

Full Length Research Article
Estimate the relationship between SNP of the IL-17 gene (rs2275913) and cardiovascular diseases (CVDs)
Anees Abdul Hassan Thamer Al-Himami*, Ali Naeem Salman Sara Hameed Rajab1*, Mayada Abdullah Shehan2
Adv. life sci., vol. 12, no. 3, pp. 547-552, August 2025
*- Corresponding Author: Anees Abdul Hassan Thamer Al-Himami (Email: anees.bio2022@utq.edu.iq)
Authors' Affiliations
[Date Received: 03/01/2025; Date Revised: 26/01/2025; Available Online: 31/10/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: One cytokine that may be important in cardiovascular diseases (CVD) is interleukin-17 (IL-17). The functions of IL-17 and its receptor genes can be affected by functional genetic variations.
Methods: The study examined the relationship between the IL-17A gene's rs2275913 single-nucleotide polymorphism (SNPs) and cardiovascular diseases (CVD) in 40 samples (30 patients and 10 controls).
Results: The rs2275913 variant had a homozygous GG, heterozygous GA, and homozygous AA pattern in the population under investigation. Most patients were in the homozygous GG state, and the homozygous variant AA was present among patients and controls. The IL-17A gene had patterns of polymorphism within the tested Thi-Qar community members.
Conclusion: This study found a genotype-specific association between IL-17A gene rs2275913 variants and cardiovascular disease. The GA heterozygous genotype was significantly higher in healthy volunteers than in patients (p = 0.02), indicating a possible protective effect. While no significant correlation was observed for overall allele frequency, these findings suggest that certain genotypes of this polymorphism may influence CVD prevalence among Thi-Qar residents.
Keywords: IL-17 gene, SNP, Cardiovascular Diseases, rs2275913
Introduction![]()
Across the world, CVDs are among the most frequent agents accountable for premature death. CVDs contributed to the fatality in more than 17.3 million people annually, approximately 17.5 million in 2015, and the number was estimated to reach nearly 23.6 million in 2030 [1].
World mortality rate has increased by 12.5% in 2016 owing to various factors, like progressive acceleration in vascular diseases due to economic pressures on poor and emerging nations. This economic pressure resulted in a higher cost for health care and manpower loss [2].
CVDs are a category of disorders and abnormalities that occur in blood vessels and the heart. They include a vast array of diseases like atherosclerosis and coronary heart disease (CHD), and this second category is a group of disorders due to muscle-supplying coronary artery malfunction or injury with mortal complications to the muscle [3]. Coronary heart diseases are depicted by angina pectoris (AP), heart attack (HA), myocardial infarction (MI), and heart failure (HF). They are outcomes of ischemic complications to the myocardium because of obstruction or stenosis in the coronary arteries that nourish the heart [4].
IL-17 is a potent cellular activator of strong inflammation that is primarily produced by activated T cells (CD4 + T), but this does not mean that they are the only ones that produce it [5].
Since IL-17 signaling promotes a few inflammatory signaling pathways that activate genes that create pro-inflammatory cytokines, it increases the production of growth mediators, tissue remodeling enzymes, and other secondary factors. In most cases, DNA is copied in new cells without any change in it, i.e., it is an identical copy of the original DNA, but sometimes a substitution of a single nucleotide occurs in the new sequence, and this results in a genetic variation known as a Single Nucleotide Polymorphism (SNP) [6].
Inflammatory cytokines, the most active of which is IL-17A, cause vascular endothelial damage, which is associated with increased expression of adhesion molecules and stimulation of chemoattractants, which collectively lead to the development of inflammation. The most significant IL-17 member, IL-17A, is found at 6q12 and is made up of three exons and two introns. Endothelial damage caused by inflammatory cytokines, together with the upregulation of adhesion molecules and the release of chemoattractants, collectively contribute to the propagation of inflammation [7].
CVD development may be influenced by variations in genes connected to signaling pathways for inflammatory cytokines [8].
Methods![]()
Design of study
In Thi-Qar Governorate, 30 samples were taken from patients who visited the heart center and were admitted to the hospital, as well as from patients who were admitted to the Coronary Care Unit due to cardiovascular diseases. As a control group, 10 samples were taken from healthy individuals. To gather data on patients and healthy individuals, a questionnaire was completed by both males and females between 1/12/2022 and 1/4/2023.
Collection of Blood Samples
2 mL of venous blood was taken from 30 patients with cardiovascular diseases and 10 healthy people without cardiovascular diseases. All blood samples were placed in tubes containing an anticoagulant-EDTA and kept at a temperature of -20 °C for the purpose of being used in the DNA extraction process.
DNA Extraction
The DNA extraction process was carried out on 30 blood samples from patients suffering from cardiovascular diseases and 10 healthy control blood samples preserved in EDTA tubes, with several steps mentioned on the leaflet attached to the DNA Extraction Kit manufactured by Geneaid.
DNA sequencing
To identify the genetic patterns of the rs2275913 gene polymorphism in the IL-17A gene, the PCR outputs were sent to Macrogen, South Korea, for DNA sequencing.
Gene mutations occurring in the target gene sequence were identified, and the obtained sequence was compared with the reference gene sequence in NCBI BLAST. Then the results were analyzed by the Mutation Surveyor to check for mutations in the DNA.
Interpretation of the data sequence
The BioEdit Sequence Alignment Editor version 7.1 (DNASTAR, Madison, WI, USA) was used to edit the sequencing findings of the PCR products from various samples. These edited sequences were matched and analyzed with the pertinent sequences in the reference database, and then the results were interpreted.
In the resulting PCR-amplified section, as well as at its corresponding locus within the reference genome, the changes seen in each sample were numbered. SnapGene Viewer version 4.0.4 (http://www.snapgene.com) was used to explain each variant found inside the gene under investigation.
Single Nucleotide Polymorphism (SNP) rs2275913
The dbSNP database received the discovered SNP for verification purposes. The location of each unique SNP within the reference genome was used to define it. Its prior SNP's presence was then confirmed by looking at its corresponding locus in the dbSNP data, and the dbSNP locus was then recorded by the Explorer for Genetic Multiplexing (SNP).
Statistical analysis
Statistical analysis was done for all studied samples using the chi-square test, OR test, and Hardy-Weinberg equilibrium (HWE) to study the frequency of genotypes of a gene under a significance level of p < 0.05, using the SPSS software package.
Results![]()
A total of 40 samples, including 30 patient samples (designated S1-S30) and 10 control samples (designated C1-C10), had their IL-17A genetic sequences examined. Following the NCBI blast (https://blast.ncbi.nlm.nih.gov/Blast.cgi), the exact identity of the genomic segment that is now amplified was determined by the sequencing reactions. This engine demonstrated over 99% sequence identity between amplified specimens and target sequence references, which cover the IL-17A gene extensively. Using a comparison of observed DNA variants of the samples currently under investigation to the returning DNA sequences (GenBank accession number NG_033021), the determination of the specific locations and other particular information of the PCR amplicons was undertaken.
After identifying and annotating the 526 bp PCR fragments, which were discovered on chromosome 6, the forward and reverse primer sites inside the amplified 526 bp segments carrying the rs2275913 SNP were determined.
The results of the sequence alignment of the 526 bp samples found that some of the samples studied from both cases and healthy individuals contained just one nucleic acid change when compared to the pertinent reference sequences.
The prevalence of the discovered polymorphic locus in the examined samples was demonstrated. Therefore, it was essential to look more closely at the places where this variant was found in the IL-17 gene's genomic DNA sequences that were part of the dbSNP database. The matching location of the IL-17 gene was retrieved from the dbSNP server (https://www.ncbi.nlm.nih.gov/projects/SNP/) to determine the targeted variant's genomic positions using the sequenced 526 bp fragments' deposited SNP database. Additionally, using the GenBank Acc. no. NG_033021.1, graphical representations of the IL-17 dbSNP database on chromosome no. 6 were shown to recognize the additional characteristics of the discovered SNP. The discovered G271A SNP was known in the genome, according to a review by the dbSNP engine.
The identified variations sequencing chromatogram files and comprehensive annotations were recorded and shown in accordance with the appropriate location in the generated 526 bp PCR amplicons.
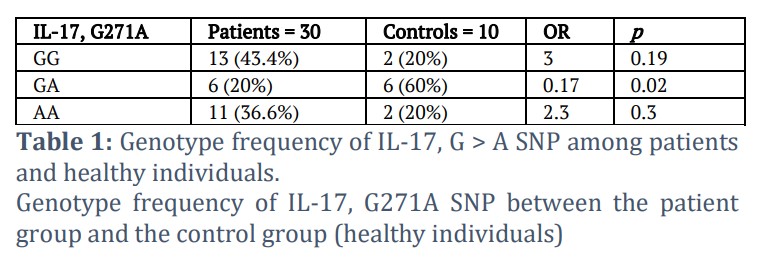
The intended high-frequency SNP, 271G > A, or G271A, was found in the examined samples, where the G nucleotide was changed to an adenine (A) nucleotide at position 271 of the produced PCR segments. The three polymorphic patterns GG, AA, and GA were the most frequent when this frequency variant was found. Homozygous GG status was found in several patients (13/30) and controls (2/10) samples, but the heterozygote pattern GA was found in a low-frequency ratio in patients (6/30) and high frequency in controls (6/10). It's interesting to note that both the patients (11/30) and the healthy group (2/10) had the homozygous AA pattern.
Hardy-Weinberg equilibrium
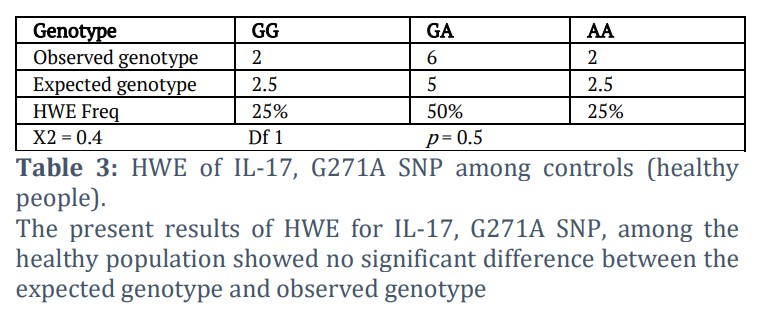
A basic theory known as the Hardy-Weinberg equilibrium (HWE) holds that, without unfavorable circumstances, a population's genetic diversity would remain constant over the course of many generations. To compute HWE, the genotype and allele frequency of the IL-17, G271A SNP are provided in Tables 1, 2, and 3. The genotype distribution in the control group was in Hardy-Weinberg Equilibrium (p > 0.05), as shown in Table 3.
The present results of the study revealed the frequency of the genotype (Genotype frequency of IL-17, G > A SNP) between the patient group and the healthy group. The frequency of the genotype GG was 13 in the group of patients, with a percentage of 43.4%, while in the control group, the number was 2, with a rate of 20%, as there was no statistically significant difference (p = 0.19). As for the pattern GA, the number of cases in the patient group was 6 cases, with a rate of 20%, and the control group had 6 cases, with a rate of 60%, where the difference was statistically significant (p = 0.02). As for the genotypic pattern AA, its frequency was 11 in the sick individuals’ group with a percentage of 36.6%, while in the healthy group it was 2, with a percentage of 20%, as there was no change that is statistically significant (p = 0.3) according to Table 1.
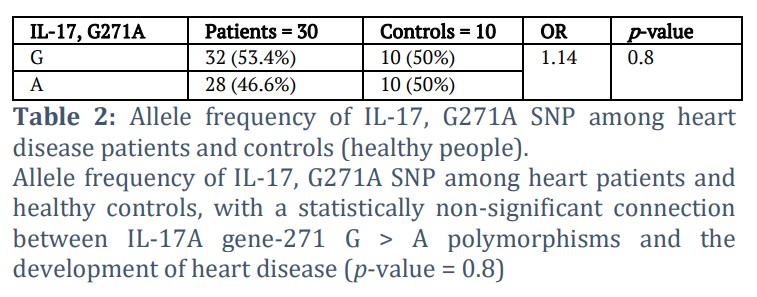
Allele frequency of IL-17, G271A SNP among heart patients and healthy people has a statistically non-significant connection between IL-17A gene, G271A polymorphisms and the development of heart disease (p-value = 0.8), as shown in Table 2.
The genotype distribution in the control group was in Hardy–Weinberg Equilibrium (p > 0.05), indicating no significant difference between the observed and expected genotype frequencies, as shown in Table 3.
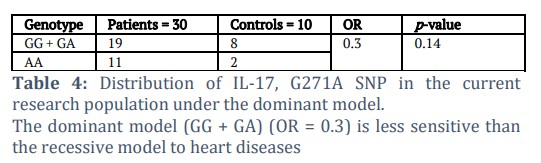
According to the distribution of IL-17, the G271A SNP in this research population under recessive and dominant models (Tables 4 and 5) showed non-significant results, so the recessive model (GA + AA) (OR = 2.7) was more sensitive to heart diseases than the dominant model.
Figures & Tables

The dbSNP service (https://www.ncbi.nlm.nih.gov/projects/SNP/) was used to obtain the gene. A graphical representation of the IL-17 dbSNP database on chromosomal number 6 (GenBank Acc. NG_033021.1) was done to determine the kind of these SNPs by going over each of the fourteen SNPs that the dbSNP engine has discovered.
An essential inflammatory cytokine for bridging innate and adaptive immunity is IL-17A. According to molecular research, IL-17 is an essential cytokine that promotes inflammation, stimulates the release of several chemokines and cytokines from several cell types, including myeloid cells and mesenchymal cells, which help to recruit neutrophils and monocytes into the inflammatory milieu [9].
Interleukin-17, a brand-new cytokine family with 6 protein members (IL-17A–IL-17F), has a major impact on several chronic inflammatory diseases [10]. Activated CD4 + T cells (Th17 cells), lymphoid tissue inducers (LTi-like cells), neutrophils, and NK cells have the potential to produce IL-17 [11]. Along with this, changes in the genes that direct the pathways to inflammatory cytokines can result in the development of CVD [12].
No correlation was observed in this research in the context of cardiac disorders and IL-17 271 G > A SNP polymorphism (p = 0.8). While there wasn't a statistically significant correlation of overall allele frequency to risk of CVD (p = 0.8), In this study, there was, a significant correlation in specific genotypes studied. The GA heterozygous genotype was significantly increased in healthy controls compared to patients (p = 0.02) and would likely have a protective role against CVD. The homozygous genotypes (AA and GG) did not have a statistically significant correlation. Therefore, while this polymorphism would not be contributing significantly to risk at an allele level, there appears to be an association of this particular GA genotype to decreased occurrence of CVD in this patient population. Zhang et al., [13] and Pei et al., [14], in two reports, have provided associations among IL-17 gene polymorphism and CVD risk.
The key transcription factor binding motif responsible for NFAT binding activity with the A allele was significantly strengthened by the variation at position rs2275913 in the promoter region within the IL-17A gene [15].
Results from recent studies showed that the G > A (rs2275913) polymorphism was not significantly associated with heart diseases; these results were combined with the research undertaken by Espinoza et al., [16], which revealed no significant correlation with the number of stenotic vessels and the polymorphism at the level of the rs2275913. This would suggest that this genetic polymorphism is not effective in providing an estimation of CVD severity. According to the statistics from the allele frequencies, the G allele was more frequently represented and the A allele was less frequently represented in the patient group (n = 32), while both G and A alleles were equally frequent in the control group (10 G alleles, 10 A alleles). IL-17A's G (rs2275913) promoter polymorphism is thought to play a role in the pathophysiology of a number of inflammatory disorders. Regarding the significance of the rs2275913 polymorphism in the CVD etiology, contradictory findings have been presented.
The current findings revealed that the frequency of the minor A allele was lower in patients (28/60 = 46.6%) than in controls (10/20 = 50%). The prevalence of the A allele was recorded in populations of Brazil (28.30%) and China (29.64%) [17]. However, its higher prevalence approximated the stated frequency for the Middle Eastern Chinese population (46.94%) [18] and was higher than the A allele’s documented frequency in the Mexican population (19.04%) [19].
Additionally, the genotype distribution of the IL-17 gene showed that the most frequent genotype among patient samples (13; 43.4%) and control samples (2; 20%) was GG, whereas 11 patient samples (36.6%) and 2 control samples (20%) had the AA genotype. According to a functional investigation, the rs2275913 A allele is linked to increased IL-17A promoter activity, indicating that the rs2275913 AA genotype may be responsible for increased inflammation levels [20]. The chance of developing coronary heart disease was reported to be higher in those with the AA genotype and the GA + AA genotype of rs2275913 than in people with the GG genotype [21].
IL-17 rs2275913 polymorphisms were linked to an increased risk of giant cell arteritis, according to Márquez et al., [22]. Geng et al., [23] found that the AA genotype and A allele of the rs2275913 genetic variant greatly enhanced the chance of developing CVD. Other research, however, did not support this genetic polymorphism's contribution to the development of CVD. A major limitation of this study is the very small control group (n = 10). This reduces statistical power and limits the generalizability of the results. Future studies should employ larger and more balanced case–control samples to draw more robust conclusions.
The present results support the findings of Zhao et al. [18] but contradict those of Geng et al. [23], who reported that the A allele and AA genotype were associated with increased risk of CVD, whereas in this study the GA genotype appeared protective, and the A allele frequency was lower in patients than in controls. The relatively lower frequency of the A allele in CVD cases (46.6%) compared to healthy controls (50%), and correspondingly significantly increased frequency of GA genotype in controls, suggests specific inheritance modes in this region possess protective effects against CVD. Consistent with the result in other research, Zhao et al., [18] demonstrated that the risk of CVD was linked to genotypes AA and AG/AA of the rs2275913. But when the study was adjusted for the demographics of the participants, no significant correlation was left to suggest that patient factors can influence the correlation between rs2275913 and risk of CVD [18].
Pei et al., [14] investigated the associations of IL-17 gene polymorphisms with cardiovascular diseases. IL-17F rs763780 polymorphism is not predicted to play a part in myocardial infarction development, according to the results of one case-control study in Chinese individuals. According to Zhang et al., [13], IL-17A rs8193037 is associated with greater CVD incidence and increased expression of IL-17A in acute myocardial infarction (AMI). In line with Vargas-Alarcón et al., [19] research, IL-17A haplotypes played a part in CVD development. Interestingly, IL-17A plays a positive part in various infections, including influenza A virus (H5N1), through the increased response to infections and recruitment of B cells to the lung [24]. Because of this reason, their polymorphism is possibly linked with the development and progression of several lung disorders, including infection in the lung [25], lung fibrosis [26], non-small cell lung carcinoma [26], and lung neutrophilia [27].
The IL-17 gene polymorphism has also played various roles, and the G > A variant in the position of rs2275913 has also contributed to a greater risk for diseases such as pancreatic cancer [28], carcinogenesis related to chronic inflammation [29], breast cancer [30], and diet-induced obesity [31]. Genetic polymorphism in the IL-17A gene was linked with various severe cellular dysfunctions [32-36], for instance, Chagas disease, lupus nephritis, Ankylosing Spondylitis, inflammatory arthritis, and several others [37-39].
Acknowledgement
The authors acknowledge the College of Science, University of Anbar, Iraq, for providing research facilities.
Author Contributions
Anees Abdul-Hassan Thamer and Ali Naeem Salman participated equally in experimentation, data analysis, writing, and finalization of the research paper.
The authors declare no conflict of interest in the publication of this manuscript.![]()
References
- Allam G, Abdel-Moneim A, Gaber AM. The pleiotropic role of interleukin-17 in atherosclerosis. Biomedicine & Pharmacotherapy, (2018); 106(2018): 1412–1418.
- Rehman S, Rehman E, Ikram M, Jianglin Z. Cardiovascular disease (CVD): assessment, prediction and policy implications. BMC public health, (2021); 21(2021): 1-14.
- Reamy BV, Williams PM, Kuckel DP. Prevention of cardiovascular disease. Primary Care: Clinics in Office Practice, (2018); 45(1): 25-44.
- Sanchis-Gomar F, Perez-Quilis C, Leischik R, Lucia A. Epidemiology of coronary heart disease and acute coronary syndrome. Annals of translational medicine, (2016); 4(13): 256.
- Veldhoen M. Interleukin 17 is a chief orchestrator of immunity. Nature immunology, (2017); 18(6): 612-621.
- O'Neill JP. DNA damage, DNA repair, cell proliferation, and DNA replication: how do gene mutations result?. Proceedings of the National Academy of Sciences, (2000); 97(21): 11137-11139.
- Ramji DP, Davies TS. Cytokines in atherosclerosis: Key players in all stages of disease and promising therapeutic targets. Cytokine & growth factor reviews, (2015); 26(6): 673-685.
- Seidi A, Mirzaahmadi S, Mahmoodi K, Soleiman-Soltanpour M. The association between NFKB1-94ATTG ins/del and NFKB1A 826C/T genetic variations and coronary artery disease risk. Molecular Biology Research Communications, (2018); 7(1): 17-24.
- Iwakura Y, Ishigame H, Saijo S, Nakae S. Functional specialization of interleukin-17 family members. Immunity, (2011); 34(2): 149-162.
- Cătană CS, Neagoe IB, Cozma V, Magdaş C, Tăbăran F, et al. Contribution of the IL-17/IL-23 axis to the pathogenesis of inflammatory bowel disease. World journal of gastroenterology: WJG, (2015); 21(19): 5823-5830.
- Cua DJ, Tato CM. Innate IL-17-producing cells: The sentinels of the immune system. Nature Reviews Immunology, (2010); 10(7): 479-489.
- Shuang L, Li Z, Chen F, Cui X, Ning Y, et al. Association between interleukin-17 gene polymorphisms and risk of coronary artery disease. International journal of clinical and experimental pathology, (2015); 8(9): 11653-11658.
- Zhang X, Pei F, Zhang M, Yan C, Huang M, et al. Interleukin-17A gene variants and risk of coronary artery disease: a large angiography-based study. Clinica chimica acta, (2011); 412(3-4): 327-331.
- Pei F, Han Y, Zhang X, Yan C, Huang M, et al. Association analysis of the IL-17F His161Arg polymorphism in myocardial infarction. Coronary artery disease, (2009); 20(8): 513-517.
- Sahu U, Biswas D, Prajapati VK, Singh AK, Samant M, et al. Interleukin‐ 17—A multifaceted cytokine in viral infections. Journal of Cellular Physiology, (2021); 236(12): 8000-8019.
- Espinoza JL, Takami A, Nakata K, Onizuka M, Kawase T, et al. A genetic variant in the IL-17 promoter is functionally associated with acute graft-versus-host disease after unrelated bone marrow transplantation. PLoS one, (2011); 6(10): e26229.
- Zacarias JM, Sippert EÂ, Tsuneto PY, Visentainer JE, Silva CD, et al. The influence of interleukin 17A and IL-17F polymorphisms on chronic periodontitis disease in Brazilian patients. Mediators of inflammation, (2015); 2015(1): 147056.
- Zhao Q, Jiang H, Ma T, Qiu H, Guo M, et al. The association between IL-17A and IL-23R polymorphisms and coronary artery disease risk in a Middle Eastern Chinese population. Journal of Clinical Laboratory Analysis, (2019); 33(6): e22893.
- Vargas-Alarcon G, Angeles-Martinez J, Villarreal-Molina T, Alvarez-Leon E, Posadas-Sanchez R, et al. Interleukin-17A gene haplotypes are associated with the risk of premature coronary artery disease in Mexican patients from the genetics of atherosclerotic disease (GEA) study. PLoS one, (2015); 10(1): e0114943.
- Min X, Lu M, Tu S, Wang X, Zhou C, et al. Serum cytokine profile in relation to the severity of coronary artery disease. BioMed research international, (2017); 2017(1): 4013685.
- Bao MH, Luo HQ, Xiang J, Tang L, Dong LP, et al. Meta‐analysis for the association between polymorphisms in interleukin‐17A and risk of coronary artery disease. International Journal of Environmental Research and Public Health, (2016); 13(7): 660.
- Márquez A, Hernández-Rodríguez J, Cid MC, Solans R, Castañeda S, et al. Influence of the IL-17A locus in giant cell arteritis susceptibility. Annals of the rheumatic diseases, (2014); 73(9): 1742-1745.
- Geng GY, Liu HL, Zhao YJ, Wu L, Mao L, et al. Correlation between polymorphisms in the IL‐17A and IL‐17F genes and development of coronary artery disease. Genetics and Molecular Research, (2015); 14(3): 11488‐11494.
- Zheng XS, Wang S, Ni M. Association between interleukin 17A gene polymorphisms and risk of coronary artery disease. Genetics and Molecular Research, (2016); 15(1): 1-6.
- Tsai HC, Velichko S, Hung LY, Wu R. IL-17A and Th17 cells in lung inflammation: an update on the role of Th17 cell differentiation and IL-17R signaling in host defense against infection. Journal of Immunology Research, (2013); 2013(1): 267971.
- Migdalska-Sęk M, Góralska K, Jabłoński S, Kordiak J, Nawrot E, et al. Evaluation of the relationship between the IL-17A gene expression level and regulatory miRNA-9 in relation to tumor progression in patients with non-small cell lung cancer: a pilot study. Molecular Biology Reports, (2020); 47(1): 583-592.
- Anthony D, Seow HJ, Uddin M, Thompson M, Dousha L, et al. Serum amyloid A promotes lung neutrophilia by increasing IL-17A levels in the mucosa and γδ T cells. American journal of respiratory and critical care medicine, (2013); 188(2): 179-186.
- Mucciolo G, Curcio C, Roux C, Li WY, Capello M, et al. IL-17A critically shapes the transcriptional program of fibroblasts in pancreatic cancer and switches on their protumorigenic functions. Proceedings of the National Academy of Sciences, (2021); 118(6): e2020395118.
- Girondel C, Lévesque K, Langlois MJ, Pasquin S, Saba-El-Leil MK, et al. Loss of interleukin-17 receptor D promotes chronic inflammation-associated tumorigenesis. Oncogene, (2021); 40(2): 452-464.
- Song X, Wei C, Li X. The potential role and status of IL-17 family cytokines in breast cancer. International Immunopharmacology, (2021); 95(2021): 107544.
- Teijeiro A, Garrido A, Ferre A, Perna C, Djouder N. Inhibition of the IL-17A axis in adipocytes suppresses diet-induced obesity and metabolic disorders in mice. Nature Metabolism, (2021); 3(4): 496-512.
- Hristova M, Kamenarska Z, Dzhebir G, Nikolova S, Hristova R, et al. The role of IL-17 rs2275913, IL-17RC rs708567 and TGFB1 rs1800469 SNPs and IL-17A serum levels in patients with lupus nephritis. Rheumatology international, (2021); 41(12): 2205-2213.
- Wielińska J, Świerkot J, Kolossa K, Bugaj B, Chaszczewska-Markowska M, et al. Polymorphisms within Genes Coding for IL-17A and F and Their Receptor as Clinical Hallmarks in Ankylosing Spondylitis. Mediators of Inflammation, (2021); 2021(1): 3125922.
- Shao M, Xu S, Yang H, Xu W, Deng J, et al. Association between IL-17A and IL-17F gene polymorphism and susceptibility in inflammatory arthritis: A meta-analysis. Clinical Immunology, (2020); 213(2020): 108374.
- Ponce-Gallegos MA, Pérez-Rubio G, Ambrocio-Ortiz E, Partida-Zavala N, Hernández-Zenteno R, et al. Genetic variants in IL-17A and serum levels of IL-17A are associated with COPD related to tobacco smoking and biomass burning. Scientific reports, (2020); 10(1): 784.
- Tayefinasrabadi H, Mohebbi SR, Hosseini SM, Azimzadeh P, Pourhoseingholi MA, et al. Association of Interleukin-17 gene polymorphisms with susceptibility to chronic hepatitis B virus infection and clearance in Iranian population. Microbial Pathogenesis, (2020); 144(2020): 104195.
- Strauss M, Palma-Vega M, Casares-Marfil D, Bosch-Nicolau P, Lo Presti MS, et al. Genetic polymorphisms of IL-17A associated with Chagas disease: results from a meta-analysis in Latin American populations. Scientific reports, (2020); 10(1): 1-8.
- Cipolla E, Fisher AJ, Gu H, Mickler EA, Agarwal M, et al. IL‐17A deficiency mitigates bleomycin‐induced complement activation during lung fibrosis. The FASEB Journal, (2017); 31(12): 5543-5556.
- Isailovic N, Daigo K, Mantovani A, Selmi C. Interleukin-17 and innate immunity in infections and chronic inflammation. Journal of autoimmunity, (2015); 60(2015): 1-11.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0

