

Full Length Research Article
Indigenous Curd as a Functional Food: A Source of Potential Pathogenic Bacterial Control
Shazia Yaqub1*, Mian Anjum Murtaza1, Shinawar Waseem Ali2, Sehrish Mushtaq2, Sohaib Afzaal2, Abdul Munim Farooq3, Ghulam Mustafa4
Adv. life sci., vol. 9, no. 1, pp. 24-31, May 2022
*- Corresponding Author: Shazia Yaqub (Email: shaziaft743@gmail.com)
Authors' Affiliations
2. Institute of Agriculture Sciences, University of the Punjab, Lahore – Pakistan
3. National Centre of Excellence for Molecular biology, Lahore – Pakistan
4. Institute of Allied Health sciences, Lahore – Pakistan
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Dahi is a functional food that is largely consumed in our community. The current study was designed to explore the efficacy of antibacterial agents that are present in the local indigenous curd of Lahore, Pakistan.
Method: Dahi was collected from various areas in Lahore. Bacterial cultures were isolated from this local curd and identified using a molecular technique that used 16S rRNA gene primers in PCR and sequencing. The antibacterial assay was carried out using the agar well diffusion method. Out of all the characterized bacteria isolate P26 displayed the best results for the control of pathogenic bacteria. Bacteriocin from isolate P26 was partially purified and its antibacterial assay was performed against sixteen indicator organisms and kanamycin was used as a standard for this experiment. Characterization of bacteriocin was performed at various temperatures, pH, and treating with enzymes.
Result: After the molecular characterization through 16S rRNA, Strain P26 was characterized as Bacillus vallismortis. A protease-sensitive antimicrobial substance was produced by Bacterial strain P26 (Bacillus vallismortis), isolated from local curd (Dahi). This isolate has a broad-spectrum inhibitory activity that can inhibit the growth of both food spoilage and pathogenic bacteria. According to results It was clear that protease-sensitive antimicrobial substance produced from strain P26 is more effective as compared to the antibiotic kanamycin. Maximum activity was shown against Staphylococcus species, a pathogen that is a common cause of gastroenteritis worldwide. This substance was active at 100 ºC for 30 minutes and pH (2-9). The activity was lost after treatment with proteases, so it is classified as bacteriocin.
Conclusion: Bacteriocin P26 is a more effective antibacterial agent than kanamycin, and it can be produced cheaply using local cardkey.
Keywords: Protease; Bacteriocin; Staphylococcus carnosus; Kanamycin
Introduction![]()
Dahi (Curd) is a fermented milk product very common among the Pakistani community due to its taste and nutritional value. Dahi (curd) is produced under uncontrolled milk fermentation conditions at both commercial and domestic levels, as milk is inoculated with one-day-old curd (Dahi) [1, 2]. Dahi is categorized as a functional food as its health benefits and therapeutic effects are strongly addressed. It is effective against a different disease that affects the gastrointestinal tract [3]. Many antibacterial chemicals are produced during fermentation, and the pH of milk is reduced, making it an unsuitable habitat for certain bacteria, so reducing the growth of undesirable pathogens. [4]. Various compounds that are by-products of fermentation and effective against other microbes include organic acids, hydrogen peroxide, and bacteriocins produced by LAB (Lactic acid bacteria) during fermentation [5].
Bacteriocins are ribosomally produced antimicrobial compounds, protein in nature, and are produced by a wide range of bacteria [6]. Since bacteriocins are natural antimicrobial agents so these bacteria could be the best possible replacement of chemical preservatives and can act as a strong linkage between food and pharmaceutical industries [7]. More effective responses can be obtained by combinations made of one bacteriocin with another, or bacteriocins with other natural antimicrobial compounds [8].
As we live in an era where multi-drug resistance (MSD) is on the rise, one of the key goals of the pharmaceutical business is to locate natural antibiotics, particularly from the local environment, in order to control harmful bacteria and, ultimately, to reduce disease prevalence. As a result of the foregoing facts, the current study is aimed to examine different antibacterial agents derived from the local indigenous curd of Lahore, Pakistan.
Methods![]()
Dahi was collected from different locations of Lahore and seventy (70) bacteria were isolated and purified.
Antibacterial Potential Assay:
Indicator organisms were taken from First culture bank of Pakistan, Faculty of Agricultural Sciences, University of the Punjab. These seventy bacterial pure cultures were further screened for antibacterial potential against sixteen indicator bacteria named as (Listeria monocytogenes, Escherichia coli, Salmonella typhi, Salmonella paratyphi, Yersinia pseudotuberculosis, Yersinia enterocolitica, Klebsiella pneumoniae, Klebsiella pneumoniae ssp. rhinosclerosis, Clostridium difficille, Bacillus subtilis, Staphylococcus caprae, and Staphylococcus carnosus, Micrococcus roseus, Micrococcus luteus, Shigella dysenteriae, and Pseudomonas. All indicator organisms were grown in tubes containing LB broth medium by inoculating loop full sample and incubating at 37 °C for 24 hrs.
The purified bacterial isolates were grown in respective MRS medium by inoculating loop full of pure culture in tubes containing 7 mL MRS and M17 broth at 37 °C for 24 hrs. It was centrifuged, and the pellet was separated from the supernatant. The pellet was separated from the supernatant after centrifugation. The pellet was used to extract DNA.
Agar well diffusion method was adopted to analyze the antibacterial spectrum of isolated bacteria P26 against indicator organisms [9]. Wells were made in an agar plate with a sterile borer of 5mm. Then 100 microliter of bacterial supernatant was filled in it. It was given time for proper absorption before incubation. The zone of inhibition was measured in mm. Kanamycin (100 mg/mL) was used as a standard. The zone of inhibition produced by antibiotic kanamycin and bacterial culture was compared.
Isolation of Bacteriocin:
Cell-free culture supernatant fluid (Cfcsf) for bacteriocin assay:
The bacterial isolate P26 was further screened for bacteriocin activity. The bacterial strain was grown in broth media and their supernatant was separated by centrifugation.
pH neutralization:
The pH of Cfcsf was set by using acid standard solutions of (Hydrochloric acid) and base (Sodium hydroxide) for the exclusion of the antimicrobial effect due to acids (organic) released by bacteria in the medium. The antagonistic action was tested by adopting the agar well diffusion method [10].
Treatment with catalase:
To check the hydrogen peroxide effect on antagonistic activity, the catalase enzyme 10 mg/mL was poured into it. It was held (37°C, 2 hrs.). Afterward, it was heated at 80 °C for enzyme inactivation and inhibitory activity as checked against indicator bacteria [10].
Confirmation of bacteriocin:
To confirm the presence of bacteriocin, a 1 mL sample was treated with filter-sterilized protease enzyme solutions for 2 h at 37 °C, trypsin proteinase Kin 20 mM Tris-HCl, pH 7.0 [11].
Antibacterial activity of bacteriocin P26:
After confirmation of bacteriocin, its antibacterial assay was performed against sixteen indicator organisms. Kanamycin 10mg/mL was used as standard.
Characterization of Isolate P26
Phenotypical characterization:
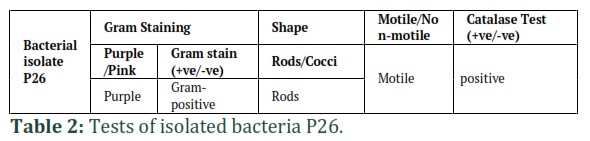
Isolate P26 was identified morphologically based on colony shape, color, texture, margins, opacity, and elevation. Bacterial isolate (P26) was gram stained and their morphological features, movement (motility) were observed under a light microscope. Then different biochemical and enzymatic (Catalase, Sugar fermentation) tests were performed for their identification following Bergey's manual of systematic bacteriology.
Molecular characterization
Genomic DNA isolation:
Genomic DNA was extracted by the following method of Cardinal and others [12] with some amendments. Activated MRS cultures (10mL) were centrifuged (12000 rpm, 10 min.), the supernatant was discarded, and the pellet was washed with 1ml EDTA-NaCl solution then re-suspended in 560 µL 1xTE buffer (pH 8). 3µL of 1mg/mL proteinase K, 3µL lysozyme (20mg/ml), 10 % SDS 20 µL were added and after gentle mixing 100 µL of 5M NaCl solution and 80 µL, CTAB/NaCl solution was added at kept at 37°C for 30 minutes. Then 700 µL chloroform/isoamyl was added before centrifugation. The upper phase was shifted into a new Eppendorf tube. It was extracted twice with 700 µL chloroform/isoamyl alcohol. DNA was precipitated by the addition of sodium acetate 20 µL and absolute ethanol 500 µL. It was mixed and kept overnight at -20 ⁰C. By centrifugation (12000rpm, 15 min), a pellet of DNA was formed. Then it was washed with 200 µL of 70% ethanol. DNA was again centrifuged after washing. Ethanol was removed and dried at room temperature. Then it was mixed with distilled water. 100µg/mL RNase (1µL/20µL DNA) was added before incubation at 37 °C for 30 minutes. Then washed with alcohol (chloroform/isoamyl 24/1, v/v). The upper phase was shifted into a new Eppendorf tube. Absolute ethanol 50 µL and sodium acetate 3 µL was added and kept overnight at -20 °C overnight and then centrifuged for pellet formation. Pellet was dried. And 25 µL of distilled water was added and mixed. Then the band was observed under UV transilluminator by conducting electrophoresis (agarose gel).
Polymerase chain reaction:
PCR-based amplification of isolated bacterial DNA was achieved through universal 16S ribosomal RNA primers obtained from Macrogen, (South Korea). 27F: 5’ AGA GTT TGA TCM TGG CTC AG-3” and 1492R: 5’ CGG TTA CCT TGT TAC GAC TT-3’. The annealing temperature was calculated according to primer sequences and Taq polymerase to be used. A thermal cycler was used for amplification. The reaction mixture contained (30 µL) contained Taq DNA polymerase buffer (contain MgCl2 and dNTPs) 15μL, Oligo forward (10 picomole/μL) 3μL, Oligo reverse (10 picomole/μl) 3μL, Phire Taq 0.6 µL, sterile deionized water 7.4μL. The DNA fragments were amplified as follows: initial denaturation at 98°C for 5 min followed by 35 cycles of denaturation at 98°C for 20 sec, annealing at 72°C for 20 sec, elongation at 72°C for 1 min, and final elongation at 72°C for 5 min. The products were stored at 4 °C until analysis. Aliquots (5 µL) of the amplified products were visualized by agarose gel electrophoresis (1%) using a DNA marker (Gene ruler 1Kb DNA) under UV Trans illuminator. Sequencing results obtained were matched and compared with primer walking and the gaps were filled. The obtained sequences were compared with the NCBI Gene Bank database by Basic local alignment search tool (BLAST) analysis. The phylogenetic tree was constructed using MEGA 6 software with a maximum likelihood method based on 16S rRNA gene fragments and was evaluated by the bootstrap method with 1000 replicates.
Partial purification of bacteriocin:
Bacteriocin (P26) was partially purified with ammonium sulphate by adopting the method of [13] 2010 with some modifications.
Ammonium sulphate precipitation:
Sterilized broth media was inoculated with bacterial culture and incubated under proper conditions. By centrifugation, pellet and supernatant were separated. The supernatant was heated at 80°C for 30 minutes. Then filter through a 0.22um syringe filter to remove any remaining cell. Its pH was adjusted at 6.5 with 2N NaOH. Then ammonium sulphate salt was added up to 40%, 60%, 80% saturation and it was continuously stirred for 15 hours. The solution was stirred in a magnetic stirrer using magnetic beads. This was kept at 4 ºC overnight. Then it was refrigerated centrifuged at 10,000 rpm for 30 min for protein precipitation. The supernatant was discarded, and precipitates collected were dissolved in 50mM tris buffer (pH 6.8).
Brad ford test:
Turn on the spectrophotometer and let it warm up for 15 minutes. It was calibrated at 595nm. In Eppendorf, 20L of each sample was accurately pipetted out, and 50 L of 1M NaOH was added. 1 mL of Coomassie brilliant blue R-250 (Bio-Rad) was gently mixed in. It was allowed to sit at room temperature for 15 minutes. The sample was placed in 1mL disposable polystyrene cuvettes, and absorbance at 595nm was measured. Using a known concentration of bovine serum albumin, a standard curve was created. The sample's concentration was determined by comparing it to the standard curve [14].
Bacteriocin Characterization
The antagonistic activity in arbitrary unit/ml (AU/mL) was calculated as a measure of bacteriocin production.
AU/ml = Diameter of zone of clearance (mm) x1000 / Volume taken in well (µL)
Activity is expressed as the percentage of residual activity (AU/ml) measured against indicator organisms by agar-well diffusion assay
Quantity of supernatant filled in wells =100µL
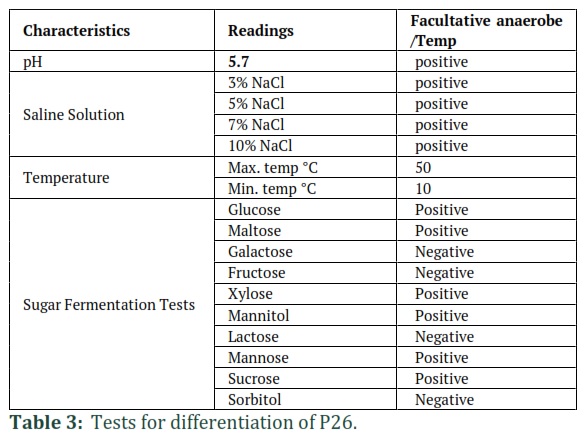
For bacteriocin characterization, the culture strain was cultivated in MRS broth (24-48 h at 37°C). It was centrifuged and Cfcsf was separated from the pellet. It was considered a crude bacteriocin [14]. Partially Purified (crude) bacteriocin was characterized at various temperatures (50 °C, 70 °C and 100 °C for 30 minutes, and at 121°C for 5 min), pH ranges (3·0, 4·0, 5·0, 6·0, 7·0, 8·0, 9·0 or 10·0) and assayed for bacteriocins activity [10]. The crude bacteriocin was also characterized by assessing sensitivity to proteolytic and non-proteolytic enzymes.
Results
![]()
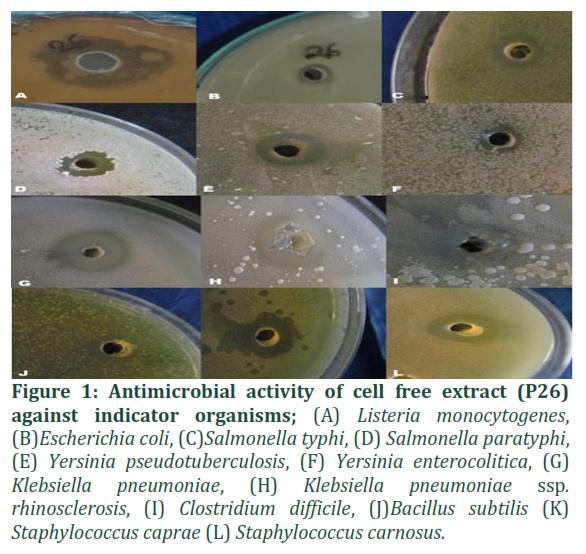
Antibacterial activity of bacterial strain P26 (Bacillus vallismortis).
The growth of twelve indicator species is inhibited by bacterial strain P26. It was discovered to be effective against Listeria monocytogenes, Escherichia coli, Salmonella typhi, Salmonella paratyphi, Yersinia pseudotuberculosis, Yersinia enterocolitica, Klebsiella pneumoniae, Klebsiella pneumoniae ssp. rhinosclerosis, Clostridium difficille, Bacillus subtilis, Sta (See Figure 1). Whereas, Micrococcus species, Shigella dysenteriae, and Pseudomonas spp. are not inhibited by it.
Bacteriocin activity assay:
pH adjustment and catalase treatment: After pH adjustment, the supernatant showed growth inhibition confirming that organic acids are not responsible for this inhibitory activity. Catalase-treated supernatant was tested for antagonistic activity against indicator bacteria by the agar well diffusion method. The results were the same confirming that this inhibition was not due to hydrogen peroxide produced by bacteria.
Confirmation of Bacteriocin Presence:
The supernatant was treated with protease enzyme and again check against indicator organisms. No inhibition was observed from any of the supernatants of 10 bacteria. This confirms that the compound that is responsible for inhibitory potential is a protein in nature. From this experiment, we named the supernatant Bacteriocin P26.
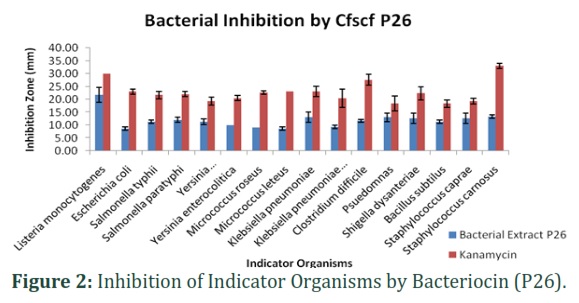
Antibacterial activity of bacteriocin P26: Bacteriocin P26 and kanamycin both inhibit the growth of all sixteen (16) indicator organisms. The inhibition zones formed by bacteriocin P26 were larger as compared to kanamycin.
Bacteriocin showed maximum inhibitory potential against Staphylococcus carnosus (33mm) as compared to kanamycin (13 mm). Minimum growth inhibition by bacterial extract P26 was observed against Pseudomonas (18.3mm) and Bacillus subtilis (18.3mm) as shown in Fig. 2
Phenotypical characterization of isolate P26:
Identification of isolate P26 was performed based on morphological traits of bacterial culture. Phenotypical characterization is shown in (Table 1 and Figure 3).-Results of the gram staining, biochemical and enzymatic tests performed for identification are shown in Table 2 and Table 3.
Molecular characterization of isolate P26:
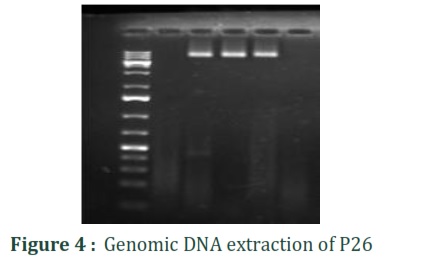
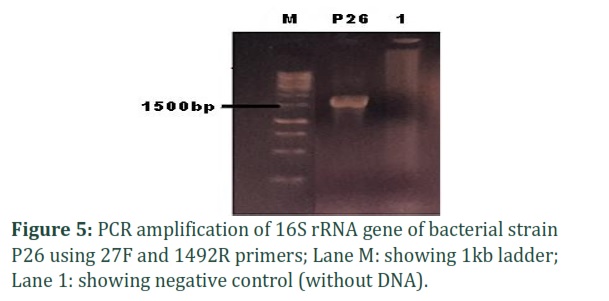
Bacterial strain P26 with potential antimicrobial activity was isolated from local indigenous curd collected from Lahore city, Punjab, Pakistan. The DNA was extracted, and agarose gel electrophoresis of DNA samples was performed to confirm it as shown in (Figure 4). The 16S rRNA gene was amplified using universal primers 27F /1492R as shown in (Figure 5).
Sequencing:
The PCR amplicon of the 16S rRNA gene of strain P26 was sequenced (Sanger Sequencing). This was found in continuous stretches of approximately 1500 base pairs (bp). The species was initially identified by using the BLAST program available at NCBI (http://www.ncbi.nlm.nih.gov/) showing greater than 97% similarity compared with the 16S rRNA sequences of type strains. The sequences were deposited in Gene Bank, assigning the accession number “MG733148 (P26)”.
Phylogenetic analysis:
The phylogenetic analysis including the representative strains and reference strains of species of isolate P26 was performed using MEGA 6 [16]. Nucleotide sequences were aligned and compared to other nucleotide sequences from GenBank using Clustal W and a phylogenetic tree was generated using the maximum likelihood method in MEGA software (Version 6.0; Biodesign Institute, Tempe, USA).
Analysis of the robustness of the trees was obtained by a bootstrap approach with 1000 replicates. The isolates and related reference strains were used to construct the phylogenetic trees. Accession numbers of the sequences included in the analysis are shown in Figure 6 available as supplementary material in NCBI gene bank Online. The phylogenetic trees were obtained to confirm the isolate. Isolate P26 closely resembles Bacillus vallismortis.
Characterization of Bacteriocin P26
The bacteriocine formed from P26 was characterized in three ways: chemically (actions of enzymes and catalase), physically (impact of heat and pH). Following partial purification and comparison with the standard curve, the protein content was calculated to be 52.52 ug/ml. The following section provides a detailed discussion of the characterisation of bacteriocine P26.
Chemical Characterization
Effects of enzymes:
The antagonistic activity was completely abolished by proteinase K, trypsin, and chymotrypsin treatment confirming its proteinaceous nature.
Effect of catalase:
Catalase-treated supernatants were tested for antagonistic activity against indicator bacteria by the agar well diffusion method. The results were the same confirming that catalase does not affect the bacteriocin activity
Physical Characterization
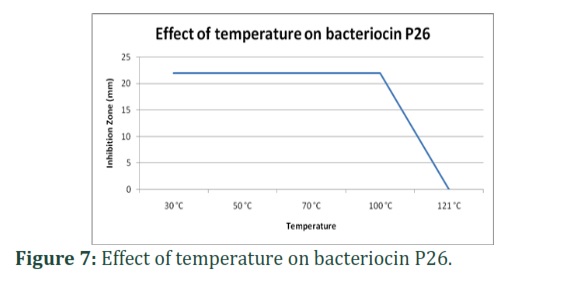
Effects of heat:
The activity was stable following temperature exposure up to 100°C for 30 minutes but lost completely at 121°C for 5 min as shown in Figure 7.
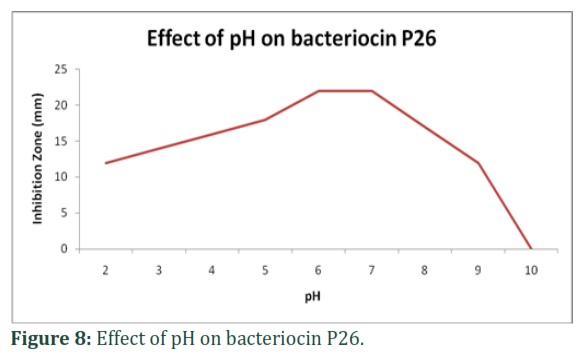
Effects of pH:
The cell-free supernatant was shown to retain the activity within the pH range of 2–9, and the optimum pH for the maximum activity was 6 and 7. Above pH 9, activity decrease to zero as shown in Figure8.
Figures & Tables


Discussion![]()
In the earlier period, the emerging problem of multidrug resistance due to excessive use or false use of antibiotics has imposed a threat of serious health issues. The only solution to this problem is to find new antimicrobial agents that are safe for health and the environment rather than chemicals. The current study suggests that bacterial Bacillus Vallismortis (bacteriocin P26) can be used as a potential antibacterial agent against human pathogenic bacteria. These can be used alone or in combination with other bacteria.
Bacillus Vallismortis was isolated from Dahi (curd) from different locations of Lahore after purification. Its antibacterial potential was assessed against sixteen indicator bacteria. It was found effective against twelve pathogens.
Bacteriocin P26 was purified by the ammonium sulphate precipitation method. After purification, it was characterized. The results revealed that bacteriocin of bacteria P26 inhibits the growth of sixteen test bacteria (both pathogenic and food spoilage). Bacteriocin P26 is more effective than the supernatant of Bacillus Vallismortis suggesting that there is some other unidentified reason due to which bacterial supernatant does not inhibit the growth of Micrococcus species, Pseudomonas and Shigella dysantrae. Both bacteriocin P26 and Kanamycin suppressed the indicator organisms, however as demonstrated above, bacteriocin P26 had a greater inhibitory capacity, implying that natural antimicrobials are more effective.
Several other bacteriocins are isolated, purified, and characterized previously as [17] isolated bacteria from traditional Bulgarian dairy products (home-made fermented milk yogurt for further isolation of bacterial strain having maximum antibacterial activity. In 2009, [18] from various food sources, isolated 601 Lactic acid bacteria (LAB). Among these 35 showed antibacterial activity against test organisms. This activity of the compound showing antibacterial activity was lost after being treated with proteolytic enzymes. This compound was found heat stable and active in pH (2-12) medium. [19] Naz and others isolated 300 bacterial strains from different ecological niches including food, water, soil, stools, and clinical samples. 54.3% of them were separated as bacteriocin producers. Among them bacteriocin from Pseudomonas aeruginosa was named as Pyocin was partially purified by ammonium sulphate precipitation (70%). But it was not protein in nature as don’t affect by proteinase enzymes. It was found heat-stable at 100 ˚C for 15 minutes and pH range of 4 to 10. Mahmood and others [20] isolated fourteen different strains of Lactobacillus bulgaricus from dahi. Amongst, a strain TLB06FT was found to have a wide array of antibacterial activities against Gram-positive and negative bacteria was selected for further characterization. Maximum production of bacteriocin at pH 5.5 and temperature of 30-37 ˚C.
Bacteriocin P26 was found to be highly stable under different physicochemical conditions. The unambiguous reason for the thermal stability of bacteriocins is still unknown however; there could be many factors responsible for providing stability to proteins at higher temperatures. The high concentration of proline residues, salt bridges, hydrogen bonds, and polar surface residues can reduce the risk of instability of any protein at elevated temperatures therefore similar factors might be involved in providing thermal stability to P26. These factors not only stabilize the proteins but are also responsible for regulating their kinetics at higher temperatures [21].
Keeping in view the present facts about Bacillus vallismortis and Bacteriocin P26 with reference to biochemical properties and antibacterial study, it is concluded that Bacteriacin P26 is a broad-spectrum antibacterial moiety that showed more inhibitory potential when compared to kanamycin. It is thermostable so can be applicable under harsh environmental conditions. Further research on the optimization of conditions for maximum production of bacteriocin P26 as its protein characterization will lead towards the development of an indigenous resource for the antibacterial agent to be used in the food as well as the pharmaceutical industry.
Author Contributions
Mian Anjum Murtaza: Designed the experiment
Shinawar Waseem Ali: Designed the study/ Reviewed the paper /Supervised the work
Shazia Yaqub: Collection of curd samples, isolation and phenotypic characterization of bacteria, Paper writing
Sehrish Mushtaq: Partial purification and characterization of bacteriocins, Paper writing
Sohaib Afzaal: Partial purification and characterization of bacteriocins, Paper writing
Abdul Munim Farooq: Molecular characterization of bacteria
Ghulam Mustafa: Optimization of PCR
The authors acknowledge the Higher Education Commission of Pakistan for funding the reported research work through the indigenous Ph.D. Fellowship Program.
References
- Agarwal, KN, Bhasin SK. Feasibility studies to control acute diarrhea in children by feeding fermented milk preparations Actimel and Indian Dahi. European Journal of Clinical Nutrition, (2003); 56(S4): S56.
- Saha SR, Islam MN, Shekh AL , Siddiki MSR. Dahi manufacturing from reconstituted skim milk partially replaced by coconut milk. Bangladesh Journal of Animal Sciences, (2014); 43(3): 218-223.
- Samanta A, Pradhan S, Mandal A, Patra A, Roy S, et al. Effect of starter culture on development of curd (dahi) and their antagonistic property against some enteric pathogen. Indian Journal of Microbiological Research, (2015); 2(1): 30-39.
- Conly JM and Johnston LB. Coming Full Circle: From Antibiotics to Probiotics and Prebiotics. Canadian Journal of Infectious Disease Medical Microbiology, (2004).,15:161-163.
- Abo-Amer, AE. Characterization of a bacteriocin-like inhibitory substance produced by Lactobacillus plantarum isolated from Egyptian home-made yogurt. Science Asia, (2007); 33: 313-319.
- Indira K, Jayalakshmi S, Gopalakrishnan A, Srinivasan M. Biopreservative potential of marine Lactobacillus spp. African Microbiological Research, (2011); 5(16): 2287-2296.
- Papagianni M, Anastasiadou S. Pediocins: The bacteriocins of Pediococci. Sources, production, properties and applications. Microbiological cell factor, (2009); 8(1): 3.
- Galvez A, Abriouel H, Lopez, RL, Omar NB. Bacteriocin-based strategies for food biopreservation. International Journal of food microbioly, (2007); 120(1-2): 51-70.
- Kang JH, Lee MS. Characterization of a bacteriocin produced by Enterococcus faecium GM‐1 isolated from an infant. Journal of Applied Microbioly, (2005); 98(5): 1169-1176.
- Ghalfi H, Thonart P, Benkerroum N. Inhibitory activity of Lactobacillus curvatus CWBI-B28 against Listeria monocytogenes and ST2-verotoxin producing Escherichia coli O157. Afrrican Journal of Biotechnology, (2006); 5(22): 1-10.
- Mahobiya D, Shrivastava R. Production, optimization and characterization of broad spectrum bacteriocins from Lactobacillus plantarum DP2 and Lactobacillus casai DD1. Research Journal of Pharmaceutical, Biological and Chemical Sciences, (2015); 6(2): 738-747.
- Cardinal MJ, Meghrous J, Lacroix C, Simard RE. Isolation of Lactococcus lactis strains producing inhibitory activity against Listeria. Food Biotechnology, (1997); 11(2): 129-146.
- Alam SI. Studies on Bacteriocins From Environmental Bacteria. PhD thesis. Deptt. of Food Tech., Fed. Urdu Univ. Arts Sci and Tech., Karachi.
- Green MR and Sambrook J. Molecular Cloning. A laboratory Manual. 4th Edition. Cold Spring Harbor Laboratory Press. NewYork.
- Zamfir M, Callewaert R, Cornea PC, Savu L, Vatafu I, et al. Purification and characterization of a bacteriocin produced by Lactobacillus acidophilus IBB 801. Journal of Applied Microbiology, (1999); 87(6): 923-93.
- Kumar S, Tsai CJ, Nussinov R. Factors enhancing protein thermostability. Protein Eng. (2000); 13:179–91.
- Simova ED, Beshkova DB and Dimitrov ZP. Characterization and antimicrobial spectrum of bacteriocins produced by lactic acid bacteria isolated from traditional Bulgarian dairy products. Journal of Applied Microbiology, (2009); 106(2): 92-701.
- Naz SA , Rasool SA. Isolation, production and characterization of bacteriocins produced by strains from indigenous environments. Pakistan Journal of Botany, (2013); 45: 261-267.
- Sezer and Guven A. Investigation of bacteriocin production capability of lactic acid bacteria isolated from foods. Kafkas University of Vet Fak Derg, (2009); 15(1): 45-50.
- Mahmood T, Masud T, Ali S, Abbasi KS, Liaquat M. Optimization and partial characterization of bacteriocin produced by Lactobacillus bulgaricus-TLBFT06 isolated from Dahi. Pakistan Journal Pharmaceutical Science, (2015); 28(2):549-555.
- Doyle MP, Beuchat LR., Montville, TJ. Food Microbiology: Fundamental and Frontiers, 2nd Edition, ASM Press Washington, D.C.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0

