

Full Length Research Article
Enhancing the Expression and Purification of IL-29: A study of autoinduction and one-step Purification Methods
Zia Ur Rahman1, Nadeem Ahmed1*, Numan Fazal1,2, Muhammad Islam Khan1, Mohsin Ahmed khan1, Saad Tahir1, Muhammad Akram1, Sajjad Ullah2, Ahmad Usman Zafar1
Adv. life sci., vol. 10, no. 1, pp. 122-128, March 2023
*– Corresponding Author: Nadeem Ahmed (Email: nadeem.ahmed@cemb.edu.pk)
Authors' Affiliations
2. University Institute of Medical Laboratory Technology, Faculty of Allied health sciences, University of Lahore – Pakistan
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Cytokines have long been viewed as a sign of hope due to their immunomodulatory and therapeutic characteristics. Developing simple, economical and readily scaled technologies to simplify their manufacturing is a critical challenge.
Method: In this study we have used a customized medium to automatically induce the expression of the IL-29 in E. coli expression system from the T7 promoter, allowing for higher yields as compared to the traditional technique of IPTG induction. Similarly, one-step purification method was employed to make the fermentation process cost-effective, along with enhancing its efficiency.
Results: From 1 L batches of IPTG-induced and autoinduced media, the harvested biomass was 11.8 g and 13.4 g, respectively and their corresponding IBs were 3.8 g and 4.8 g. Total protein purified from 1 L batch was 132 mg, at a concentration of 13 mg/mL, with an indicated high purity of 97%. IL-29 significantly decreased the metabolic activity of HepG2 cells. Specifically, 50% of the cells died at a concentration of 0.156 μg/mL, while 80% of the cells died at a concentration of 5 μg/mL.
Conclusion: This study presents an economical solution for producing and purifying IL-29 in E. coli, resulting in higher yields of biomass and IBs than the expensive traditional method. The purified protein was highly pure and showed immunomodulatory effects on HepG2 cells. These findings have important implications for developing simplified and scalable technologies for cytokine production with therapeutic potential.
Keywords: Escherichia coli; Cytokines; Interleukins; Interferons; Protein purification
Introduction![]()
Cytokines have always been regarded as a beacon of hope because of their immunomodulatory and therapeutic properties [1]. Type I interferon (IFN- α/β) has shown great potential in inhibition of tumor growth but the severe toxicity associated with it has diminished its use as a potential anticancer drug [2]. Since they are limited to cells of a certain lineage, the third type of IFN, which comprises FN-λ1 (IL-29), IFN-λ2 (IL28A), and IFN-λ3 (IL28B), shares the same biological features as type I IFNs and opens the door to assessing its therapeutic potential [3]. Many studies suggest that, in contrast to IFN-α therapy, IFN-λ1 (IL-29) therapy is less toxic and suitable for certain types of malignancies as not all cells are responsive to this cytokine [4-6]. IL-29 has a strong antineoplastic effect against the plethora of cancers such as lung cancer, skin cancers, oesophageal carcinomas, colorectal cancers, gastric cancer, hepatocellular carcinomas, pancreatic cancer, neuroendocrine tumor cells, and glioblastomas [7]. Similarly, IL-29 has a strong therapeutic potential in the treatment of asthma, as it reduces inflammatory response by decreasing IL-5, IL-13, and IL-19 expression levels and enhancing the Foxp3 T-regulatory cells [7-9]. Further, IL-29 has been effective against viruses like Hepatitis B Virus, Hepatitis C virus, Influenza A, and SARS-Corona viruses [10-14]. The studies suggested that IL-29 has potential as a therapeutic replacement of type I interferons in terms of asthma, tumors, and viral infections. Furthermore, as IL-29 targets only specific cells type that allows it to be used in treatments while reducing the side effects associated with Type 1 Interferon therapies [15]. Therapies of IL-29 could be particularly targeted toward the liver and epithelial tissues while eliminating side effects on the kidney, central nervous system, spleen, and endothelial cells, as observed with therapies of Type I interferon[16]. All of these properties of IL-29 produce a dire need to increase its production by using multiple techniques of genetic engineering for further studies to develop it as a new therapeutic, particularly for cancer and viral infections [7,17].
Due to its simplicity and low-cost Escherichia coli is the most attractive host to produce therapeutic proteins. Usually, in E. coli the expression of eukaryotic proteins is highly regulated by using an inducible promoter. Bacteriophage T7 RNA polymerase system is nowadays mostly used to check the expression of different proteins among various E. coli–based expression systems [18-20]. As this system operates through the Lac operon moreover, protein expression initiates after the induction of IPTG [21]. For commercial and economic purposes, it is crucial to produce recombinant protein to maximize product yields in a minimal timeframe while ensuring cost-effectiveness [22]. Recently, the autoinduction process has been used in the replacement of IPTG induction process for the manufacturing of recombinant proteins underneath the regulator of T7 promoter in E. coli. In the autoinduction method, an autoinducing media is used which contains glucose, lactose, glycerol, and essential nutrients to help cellular growth and increase the expression of the desired protein. The autoinducing media utilizes glucose in the initial stages of cell culture to ensure smooth cell growth, followed by lactose and glycerol to induce protein production in a Lac-operon regulated system [23]. By adding lactose during the early stage of media preparation, protein production can be automatically induced, eliminating the need for monitoring cellular growth and adding an inducer. This system is quite effective for achieving maximum protein yield by reducing the steps involved in culturing. In addition to all this, the yield of autoinduction media is relatively high as compared to those which conventionally use IPTG as an inducer [24]. This study aimed to evaluate, for the first time, the expression of recombinant human IL-29 in E. coli using an autoinduction procedure. Additionally, the study sought to perform a one-step ion-exchange purification of human recombinant IL-29 and assess its biological activity.
Methods![]()
IL-29 gene cloning in expression vector
The IL-29 gene was codon-optimized in the Genescript codon optimization programme and received with the BamHI and NdeI sites in the pUC19 vector from IDT USA. The restriction enzymes BamH1 and Nde1 were used to digest the pUC19-IL-29 vector. Gene digested was then subcloned into the expression vector pET28a (Novagen, USA). For expression studies, the confirmed expression construct pET28a-IL-29 was transformed into BL21-DE3 E. coli strain.
IL-29 expression in culture flasks
From the plate, different colonies, containing the IL-29 gene, were cultured in Luria Bertani (LB) broth with 50 mg/mL kanamycin. The culture was agitated all night in a shaking incubator at 250 rpm and 37 °C. The culture was diluted separately into 30 mL of autoinduction media and 30 mL of terrific broth in a 500 mL culture flask at a ratio of 1:100, and it was then cultured at 37 °C at 250 rpm until the OD600 reached 0.7-1.0. One millimolar IPTG was applied to the terrific broth culture and allowed to grow overnight. Samples were collected on a regular basis (4, 8, 12, 16, and 20 H). The expression levels of IL-29 produced by 1 mM IPTG and autoinduction were evaluated using SDS-PAGE. Higher expressing colonies were selected for large scale production.
IL-29 production in E. coli through IPTG induction and autoinduction
To evaluate the expression of IL-29 in autoinduction media and IPTG induced media, the selected colonies were grown in one-liter batches. Autoinduction media was prepared using previously published protocol [23,24]. Selected colony was cultured in 2 different flasks containing 500 mL autoinduction medium (1% tryptone, 0.5% yeast extract, 25 mM potassium dihydrogen phosphate, 50 mM ammonium chloride, 25 mM dipotassium hydrogen phosphate, 0.5 percent glycerol, 0.05 percent glucose, 0.2 percent lactose.) in 2 L baffled flask at 37 °C. Similarly, selected colony was cultured in 2 different flasks containing 500 mL terrific broth in 2 L baffled flask and allowed to grow for 8 H. Growth culture was then induced with 1 mM IPTG and allowed to grow for 16 H. After 24 H of culturing, biomass from both batches was harvested by centrifugation at 8000 g for 15 minutes, and was stored at -80 °C.
Inclusion bodies isolation
The harvested biomass from both the batches were resuspended in autoclaved Milli-Q water at a ratio of 1 g biomass per 10 ml of water. Meanwhile, cell lysis French press cell disrupter (Continuous Flow Cell Disruptor, Constant Systems LTD) was chilled and sterilized with Milli-Q Water and 0.5 M NaOH at a pressure of 0.5 Bar. The dissolved cell pellet was run through the cell disruptor twice at a pressure of 1.5 bars, resulting in the collection of ruptured cells. The collected fraction was centrifuged at 12,000 rpm at 4°C for 30 min. Following washing twice with autoclaved Milli-Q water to eliminate cell debris, host cell’s DNA and endotoxin, the greenish supernatant was discarded, and a hard pellet was retrieved.
Solubilization of IBs and Protein refolding
A solubilization buffer of 6 M Guanidine HCL, 100 mM Tris-Cl (pH 8.0), and 2 mM EDTA (pH 8.0) was produced, and 1g of IBs was resuspended in 33 ml of the buffer at room temperature using simple vortexing. After being stirred on a magnetic stirrer for 30 minutes, the sample was centrifuged at 4°C at 10,000 rpm. The supernatant was retained, and the insoluble material was discarded. While a 200 μL sample was preserved for SDS-Page analysis, the remaining solubilized protein was refolded. To determine protein quantity in solubilized buffer, Bradford assay was performed. To begin the refolding process, the solubilized protein was dropwise mixed with the freshly made refolding buffer (consisting of 0.5M L-Arginine, 100 mM Tris-Cl, Cystine, Cystine, and 0.1% Tween 20). The reaction was then constantly stirred overnight at 20°C.
Diafilteration and purification
To purify the IL29 protein, it underwent diafiltration to eliminate unwanted components such as L-arginine and salts. Initially, the refolded IL-29 was filtered through a 0.45 um membrane, and then diafiltered and concentrated with Millipore tangential flow filtration (TFF) system that featured cut-off size of 10 KDa hollow fibre filter, at a 3 mL/min flow rate. A 20 mM sodium phosphate buffer at pH 7.4 was used as an exchange buffer, with a flow rate of 3 ml/min into the retentate reservoir. A buffer exchange of 90% was carried out using six times the original sample volume. For the 600 ml protein sample, about 3 L of sodium phosphate buffer were employed, which was subsequently concentrated down to 300 ml.
AKTA explorer system from GE Healthcare was utilized to achieve ultimate purification of the protein. The ion exchange column (XK26/10) was employed in the final purification step, loaded with pre-calibrated Source S Resin in sodium phosphate buffer with a pH of 7.4 and a bed volume of 10 mL. IL-29 in sodium phosphate buffer was introduced into the column at a 3 mL/min flow rate and the flow-through was collected. Washing with three to four column volumes of the same buffer was conducted to remove impurities and bind proteins. Ultimately, the sample was eluted using a linear gradient of buffer (sodium phosphate, 0.5M NaCl, pH 7.4), and all fractions were collected separately based on detector's graph. To ensure purity, all fractions were evaluated using SDS-PAGE, and their concentrations were determined by the Bradford assay. The purified protein fractions were stored at 4°C until confirmation, followed by preservation at -80°C.
SDS-PAGE and Western blot
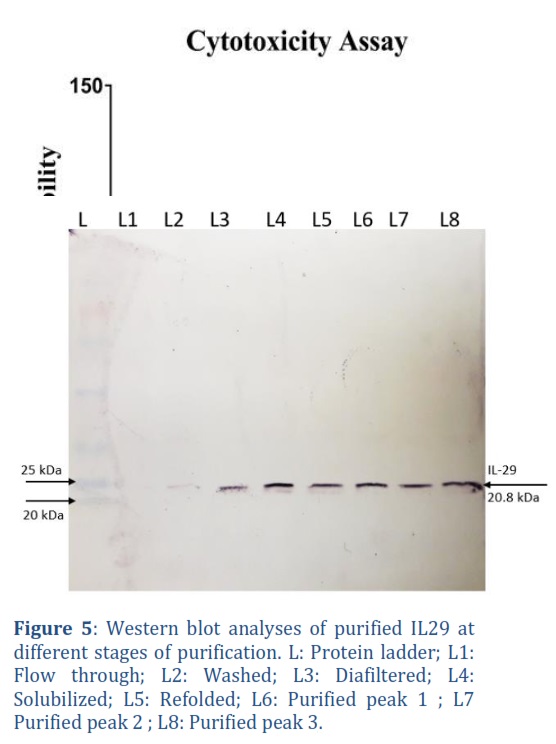
To analyze the quality and purity of protein at different stages of purification process, protein was run on 12% SDS-PAGE using the Hoffergel apparatus. Samples for SDS-PAGE were prepared by mixing equal of protein sample and 2X loading dye, followed by 10 min heat shock in boiling water. After centrifugation, 20 μL of supernatant was loaded onto 12% SDS-PAGE gel with protein ladder and run at 110 volts for 90 minutes. The gel was subsequently stained with Coomassie blue dye to visualize bands.
For western blot analysis, with the semi-dry blot Hoefer’s Semiphor system, protein bands from gel were deposited on to nitrocellulose membrane at 15 V for 30 minutes. Unexposed area of nitrocellulose membrane was blocked with skim milk in PBST. After several washes with PBS, membrane was incubated with anti-IL-29 monoclonal antibody at continues shaking for 1 hour. Then after several washes, membrane was incubated with anti-goat AP-conjugated antibody for 1 hour. Finally, protein bands were developed using the NBT/BCIP substrate.
IEX-HPLC
Purified IL-29 was evaluated using TSK gel columns (7.5 x 50 cm) in Size Exclusion Chromatography-High Performance Liquid Chromatography (SEC-HPLC) (Schemadzu LC-20 systems), alongside a standard. The column was equilibrated with buffer (0.2M sodium chloride, 20mM sodium acetate pH 5.5, and 10% ethanol in water) at 1 mL/min flow rate. Subsequently, 50 µL of Sample and Standard were loaded on to the column, and a linear gradient of 1M sodium chloride (0-100%) was employed to elute the protein. The resulting chromatogram was over a period of 60 minutes at 280 nm.
Biological activity assay
We obtained the HepG2 cell line from the CEMB cell line repository to conduct in vitro testing. The cells were cultured in RPMI-1640 medium with 10% fetal bovine serum and 2 mM l-glutamine at 37°C, 5% CO2, 95% air, and 100% relative humidity in a T-75 flask for 24 hours. Following growth, 100 µL media with cells were seeded onto 96-well plates at a concentration of 5 X 103 cells per well. IL-29 was diluted to a concentration of 0.5 mg/mL using water. On the following day, each well was filled with100 mL medium and maintained under standard conditions with drug (IL-29) concentrations of 0.078, 0.156, 0.31, 0.625, 1.25, 2.5, and 5 µg/mL for 48 hours. Following incubation, media was discarded, and 100 µL of MTT solution (5 mg/mL in 1X PBS) was introduced to every well. The plate was incubated for 3 hours in aluminum foil in humid atmosphere at 37 °C. The formazan crystals in each well were dissolved using 100 µL of DMSO, and after 24 hours metabolic activity was measured at 570 nm using a microplate reader.
Results![]()
IL29 cloning and expression cassette Construction
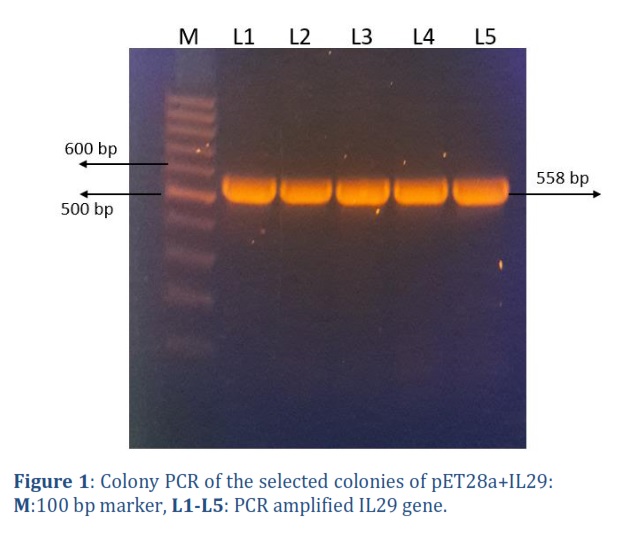
The IL-29 gene was synthesized from IDT USA, cloned in pET28a, and transformed into E. coli TOP10F strain. After transforming the plasmid construct carrying gene of interest into E. coli, colony PCR was performed on 5 selected colonies using IL-29 specific primers, and on agarose gel a 558 bp band was observed against reference 100 bp ladder Figure 1. The presence of 558 bp band confirmed the successful cloning and its transformation into E. coli TOP10F.
Expression of IL-29
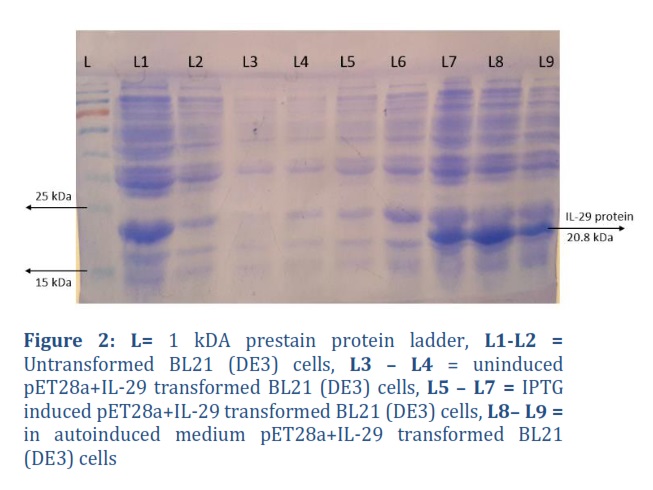
The expression host BL21 (DE3) was transformed with a positive construct (pET28a+IL-29). IL-29 was expressed in autoinduced media and IPTG induced media. Total Protein expressed in both the media was run on a 12% SDS-PAGE, a protein band of approximately 21 kDa in size was observed, which matched the actual size of the IL-29 protein. The crude protein from the autoinduced culture and the culture induced with 1 mM IPTG were analyzed. Figure 2, L7, L8 and L9 revealed that these colonies are expressing IL-29 protein. Later, in large scale fermentation, the harvested biomass from IPTG induced media was around 11.8 g, while biomass from autoinduced media was 13.4 g, while their respective IBs were 3.8 g and 4.8 g.
IBs isolation and Solubilization
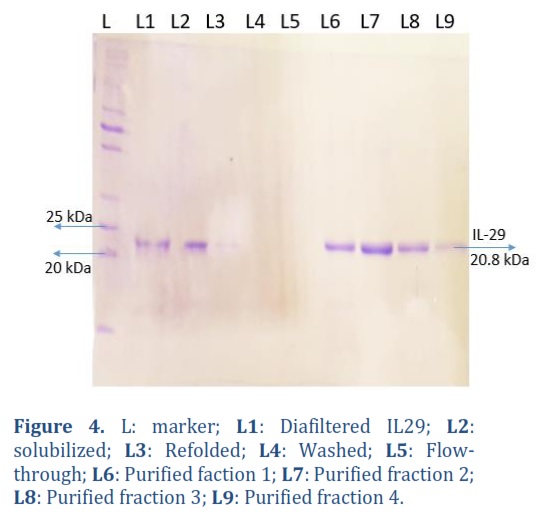
After passing biomass through cell disruptor, inclusion bodies were isolated. Around 4.1 Ibs were isolated from 14.3 g of biomass. To solubilize IBs obtained, 33 mL buffer per 1 g of IBs. Protein concentration was determined, which was around 5.2 mg/mL, and diluted further in refolding buffer to achieve optimal refolding results at a different final concentration. The solubilized protein was analyzed using SDS-Page and can be observed in Figure 4.
Refolding and purification of IL-29
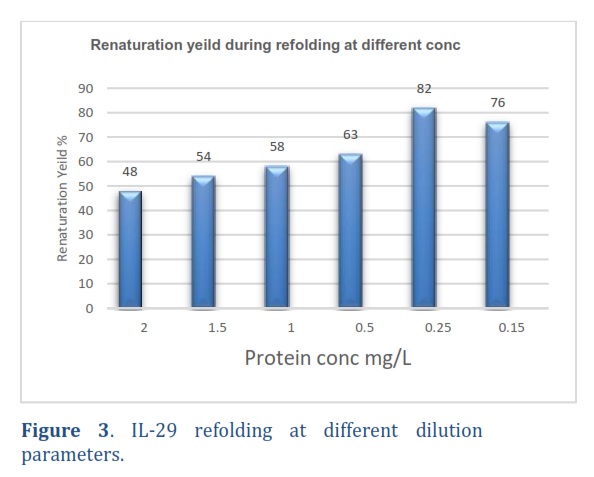
The protein solubilized was diluted to different final concentrations in the refolding buffer. High-Performance Liquid Chromatography (HPLC) was used to measure the proportions of properly folded, misfolded, and precipitated proteins. Some percentage of protein was observed to be precipitated and misfolded, while the remaining was recovered after being diluted to 2 mg/mL final concentration. At 0.25 mg/mL concentration, the protein recovery rate was 82%, and further dilution did not impact the final recovery as can be observed in the graph in Figure 3. Therefore, a concentration of 0.25 mg/mL was used for all subsequent refolding processes. The above figure shows that when recombinant IL-29 was diluted to a concentration of 0.25 mg/mL in the refolding buffer, it exhibited high levels of renaturation across all final concentrations.
The refolded IL-29 proteins underwent concentration and diafiltration using a 10 kDa hollow fibre in Tangential Flow Filtration (TFF) through the Millipore-TFF system with an 800 mL refolding buffer from Merck. The refolding buffer was then replaced with a PBST buffer six times its volume. Finally, total volume was concentrated to 300 ml in PBST buffer, resulting in a loss of approximately 20% of the protein during the concentration and diafiltration processes. In Figure 4, Lane 2 depicts the protein sample that underwent diafiltration. Following concentration and diafiltration, IL-29 protein was purified using the AKTA explorer system (GE Healthcare) through cation exchange chromatography on a Source S Resin, XK16/20 column with a bed volume of 10 ml. Protein was eluted from the column with a linear gradient of 0.5M NaCl in 20mM phosphate buffer with a pH of 7.0. Multiple protein fractions were obtained, analyzed on SDS gels, and quantified using a UV spectrophotometer. Highly pure IL-29 at a concentration of 132 mg/L (13 mg/mL) was obtained. Figure 6 shows purified IL-29 protein against IL-29 standard.
Characterization of IL-29
The purity and concentration of IL-29 were examined using SDS-PAGE throughout different stages, including solubilization, refolding, diafiltration, and purification, as illustrated in Figure 4. To confirm the presence of IL-29 protein, Western blot analysis was performed on various purification fractions using mouse-raised human-IL29 specific monoclonal antibodies (anti-IL29, Cat #CF812899 Thermo scientific). The NBT/BCIP system (Sigma) was employed for staining, which produces a black colour showing the presence of IL-29 as can be seen in Figure 5.
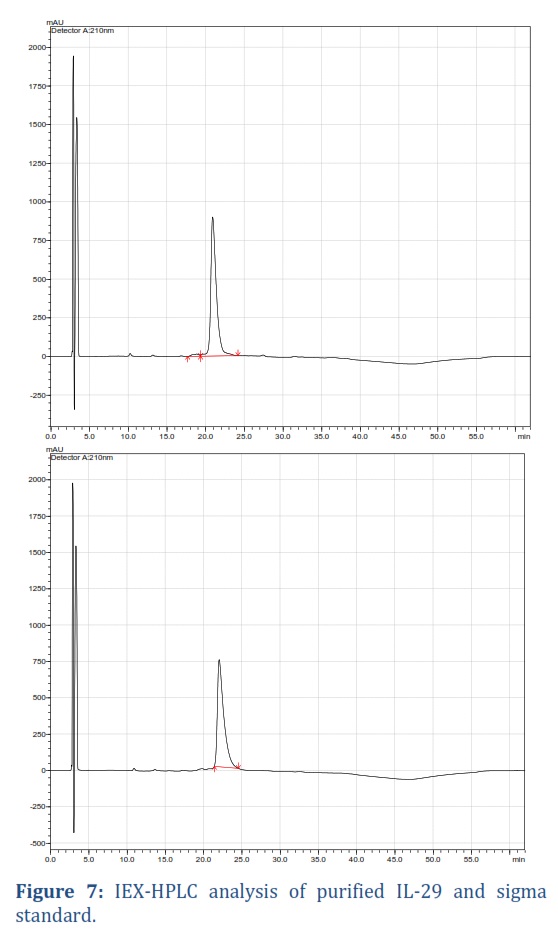
IEX-HPLC analysis of purified protein
To validate the purity of the product, HPLC was used to confirm the presence of a single peak. The results showed that up to 97% of rIL-29 was pure, with a retention time of 22.02 minutes, matching the reference standard, as demonstrated in Figure 7.
Cytotoxicity assay
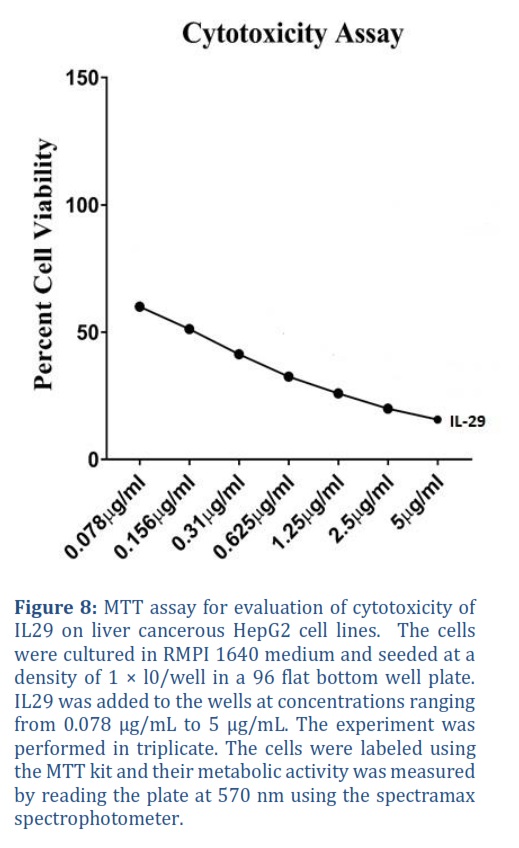
To find cytotoxicity of recombinant IL-29, drug was applied on cancerous cell line HEPG2 and performed MMT assay to find percent cell viability. The drug was applied in dose dependent manner at various concentrations of IL-29 ranging from 0.078 μg/mL – 5 μg/mL. Treatment of cells with IL-29 resulted in a significant reduction in their metabolic activity. Around, 50 % cells were killed at 0.156 μg/mL, while 80 % of cells died at a concentration of 5 μg/mL as can be observed in Figure 8. This shows that purified recombinant IL-29 is inactive form.
Figures & Tables

Constructed plasmid containing IL-29 gene were transformed into the BL21 (DE3) E. coli expression host. Due to its ease of culturing, low costs, high protein production, and FDA approval for human use, the E. coli expression system is the favored approach for generating therapeutic proteins, both in laboratory settings and on an industrial level [25]. Biopharmaceuticals of significant clinical importance, including interleukins, growth hormones, and proinsulin are manufactured in bacterial expression system [19]. Foreign proteins synthesized in E. coli can account for anywhere between 5% to 50% of the total protein produced within the cell [26].
BL21 (DE3) has been identified as an ideal bacterial expression strain for toxic protein synthesis, primarily due to the presence of the lambda DE3 lysogen which produces Interleukins (IL) in high amount compared to other bacterial expression strains [13]. In this study, Expression of IL-29 in autoinduction media and Terrific broth media induced with 1mM IPTG were compared. The biomass obtained from IPTG induced media was 11.8 g and autoinduced media was 13.4 g/L of batch, while their Ibs were 3.8 and 4.8 respectively. In order to achieve large-scale production of economically valuable therapeutic proteins, purification is an essential step. Significant progress has been made in the field of interleukin purification, with various techniques available such as antibody affinity, GST tag-fusion, FPLC chromatography, and ion exchange. For instance, in the case of IL-29 protein, cation exchange chromatography was utilized to accomplish the purification process. The yield of protein obtained from a one-litre batch at a concentration of 13 mg/mL was 132 mg, and HPLC and SDS-PAGE analyses indicated a high purity of 97%. The protein remained stable when stored at 4°C for several months, although some precipitation occurred after four months of storage in PBS buffer. Active purified recombinant IL-29 was applied to malignant HepG2 cell lines in a dose-dependent manner, showing its efficacy. Cells treated with IL-29 exhibited significantly lower metabolic activity, with 50% of cells dying at a concentration of 0.156 μg/mL, and 80% of cells dying at a concentration of 5 μg/mL [26].
In this study, the expression of recombinant IL-29 protein in E. coli was achieved through the utilization of an autoinduction method, which, when combined with a one-step purification process, resulted in the production of 132 mg/L of physiologically active IL-29 with a purity level of over 97%. The approach outlined in this study could be applied to the scaling up of IL-29 and other recombinant protein syntheses in E. coli.
Author Contributions
Conceived and designed the experiments: Zia ur Rahman, Numan Fazal, Muhammad Islam Khan, Mohsin Ahmed khan, Saad Tahir, Muhammad Akram
Performed the experiments: Zia ur Rahman, Numan Fazal, Muhammad Islam Khan
Analyzed the data: Zia ur Rahman
Wrote the paper: Zia ur Rahman
Critical Review: Nadeem Ahmed, Sajjad Ullah, Ahmad Usman Zafar
![]()
The authors declare no conflict of interest regarding the publication of this article.![]()
References
- Berraondo P, Sanmamed MF, Ochoa MC, Etxeberria I, Aznar MA, et al. Cytokines in clinical cancer immunotherapy. British journal of cancer, (2019); 120(1): 6-15.
- Sleijfer S, Bannink M, Van Gool AR, Kruit WH, Stoter G. Side effects of interferon-α therapy. Pharmacy world and science, (2005); 27423-431.
- Andreakos E, Zanoni I, Galani IE. Lambda interferons come to light: dual function cytokines mediating antiviral immunity and damage control. Current opinion in immunology, (2019); 5667-75.
- Meager A, Heath A, Dilger P, Zoon K, Wadhwa M, et al. Standardization of human IL-29 (IFN-lambda1): establishment of a World Health Organization international reference reagent for IL-29 (IFN-lambda1). Journal of Interferon & Cytokine Research, (2014); 34(11): 876-884.
- Chrysanthopoulou A, Kambas K, Stakos D, Mitroulis I, Mitsios A, et al. Interferon lambda1/IL-29 and inorganic polyphosphate are novel regulators of neutrophil-driven thromboinflammation. The Journal of Pathology, (2017); 243(1): 111-122.
- Steen HC, Gamero AM. Interferon-lambda as a potential therapeutic agent in cancer treatment. Journal of Interferon & Cytokine Research, (2010); 30(8): 597-602.
- Kelm NE, Zhu Z, Ding VA, Xiao H, Wakefield MR, et al. The role of IL-29 in immunity and cancer. Critical Reviews in Oncology/Hematology, (2016); 10691-98.
- Koch S, Finotto S. Role of interferon-λ in allergic asthma. Journal of innate immunity, (2015); 7(3): 224-230.
- Wills‐Karp M. Interleukin‐13 in asthma pathogenesis. Immunological reviews, (2004); 202(1): 175-190.
- O’Brien TR, Thomas DL, Jackson SS, Prokunina-Olsson L, Donnelly RP, et al. Weak induction of interferon expression by SARS-CoV-2 supports clinical trials of interferon lambda to treat early COVID-19. Clinical Infectious Diseases, (2020); 71(6): 1410-1412.
- Marcello T, Grakoui A, Barba–Spaeth G, Machlin ES, Kotenko SV, et al. Interferons α and λ inhibit hepatitis C virus replication with distinct signal transduction and gene regulation kinetics. Gastroenterology, (2006); 131(6): 1887-1898.
- Kanda T, Wu S, Kiyohara T, Nakamoto S, Jiang X, et al. Interleukin-29 suppresses hepatitis A and C viral internal ribosomal entry site-mediated translation. Viral Immunology, (2012); 25(5): 379-386.
- Wang J, Oberley-Deegan R, Wang S, Nikrad M, Funk CJ, et al. Differentiated human alveolar type II cells secrete antiviral IL-29 (IFN-λ1) in response to influenza A infection. The Journal of Immunology, (2009); 182(3): 1296-1304.
- Qian Z, Travanty EA, Oko L, Edeen K, Berglund A, et al. Innate immune response of human alveolar type ii cells infected with severe acute respiratory syndrome–coronavirus. American journal of respiratory cell and molecular biology, (2013); 48(6): 742-748.
- Egli A, Santer DM, O’Shea D, Tyrrell DL, Houghton M. The impact of the interferon-lambda family on the innate and adaptive immune response to viral infections. Emerging microbes & infections, (2014); 3(1): 1-12.
- Sommereyns C, Paul S, Staeheli P, Michiels T. IFN-lambda (IFN-λ) is expressed in a tissue-dependent fashion and primarily acts on epithelial cells in vivo. PLoS pathogens, (2008); 4(3): e1000017.
- Novotny LA, Evans JG, Su L, Guo H, Meissner EG. Review of lambda interferons in hepatitis B virus infection: outcomes and therapeutic strategies. Viruses, (2021); 13(6): 1090.
- Rosano GL, Ceccarelli EA. Recombinant protein expression in Escherichia coli: advances and challenges. Frontiers in microbiology, (2014); 5172.
- Kamionka M. Engineering of therapeutic proteins production in Escherichia coli. Current pharmaceutical biotechnology, (2011); 12(2): 268-274.
- Studier FW, Moffatt BA. Use of bacteriophage T7 RNA polymerase to direct selective high-level expression of cloned genes. Journal of molecular biology, (1986); 189(1): 113-130.
- Selas Castiñeiras T, Williams SG, Hitchcock AG, Smith DC. E. coli strain engineering for the production of advanced biopharmaceutical products. FEMS microbiology letters, (2018); 365(15): fny162.
- Jia B, Jeon CO. High-throughput recombinant protein expression in Escherichia coli: current status and future perspectives. Open biology, (2016); 6(8): 160196.
- Bashir H, Ahmed N, Khan MA, Zafar AU, Tahir S, et al. Evaluating the autoinduction expression system and one‐step purification for high‐level expression and purification of gallbladder‐derived rhIL‐1Ra. Biotechnology and Applied Biochemistry, (2017); 64(1): 20-26.
- Studier FW. Protein production by auto-induction in high-density shaking cultures. Protein expression and purification, (2005); 41(1): 207-234.
- Gupta V, Sengupta M, Prakash J, Tripathy BC, Gupta V, et al. Production of recombinant pharmaceutical proteins. Basic and applied aspects of biotechnology, (2017); 77-101.
- Schumann W, Ferreira LCS. Production of recombinant proteins in Escherichia coli. Genetics and Molecular Biology, (2004); 27442-453.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0