon DN, Rout MP. Cancer and the nuclear pore complex. Cancer Biology and the Nuclear Envelope, (2014); 285-307.
- Rodriguez-Bravo V, Pippa R, Song W-M, Carceles-Cordon M, Dominguez-Andres A, et al. Nuclear pores promote lethal prostate cancer by increasing POM121-driven E2F1, MYC, and AR nuclear import. Cell, (2018); 174(5): 1200-1215. e1220.
- Franks TM, McCloskey A, Shokhirev MN, Benner C, Rathore A, et al. Nup98 recruits the Wdr82–Set1A/COMPASS complex to promoters to regulate H3K4 trimethylation in hematopoietic progenitor cells. Genes & development, (2017); 31(22): 2222-2234.
- Agudo D, Gómez‐Esquer F, Martínez‐Arribas F, Núñez‐Villar MJ, Pollán M, et al. Nup88 mRNA overexpression is associated with high aggressiveness of breast cancer. International journal of cancer, (2004); 109(5): 717-720.
- Bersini S, Lytle NK, Schulte R, Huang L, Wahl GM, et al. Nup93 regulates breast tumor growth by modulating cell proliferation and actin cytoskeleton remodeling. Life science alliance, (2020); 3(1).
- Cerami E, Gao J, Dogrusoz U, Gross BE, Sumer SO, et al. The cBio cancer genomics portal: an open platform for exploring multidimensional cancer genomics data. Cancer discovery, (2012); 2(5): 401-404.
- Gao J, Aksoy BA, Dogrusoz U, Dresdner G, Gross B, et al. Integrative analysis of complex cancer genomics and clinical profiles using the cBioPortal. Science signaling, (2013); 6(269): pl1-pl1.
- Khan AU, Qu R, Ouyang J, Dai J. Role of nucleoporins and transport receptors in cell differentiation. Frontiers in Physiology, (2020); 11239.
- Goldstein AL, Snay CA, Heath CV, Cole C. Pleiotropic nuclear defects associated with a conditional allele of the novel nucleoporin Rat9p/Nup85p. Molecular biology of the cell, (1996); 7(6): 917-934.
- Vasu S, Shah S, Orjalo A, Park M, Fischer WH, et al. Novel vertebrate nucleoporins Nup133 and Nup160 play a role in mRNA export. The Journal of cell biology, (2001); 155(3): 339-354.
- Boehmer T, Enninga J, Dales S, Blobel G, Zhong H. Depletion of a single nucleoporin, Nup107, prevents the assembly of a subset of nucleoporins into the nuclear pore complex. Proceedings of the National Academy of Sciences, (2003); 100(3): 981-985.
- Moroianu J, Hijikata M, Blobel G, Radu A. Mammalian karyopherin alpha 1 beta and alpha 2 beta heterodimers: alpha 1 or alpha 2 subunit binds nuclear localization signal and beta subunit interacts with peptide repeat-containing nucleoporins. Proceedings of the National Academy of Sciences, (1995); 92(14): 6532-6536.
- Powers MA, Forbes DJ, Dahlberg JE, Lund E. The vertebrate GLFG nucleoporin, Nup98, is an essential component of multiple RNA export pathways. The Journal of cell biology, (1997); 136(2): 241-250.
- Stagg SM, LaPointe P, Razvi A, Gürkan C, Potter CS, et al. Structural basis for cargo regulation of COPII coat assembly. Cell, (2008); 134(3): 474-484.
- Loïodice I, Alves A, Rabut G, Van Overbeek M, Ellenberg J, et al. The entire Nup107-160 complex, including three new members, is targeted as one entity to kinetochores in mitosis. Molecular biology of the cell, (2004); 15(7): 3333-3344.
- Cronshaw JM, Krutchinsky AN, Zhang W, Chait BT, Matunis MJ. Proteomic analysis of the mammalian nuclear pore complex. The Journal of cell biology, (2002); 158(5): 915-927.
- Rasala BA, Orjalo AV, Shen Z, Briggs S, Forbes DJ. ELYS is a dual nucleoporin/kinetochore protein required for nuclear pore assembly and proper cell division. Proceedings of the National Academy of Sciences, (2006); 103(47): 17801-17806.
- Van Laer L, Van Camp G, Van Zuijlen D, Green E, Verstreken M, et al. Refined mapping of a gene for autosomal dominant progressive sensorineural hearing loss (DFNA5) to a 2-cM region, and exclusion of a candidate gene that is expressed in the cochlea. European Journal of Human Genetics, (1997); 5397-405.
- Beddow AL, Richards SA, Orem NR, Macara IG. The Ran/TC4 GTPase-binding domain: identification by expression cloning and characterization of a conserved sequence motif. Proceedings of the National Academy of Sciences, (1995); 92(8): 3328-3332.
- Kraemer D, Wozniak RW, Blobel G, Radu A. The human CAN protein, a putative oncogene product associated with myeloid leukemogenesis, is a nuclear pore complex protein that faces the cytoplasm. Proceedings of the National Academy of Sciences, (1994); 91(4): 1519-1523.
- Fornerod M, Van Deursen J, Van Baal S, Reynolds A, Davis D, et al. The human homologue of yeast CRM1 is in a dynamic subcomplex with CAN/Nup214 and a novel nuclear pore component Nup88. The EMBO journal, (1997); 16(4): 807-816.
- Tullio-Pelet A, Salomon R, Hadj-Rabia S, Mugnier C, de Laet M-H, et al. Mutant WD-repeat protein in triple-A syndrome. Nature genetics, (2000); 26(3): 332-335.
- Watkins JL, Murphy R, Emtage JL, Wente SR. The human homologue of Saccharomyces cerevisiae Gle1p is required for poly (A)+ RNA export. Proceedings of the National Academy of Sciences, (1998); 95(12): 6779-6784.
- Carmo-Fonseca M, Kern H, Hurt E. Human nucleoporin p62 and the essential yeast nuclear pore protein NSP1 show sequence homology and a similar domain organization. European journal of cell biology, (1991); 55(1): 17-30.
- Team* MGCP. Generation and initial analysis of more than 15,000 full-length human and mouse cDNA sequences. Proceedings of the National Academy of Sciences, (2002); 99(26): 16899-16903.
- Bharathi A, Ghosh A, Whalen WA, Yoon JH, Pu R, et al. The human RAE1 gene is a functional homologue of Schizosaccharomyces pombe rae1 gene involved in nuclear export of Poly (A)+ RNA. Gene, (1997); 198(1-2): 251-258.
- Grandi P, Dang T, Pané N, Shevchenko A, Mann M, et al. Nup93, a vertebrate homologue of yeast Nic96p, forms a complex with a novel 205-kDa protein and is required for correct nuclear pore assembly. Molecular biology of the cell, (1997); 8(10): 2017-2038.
- Miller BR, Powers M, Park M, Fischer W, Forbes DJ. Identification of a new vertebrate nucleoporin, Nup188, with the use of a novel organelle trap assay. Molecular biology of the cell, (2000); 11(10): 3381-3396.
- Nagase T, Seki N, Ishikawa K-i, Ohira M, Kawarabayasi Y, et al. Prediction of the coding sequences of unidentified human genes. VI. The coding sequences of 80 new genes (KIAA0201-KIAA0280) deduced by analysis of cDNA clones from cell line KG-1 and brain. DNA Research, (1996); 3(5): 321-329.
- Handa N, Kukimoto-Niino M, Akasaka R, Kishishita S, Murayama K, et al. The crystal structure of mouse Nup35 reveals atypical RNP motifs and novel homodimerization of the RRM domain. Journal of molecular biology, (2006); 363(1): 114-124.
- Zhang X, Yang H, Corydon MJ, Zhang X, Pedersen S, et al. Localization of a human nucleoporin 155 gene (NUP155) to the 5p13 region and cloning of its cDNA. Genomics, (1999); 57(1): 144-151.
- Krull S, Thyberg J, Bjorkroth B, Rackwitz H-R, Cordes VC. Nucleoporins as components of the nuclear pore complex core structure and Tpr as the architectural element of the nuclear basket. Molecular biology of the cell, (2004); 15(9): 4261-4277.
- McMorrow I, Bastos R, Horton H, Burke B. Sequence analysis of a cDNA encoding a human nuclear pore complex protein, hnup153. Biochimica et Biophysica Acta (BBA)-Gene Structure and Expression, (1994); 1217(2): 219-223.
- Trichet V, Shkolny D, Dunham I, Beare D, McDermid H. Mapping and complex expression pattern of the human NPAP60L nucleoporin gene. Cytogenetic and Genome Research, (1999); 85(3-4): 221-226.
- Trichet V, Shkolny D, Dunham I, Beare D, McDermid H. Mapping and complex expression pattern of the human NPAP60L nucleoporin gene. Cytogenetic and Genome Research, (1999); 85(3-4): 221-226.
- Schirmer EC, Florens L, Guan T, Yates III JR, Gerace L. Nuclear membrane proteins with potential disease links found by subtractive proteomics. Science, (2003); 301(5638): 1380-1382.
- Greber UF, Senior A, Gerace L. A major glycoprotein of the nuclear pore complex is a membrane‐spanning polypeptide with a large lumenal domain and a small cytoplasmic tail. The EMBO journal, (1990); 9(5): 1495-1502.
- Gusev FE, Reshetov DA, Mitchell AC, Andreeva TV, Dincer A, et al. Epigenetic‐genetic chromatin footprinting identifies novel and subject‐specific genes active in prefrontal cortex neurons. The FASEB Journal, (2019); 33(7): 8161-8173.
- Hallberg E, Wozniak RW, Blobel G. An integral membrane protein of the pore membrane domain of the nuclear envelope contains a nucleoporin-like region. The Journal of cell biology, (1993); 122(3): 513-521.
- Mungall AJ, Palmer S, Sims S, Edwards C, Ashurst J, et al. The DNA sequence and analysis of human chromosome 6. Nature, (2003); 425(6960): 805-811.
- Ota T, Suzuki Y, Nishikawa T, Otsuki T, Sugiyama T, et al. Complete sequencing and characterization of 21,243 full-length human cDNAs. Nature genetics, (2004); 36(1): 40-45.
- Funakoshi T, Maeshima K, Yahata K, Sugano S, Imamoto F, et al. Two distinct human POM121 genes: requirement for the formation of nuclear pore complexes. FEBS letters, (2007); 581(25): 4910-4916.
- Nagase T, Ishikawa K-i, Suyama M, Kikuno R, Miyajima N, et al. Prediction of the coding sequences of unidentified human genes. XI. The complete sequences of 100 new cDNA clones from brain which code for large proteins in vitro. DNA research, (1998); 5(5): 277-286.
- Hu T, Guan T, Gerace L. Molecular and functional characterization of the p62 complex, an assembly of nuclear pore complex glycoproteins. The Journal of cell biology, (1996); 134(3): 589-601.
- Schmitt C, von Kobbe C, Bachi A, Pante N, Rodrigues JP, et al. Dbp5, a DEAD-box protein required for mRNA export, is recruited to the cytoplasmic fibrils of nuclear pore complex via a conserved interaction with CAN/Nup159p. The EMBO journal, (1999); 18(15): 4332-4347.
- Goldman MJ, Craft B, Hastie M, Repečka K, McDade F, et al. Visualizing and interpreting cancer genomics data via the Xena platform. Nature biotechnology, (2020); 38(6): 675-678.
- Lonsdale J, Thomas J, Salvatore M, Phillips R, Lo E, et al. The genotype-tissue expression (GTEx) project. Nature genetics, (2013); 45(6): 580-585.
- Györffy B, Lanczky A, Eklund AC, Denkert C, Budczies J, et al. An online survival analysis tool to rapidly assess the effect of 22,277 genes on breast cancer prognosis using microarray data of 1,809 patients. Breast cancer research and treatment, (2010); 123(3): 725-731.
- Mehmood R, Jibiki K, Shibazaki N, Yasuhara N. Molecular profiling of nucleocytoplasmic transport factor genes in breast cancer. Heliyon, (2021); 7(1): e06039.
- Stanley GJ, Fassati A, Hoogenboom BW. Biomechanics of the transport barrier in the nuclear pore complex; 2017. Elsevier. pp. 42-51.
- Tian C, Zhou S, Yi C. High NUP43 expression might independently predict poor overall survival in luminal A and in HER2+ breast cancer. Future Oncology, (2018); 14(15): 1431-1442.
- Mullan PB, Bingham V, Haddock P, Irwin GW, Kay E, et al. NUP98–a novel predictor of response to anthracycline-based chemotherapy in triple negative breast cancer. BMC cancer, (2019); 19(1): 1-13.
- Michmerhuizen NL, Klco JM, Mullighan CG. Mechanistic insights and potential therapeutic approaches for NUP98-rearranged hematologic malignancies. Blood, (2020); 136(20): 2275-2289.
- Aramburu IV, Lemke EA. Floppy but not sloppy: Interaction mechanism of FG-nucleoporins and nuclear transport receptors; 2017. Elsevier. pp. 34-41.
- Holzer K, Ori A, Cooke A, Dauch D, Drucker E, et al. Nucleoporin Nup155 is part of the p53 network in liver cancer. Nature communications, (2019); 10(1): 1-13.
- Kehat I, Accornero F, Aronow BJ, Molkentin JD. Modulation of chromatin position and gene expression by HDAC4 interaction with nucleoporins. Journal of Cell Biology, (2011); 193(1): 21-29.
- Preston CC, Storm EC, Burdine RD, Bradley TA, Uttecht AD, et al. Nucleoporin insufficiency disrupts a pluripotent regulatory circuit in a pro-arrhythmogenic stem cell line. Scientific reports, (2019); 9(1): 1-10.
- Enninga J, Levay A, Fontoura BM. Sec13 shuttles between the nucleus and the cytoplasm and stably interacts with Nup96 at the nuclear pore complex. Molecular and cellular biology, (2003); 23(20): 7271-7284.
- Lim KS, Wong RW. Targeting Nucleoporin POM121-Importin β Axis in Prostate Cancer. Cell chemical biology, (2018); 25(9): 1056-1058.
- Wang H, Lin Y, Jin J, Shen H, Dai C. Nuclear Pore Complex 62 Promotes Metastasis of Gastric Cancer by Regulating Wnt/β-Catenin and TGF-β Signaling Pathways. Journal of Environmental Pathology, Toxicology and Oncology, (2021); 40(2).
- Ribbeck K, Görlich D. The permeability barrier of nuclear pore complexes appears to operate via hydrophobic exclusion. The EMBO journal, (2002); 21(11): 2664-2671.
- Liashkovich I, Pasrednik D, Prystopiuk V, Rosso G, Oberleithner H, et al. Clathrin inhibitor Pitstop-2 disrupts the nuclear pore complex permeability barrier. Scientific reports, (2015); 5(1): 1-9.
- Souquet B, Freed E, Berto A, Andric V, Audugé N, et al. Nup133 is required for proper nuclear pore basket assembly and dynamics in embryonic stem cells. Cell reports, (2018); 23(8): 2443-2454.
- Sajidah ES, Lim K, Wong RW. How SARS-CoV-2 and other viruses build an invasion route to hijack the host nucleocytoplasmic trafficking system. Cells, (2021); 10(6): 1424.
- Shen Q, Wang YE, Palazzo AF. Crosstalk between nucleocytoplasmic trafficking and the innate immune response to viral infection. Journal of Biological Chemistry, (2021); 297(1).
- Szklarczyk D, Franceschini A, Kuhn M, Simonovic M, Roth A, et al. The STRING database in 2011: functional interaction networks of proteins, globally integrated and scored. Nucleic acids research, (2010); 39(suppl_1): D561-D568.
- Holzer G, Antonin W. Breaking the Y. PLoS Genetics, (2019); 15(5): e1008109.
- Souquet B, Freed E, Berto A, Andric V, Audugé N, et al. Nup133 is required for proper nuclear pore basket assembly and dynamics in embryonic stem cells. Cell reports, (2018); 23(8): 2443-2454.
- Jevtić P, Schibler AC, Wesley CC, Pegoraro G, Misteli T, et al. The nucleoporin ELYS regulates nuclear size by controlling NPC number and nuclear import capacity. EMBO reports, (2019); 20(6): e47283.
- Nakano H, Wang W, Hashizume C, Funasaka T, Sato H, et al. Unexpected role of nucleoporins in coordination of cell cycle progression. Cell Cycle, (2011); 10(3): 425-433.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0