

Full Length Research Article
Characterization of Halophilic/Halotolerant Bacteria Isolated from the Hypersaline Environment of Khewra, District Jhelum
Ayisha Javed1, Sana Zahoor2, Muhammad Mohsin Javed2, Faisal Sheraz Shah3, Fatima Mansoor1
Adv. life sci., vol. 10, no. 1, pp. 115-121, March 2023
*– Corresponding Author: Sana Zahoor (Email: sana.zahoor@vu.edu.pk)
Authors' Affiliations
2. Virtual University of Pakistan – Pakistan
3. National University of Sciences and Technology – Pakistan
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Khewra salt mine is an extremely saline habitat of Pakistan that provides a suitable environment for salt loving microorganisms, halophiles. Halophiles grow in high salt concentration and are excellent sources of enzymes that perform catalytic reactions competently under extreme conditions.
Methods: A total of twenty-five samples (rocks, soil, water, and salt drippings), collected from Khewra salt mine and its vicinity, were checked for growth. Isolates were purified and characterized by biochemical and molecular tests. The isolates were also screened for biotechnological potential.
Results: Samples (n=12) did not show growth, while samples (n=13) showed growth on high salt media. Isolates showed growth on 1-29% NaCl concentration, 15-40 oC temperature and, pH 6.5-9.0. Molecular analysis showed that isolate AJS-21y was closely related (98% similarity) to Salinicoccus roseus while isolate AJS-22 belonged to Bacillaceae family and was closely related (99% similarity) to Piscibacillus sp. Both isolates gave extracellular amylase production.
Conclusion: Current study showed the presence of Salinicoccus sp. and Piscibacillus sp. in Khewra salt mine, Pakistan. The ability of isolates AJS-21y and AJS-22 to survive at high salt concentrations and production of extracellular amylase made them highly attractive for industrial applications and synthesizing transgenic crops tolerating high salinity.
Keywords: Khewra; Characterization; Halophilic; Halotolerant; Salinicoccus; Piscibacillus
Introduction![]()
Extremophiles are an important part of the microbial world. These microbes can tolerate a broader range of environmental extremes. Extremophiles are differentiated by their capability to endure harsh conditions like high concentration of salt, low or high temperature, and acidic or alkaline environments. They are categorized into five groups: halophiles, thermophiles, acidophiles, psychrophiles, and alkaliphiles [1].
Halophiles are microorganisms that live in areas where salt concentration is high. Salt loving microbes are present in all three domains of life i.e., Archaea, Bacteria and Eukarya. Halophiles are categorized as extreme, moderate and slight halophiles depending on the concentration of sodium chloride [2]. The sodium chloride concentration of 1.8-4.7% is optimum for the growth of slight halophiles, 4.7% to 20% concentration of sodium chloride is optimum for moderate halophiles to grow, whereas the sodium chloride concentration of 20% to 30% is suitable for the growth of extreme halophiles [3]. Microorganisms that thrive in a hypersaline environment have high amounts of inorganic nutrients and little water. The cell membrane of halophiles is composed of chromoproteins and bacteriorhodopsin, which act as a differentially permeable membrane allowing selective molecules to pass, functioning as a proton pump [2]. Halophilic microbes require nutrients in less amounts and are resistant to high concentration of salt with the ability to maintain the osmotic pressure of the environment [4]. The mechanism of halo adaptation depends on the intracellular potassium chloride storage of > 37% (salt-in technique) or aggregation of viable solutes (salt-out procedure) to keep the equalization of Na+ into the cytoplasm and balance osmotic pressure of the outside environment provided by high salt concentration [5]. Halotolerant microbes can keep up their survival in high salt conditions and also can thrive without salt. Moreover, stress tolerance proteins are present in these microbes which assist them for their survival in harsh environments [6-8].
Halophilic microorganisms maintain their osmotic balance in the cytoplasm and its surroundings by incorporating and aggregating organic solutes and manage their internal cellular environment at low ionic level [9-12]. Halophilic microorganisms are acclaimed in the oil industry because of their excessive need in the processes of oil recovery and their role in the breakdown of industrial pollutants [13, 14].
Extremophiles such as halophilic microorganisms are a gift of nature to the human race [15]. Halophilic microorganisms have incredible metabolic flexibility, less nutritional prerequisites and variation in hereditary mechanisms to survive in harsh conditions. Halophiles upgrade the feature of the environment in terms of assisting saline environments to self-regulate. It helps plants produce substances that enable their survival in such outrageous conditions. They produce various synthetics, chemicals and metabolites that are being utilized in drug, food, leather and textile industry [16].
The Khewra Salt Mine is located in District Jhelum, Punjab, Pakistan. Khewra is located north of Pind Dadan Khan [17]. Khewra mine is also called Mayo Salt Mine. It is the world’s biggest salt mine which spreads over a zone of about 3,398.53 acres. Salt ranges in color from transparent, pink, grayish, and reddish to beef-red color [18].
The present study was aimed to isolate and characterize halophilic/halotolerant bacteria from the hypersaline environment of Khewra and to determine their antibiotic resistance and biotechnological potential.
Methods![]()
Sample Collection
Samples (rocks, brine, soil and water) were collected in sterile bottles and plastic bags from the Khewra mine and were refrigerated at 4 oC till further processing.
Halophilic bacteria isolation
Halophilic bacteria were isolated through serial dilution technique. Soil suspension (1.0 ml) was taken and added to nutrient broth medium containing 15% NaCl. After incubation, turbidity (indicating growth) appeared in samples. It was further inoculated on nutrient agar plates to obtain pure growth. Pure cultures were acquired by repeated streaking on 15% NaCl nutrient agar plates followed by incubation at 37oC for 3-4 days.
Morphological Characterization
Colonies (72 h old) were examined for shape, color, elevation, margins, internal structure and configuration. Gram’s staining procedure was used to study the characteristics of single cells of the isolates.
Physiological Characterization
Effect of temperature
All isolates were tested for variable temperature range. Nutrient broth medium containing 15% sodium chloride concentration was used for the growth of isolates at different temperatures (25-40 oC) with the interval of 5oC. A single pure colony was transferred to nutrient broth (50 ml) followed by incubation at 37oC for 3-4 days. Then 50 µL of this culture was added to a 50 ml nutrient broth flask having 15% concentration of sodium chloride and incubated at 25 to 40oC for 3-4 days. The experiments were conducted in duplicate.
Effect of Sodium Chloride concentration
Effect of variable salt concentration was checked over the range of 0-30% NaCl. A single pure colony was added to 50 ml nutrient broth media containing 15% sodium chloride and incubated for 3-4 days at 37oC to get sufficient growth. Culture (50 µl) was added to a 50 ml nutrient broth flask having varied concentration of sodium chloride (0-30%) followed by incubation at 37oC for 3-4 days. The experiments were conducted in duplicate.
Effect of pH
All isolates were tested for variable pH range. Isolates were grown on high salt nutrient broth at variable pH ranging from 6.0-9.0 for 3-4 days at 37oC to get sufficient growth. The experiments were conducted in duplicate.
Biochemical Analysis
Oxidase and catalase tests were performed.
Molecular Characterization
For molecular analysis, Genomic DNA was extracted following the method of Kronstad et al. [19]. PCR amplification of 16S rRNA gene was done using primers 9F 5’-GAGGTTTGATCCTGGCTCAG-3’ and 1510R 5-GGTTACCTTGTTACGACTT-3’.
Reaction mixture was prepared by adding PCR master mix (4.0 µL,) RNase treated genomic DNA (2.0 µl), 12 µl of ddH2O water and 1 µl of forward and 1 µl of reverse primer. Initial denaturation was done at 94oC (5 minutes), followed by 30 cycles of 94oC (30 seconds), 54oC (30 seconds) and 72oC (90 seconds). One final cycle of 72oC for 10 minutes was provided for extension. 1% agarose gel was prepared for the analysis of Polymerase chain reaction products and visualized under UV in the gel documentation system. Amplified 16S rRNA gene was sequenced commercially. The homology between the strains of bacteria was analyzed through Basic Local Alignment Search Tool in GenBank. Software ClustalW was used to align experimental 16S rRNA gene sequences with 16S rRNA gene sequences (belonging to relevant taxa) [20]. Neighbor-joining method was applied for phylogenetic tree analysis utilizing MEGA version 6.0 [21]. Based on 1000 re-samplings, bootstrap analysis of neighbor-joining data was done to construct the phylogenetic tree for the study of relationship stability.
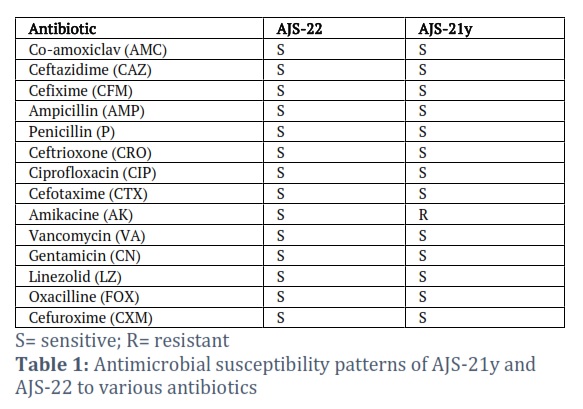
Antimicrobial Susceptibility Testing
Kirby-Bauer disk diffusion method was applied to check resistance of isolates against different antibiotics. Mueller Hinton agar was used for evaluating antimicrobial susceptibility. Amikacin, penicillin, ceftazidime, cefotaxime, ceftriaxone, cefixime, linezolid, ampicillin, ciprofloxacin, vancomycin, gentamicin, amoxicillin and oxacillin antibiotics were used to check the antimicrobial susceptibility of isolates. Incubation was done at 37°C for 72 hours. Following 72 hours of incubation, the clearance zone of diameter (mm) was noted for each antibiotic. Organism was reported as Sensitive or Resistant for each antibiotic after consulting key chart provided by CLSI 2015.
Screening for extracellular amylase activity
Starch agar medium was utilized for the screening of extracellular amylase activity (g/L: Peptone 10, yeast extract 5, starch 20, NaCl 150, and Agar 15). After incubation at 37°C for 3-5 days, iodine solution was poured on the plate, a clear zone around the growth showed starch hydrolysis.
Screening for extracellular CMCase activity
CMC agar medium was utilized for the screening of extracellular CMCase activity (g/L: Peptone 10, yeast extract 5, CMC 20, NaCl 150, and Agar 15). After incubation at 37°C for 3-5 days, Congo Red (0.1%) was added and then destained with NaCl (0.1 M). A clear zone around the growth showed the production of CMCase.
Screening for extracellular protease activity
Skim milk agar medium was utilized for the screening of extracellular protease activity (g/L: Peptone 10, yeast extract 5, Skim milk 20, NaCl 150 g, Agar 15). After incubation at 37°C for 3-5 days, formation of clear zone around the colonies indicated extracellular protease.
Accession Number of Nucleotide Sequence
The 16S rRNA gene sequence of isolates AJS-21y and AJS-22 have been submitted in the GenBank with accession no. MG471435 and MG471436, respectively.
Statistical analysis
Standard deviation was calculated using computer software SPSS (IBM, USA).
Results![]()
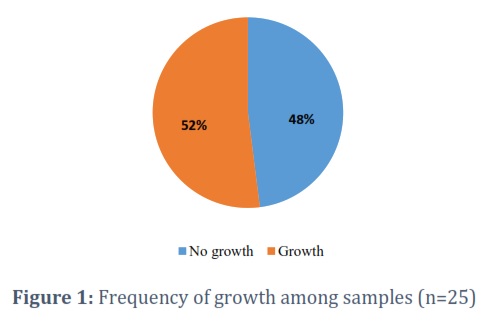
A total of 25 samples were obtained from Khewra salt mine and its surroundings. Samples were grown in 15% NaCl nutrient agar and 15% NaCl nutrient broth to obtain growth. 48% samples showed no growth while 52% of samples showed growth were further processed and characterized (Fig. 1). Samples showing no growth were excluded from further processing.
Morphological, Physiological and Biochemical characterization
Halophilic bacteria, (AJS-21y) and (AJS-22) isolated from the Khewra salt mine, were characterized in terms of morphology and physiology. Biochemical characterization of the isolates was also done. Colonies were shiny, big, and circular with entire margins for isolate AJS-21y and cells were Gram-positive cocci while reddish yellow, mucoid, small, irregular, flat and undulate colonies were observed for isolate AJS-22 and cells were Gram-positive rods. Isolate AJS-21y was oxidase and catalase positive while isolate AJS-22 was catalase and oxidase negative.
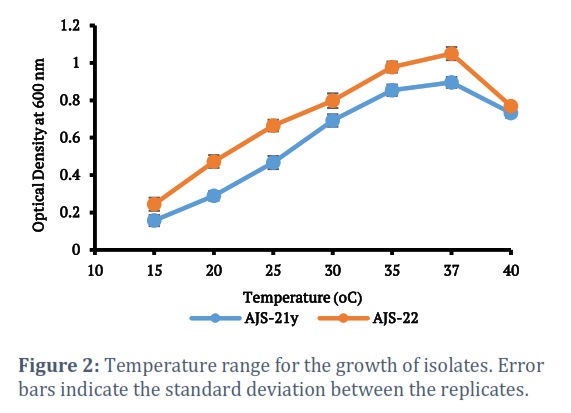
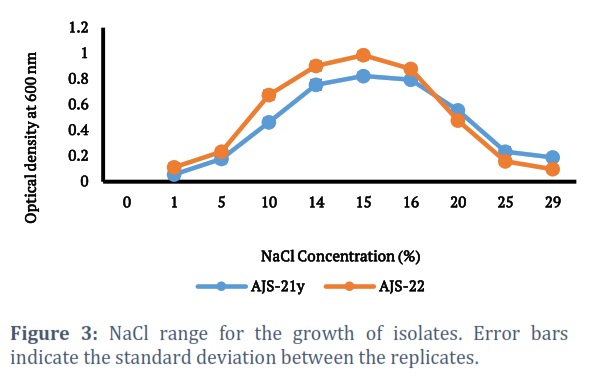
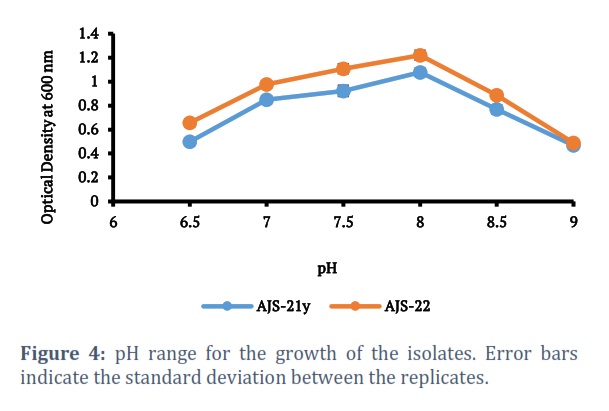
Isolates AJS-21y and AJS-22 were evaluated for growth at variable temperature ranges. At 10oC or temperatures lower than 10oC, no growth was observed. Isolates showed growth within the temperature range of 15oC-40oC (Fig. 2). It took 72 hours for isolates to give maximum growth at 37oC, which is a known regular time period for growth of halophiles. Isolates were also examined for growth at variable salt concentration (0-30%). Maximum growth of isolates, AJS-21Y and AJS-22, was obtained at 15% sodium chloride concentration. At 30% sodium chloride concentration, no growth was observed (Fig. 3). The growth of the isolates, AJS-21y and AJS-22, was checked at pH ranging from 6.0 to 9.0. No growth was observed at pH 6.0 while both isolates showed growth at pH 6.5-9.0. pH 8.0 was found optimum for growth of both isolates (Fig. 4).
Molecular Characterization
DNA was isolated and 16S rRNA gene of the isolates (AJS-21y and AJS-22) was amplified using 9F and 1510R primers. 16S rRNA gene was sequenced commercially. Phylogenetic analysis showed that isolate AJS-21y was 98% identical to Salinicoccus roseus and isolate AJS-22 was 99% similar to Piscibacillus species. Sequences of isolates AJS-21y and AJS-22 and sequences of their closely related species were aligned using ClustalW and MEGA 6.0 was used to construct phylogenetic tree of isolates and related organisms, which depicted that isolate AJS-21y was closely related to Salinicoccus roseus while isolate AJS-22 was closely related to Piscibacillus sp. (Fig. 5&6).
Antimicrobial susceptibility and Enzyme production
Results of the antimicrobial susceptibility testing of the isolates revealed that the isolate AJS-22 was sensitive to all the antibiotics tested while isolate AJS-21y was resistant against Amikacin (Table 1). Enzyme screening of isolates (AJS-21y and AJS-22) to produce extracellular protease, CMCase and amylase showed that both isolates AJS-21y and AJS-22 gave positive results for amylase production but gave negative results for extracellular protease and CMCase.
Figures & Tables


In the ongoing years, saline soils have received considerable attention because of the deficiency of arable land and the expanding need of rebuilding efforts of regions influenced by secondary saltiness [22]. Halophilic bacteria are abundant in hypersaline environment and possess an extraordinary biotechnological potential that can be utilized [23,24].
Twenty-five different sites at Khewra salt mines and its vicinity were selected for sample collection (soil, rocks, water and salt drippings). Out of 25 samples, thirteen samples (52%) showed growth.Isolate AJS-21y formed shiny, big, and circular colonies while isolate AJS-22 formed reddish yellow, mucoid, small and irregular colonies. Cells of both isolates were Gram-positive in nature. Studies have shown that Gram-positive bacteria belonging to the genus Bacillus were largely found in saline habitats [25]. Pigmented colonies for the halophilic bacteria have been reported by various scientists.Colonial pigmentation of the halophilic bacteria isolated from Khewra salt mines included orange, yellow, red, cream colored, and white [26]. Sarwar et al. isolated 7Gram positive and 2 Gram negative bacteria from saline waters of Khewra salt mines [27].
Isolates AJS-21y and AJS-22 showed optimum growth at 37oC while no growth was obtained below 15oC.This study is in accordance with the study of Akhtar et al. who reported 37 ºC as the optimum temperature for the growth of halophilic isolates obtained from Khewra Salt mines [26]. In contrast, some halophilic bacterial strains isolated from rhizospheric and non-rhizospheric soils of Khewra tolerated 4 °C and some showed better growth at 42 °C [28]. Roohi et al. isolated halophilic bacteria from the Karak Salt mines that showed optimal growth within a temperature range of 28-37 oC [29]. Halophilic bacteria isolated from the lake of Urmia showed growth within the temperature range of 25-35 oC, that is close to the temperature range of halophilic bacteria isolated in this study [30]. Salinicoccus roseus strains showed growth within the temperature range of 20-45 oC with optimal growth observed at 37oC [31].
Isolates AJS-21y and AJS-22 showed growth within the NaCl range of (1-29%) with optimum growth observed at 15%. The halophilic bacteria isolated in this study showed growth up to 29% NaCl concentration which is more than the NaCl concentration reported for the growth of most of the halophilic bacteria from Khewra salt mines in the previous studies. Laraibet al. isolated halophilic bacteria from Khewra Salt mines that showed growth within the NaCl range of 3-15% [32] while study of Sarwaret al. and Mukhtaret al. reported NaCl range of 1.5–3.0 M for the growth of the halophilic isolatesfrom Khewra [27, 28]. Similar results were also obtained from Urmia Lake, where halophilic isolates grew at sodium chloride concentration (7-20%) [30]. Salinicoccus roseus strains depicted growth within the range of 5-15% (w/v) NaCl concentration with optimum growth observed at 10% for S. roseus rvscokh1 and 6% for S. roseus rsk1 [31].
Isolates AJS-21y and AJS-22 showed growth within the pH range of 6.5-9.0 with optimal growth at pH 8.0. This is in accordance to the findings of Mukhtaretal., who reported pH 8.0 as the optimum pH for the growth of the most of the halophilic strains isolated from Khewra salt mines while in contrast, some of the isolates showed growth at pH as low as 4.0 [28]. Bacillus megaterium CTBmeg1 showed growth within pH range of 7.0–9.0 [33]. Isolates obtained from Urmia Lake showed growth with variable pH range of 6.0-9.0 [30]. Salinicoccus roseus strains showed growth within the pH range of 6.0-9.0 with optimal growth observed at pH 7.4 [31].
16S rRNA gene analysis of isolates showed that AJS-21y and AJS-22 were closely related to Salinococcus sp. and Piscibacillus sp. respectively. Presence of Salinococcus sp. in halophilic environments has also been reported by various scientists [31, 36]. Piscibacillushalophilus sp., was isolated from a hypersaline Iranian lake.It was a moderately halophilic bacterium that formed mucoid, flat, irregular colonies while the cells were Gram positive non-motile spore forming rods. It was oxidase and catalase positive [22]. Salinicoccus roseus was a moderately halophilic non-spore forming bacterium that formed circular colonies with Gram positive cells [37]. Salinococcus sp. and Picibacillus sp. obtained in this study were not reported in the previous studies conducted at Khewra salt mines. Previously, Bacillus megaterium, Bacillus tequilensis, Bacillus xiamenensis, Bacillus anthracis, Staphylococcus saprophyticus, Pseudomonas putida, Pseudomonas aeruginosa, Staphylococcus pasteuri, Halobacillus, Escherichia coli, Staphylococcus arlettae, Staphylococcus gallinarum and Kocuria sp. have been reported at Khewra Salt mines by different scientists [26-28, 37].
Isolate AJS-22 was found to be sensitive to all the antibiotics tested in this investigation while isolate AJS-21y was resistant against Amikacin. Kushneria pakistanensis NCCP-934T isolated from Karak salt Mines, Pakistan was found to be sensitive against Penicillin and resistant against Oxacillin [38] while Bacillus pakistanensis NCCP-168T also isolated from the Karak salt mines was sensitive against Oxacillin and resistant against Penicillin [39]. Shinde and Thombre found that marine halophilic isolates (RR2, RR3 and RR5) were sensitive to Gentamicin, while they were resistant to Ampicillin, Bacitracin, Ciprofloxacin, Chloramphenicol and Tetracycline. Isolate RR1 was sensitive to Novobiocin and Trimethoprim [23].
Halophiles are rich sources of hydrolytic enzymes. Isolates were checked for the production of extracellular amylase, CMCase and protease. Isolates AJS-21y and AJS-22 gave extracellular production of amylase. Amylases are chief components of industries likefood, fermentation and pharmaceutical industries. Halophilic amylase offers an added potential of being salt tolerant. Production of protease, amylase, and cellulase/CMCasebyhalophilicisolates from Khewra salt mines have also been reported by different scientists [26, 28, 40]. Halophilic bacteria isolated from hyper saline lake in Tunisia produced amylase, protease and cellulase [41]. Salinicoccus roseus strain gave positive results for amylase production [31]. Halophilic strain LY9 isolated from soil samples collected from Yuncheng, China showed amylase activity [42]. Twenty-two halophilic strains isolated from Indian salterns produced amylase while twelve isolates produced cellulase and ten isolates produced protease [43].
Author Contributions
Conception/ design of the work: Muhammad MohsinJaved
Sample collection: Muhammad MohsinJaved, Sana Zahoor, AyishaJaved, Fatima Mansoor
Experimental work: AyishaJaved, Fatima Mansoor
Data analysis and interpretation: Sana Zahoor, AyishaJaved
Drafting the article: AyishaJaved, Sana Zahoor, Faisal Sheraz Shah
Critical revision of the article: Sana Zahoor
The authors declare that there is no conflict of interest.
![]()
References
- Babu P, Chandel AK, Singh OV. Extremophiles and their applications in medical processes. New York, NY (2015) Springer International Publishing
- Sarwar MK, Azam I, Iqbal T. Biology and Applications of Halophilic Bacteria and Archaea: A. Electronic Journal of Biology, (2015); 11(3): 98-103.
- Ollivier B, Caumette P, Garcia JL, Mah RA. Anaerobic bacteria from hypersaline environments. Microbiology and Molecular Biology Reviews, (1994); 58(1): 27-38.
- Martin S, Marquez MC, Sanchez-Porro C, Mellado E, Arahal DR, Ventosa A. Marinobacter lipolyticus sp. nov., a novel moderate halophile with lipolytic activity. Journal of Medical Microbiology, (2003); 53(5): 1383-87.
- Bremer E, Kramer R. Responses of microorganisms to osmotic stress. Annual review of Microbiology, (2019);73: 313-334.
- Ara I, Daram D, Baljinova T, Yamamura H, Hozzein WN, Bakir MA, Suto M. Isolation, classification, phylogenetic analysis and scanning electron microscopy of halophilic, halotolerant and alkaliphilic actinomycetes isolated from hypersaline soil. African Journal of Microbiology Research, (2013); 7(4): 298-308.
- Hillmann F, Fischer RJ, Bahl H. The rubrerythrin-like protein Hsp21 of Clostridium acetobutylicum is a general stress protein. Archives of Microbiology, (2006); 185(4): 270-276.
- Kapardar RK, Ranjan R, Grover A, Puri M, Sharma R. Identification and characterization of genes conferring salt tolerance to Escherichia coli from pond water metagenome. Bioresource Technology, (2010); 101(11): 3917-24.
- De Bruijn FJ. Use of repetitive (repetitive extragenic palindromic and enterobacterial repetitive intergeneric consensus) sequences and the polymerase chain reaction to fingerprint the genomes of Rhizobium meliloti isolates and other soil bacteria. Applied and Environmental Microbiology, (1992); 58(7): 2180-87.
- Ghasemi Y, Rsoul-Amini S, Ebrahiminezhad A, Kazemi A, Shahbazi M, Talebnia N. Screening and isolation of extracellular protease producing bacteria from the Maharloo Salt Lake. Iranian Journal of Pharmaceutical Sciences, (2011); 7(3): 175-180.
- Ghasemi Y, Rasoul-Amini S, Kazemi A, Zarrini G, Morowvat MH, Kargar M. Isolation and characterization of some moderately halophilic bacteria with lipase activity. Microbiology, (2011); 80(4): 483-7.
- Oren A. Bioenergetic aspects of halophilism. Microbiology and Molecular Biology Reviews, (1999); 63(2): 334-48.
- Birbir M, Ilgaz A. Isolation and identification of bacteria adversely affecting hide and leather quality. Journal of the Society of Leather Technologies and Chemists, (1996); 80:147-153.
- Ventosa A, Nieto JJ. Biotechnological applications and potentialities of halophilic microorganisms, World Journal of Microbiology and Biotechnology, (1995); 1(1): 85-94.
- Ventosa A, Nieto JJ, Oren A. Biology of moderately halophilic aerobic bacteria. Microbiology and Molecular Biology Reviews, (1998); 62(2): 504-44.
- Charlesworth J, Burns BP. Extremophilic adaptations and biotechnological applications in diverse environments. AIMS Microbiology, (2016); 2(3): 251-61.
- CamerapixSpectrum Guide to Pakistan. (1998). Interlink Books. p. 150.
- Hussain MT, Sharif QM, Hussain M. Chemical evaluation of major salt deposits of Pakistan. Journal of the Chemical Society of Pakistan, (2011); 29(6): 569.
- Kronstad JW, Schnepf HE, Whiteley HR. Diversity of locations for Bacillus thuringiensis crystal protein genes. Journal of Bacteriology, (1983); 154(1): 419-28.
- Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic acids research, (1994); 22(22): 4673-80.
- Tamura K, Peterson D, Peterson N, Stecher G, Nei M, Kumar S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Molecular Biology and Evolution, (2011); 28(10): 2731-9.
- Amoozegar MA, Sánchez-Porro C, Rohban R, Hajighasemi M, Ventosa A. Piscibacillus halophilus sp. nov., a moderately halophilic bacterium from a hypersaline Iranian lake. International Journal of Systematic and Evolutionary Microbiology, (2009); 59(12): 3095-99.
- Shinde V, Thombre RS. Antibiotic resistance profiling of marine halophilic bacteria and haloarchaea. Journal of Applied Pharmaceutical Science, (2016); 6(10): 132-7.
- Canfora L, Bacci G, Pinzari F, Papa GL, Dazzi C, Benedetti A. Salinity and bacterial diversity: to what extent does the concentration of salt affect the bacterial community in a saline soil? PLoS One, (2014); 9(9): e106662.
- AbdSamad NS, Amid A, Jimat DN, Shukor NA. Isolation and identification of halophilic bacteria producing halotolerant protease. Science Heritage Journal, (2017); 1: 7-9.
- Akhtar N, Ghauri, MA, Iqbal, A, Anwar, MA, Akhtar, K. Biodiversity and phylogenetic analysis of culturable bacteria indigenous to Khewra salt mine of Pakistan and their industrial importance. Brazilian Journal of Microbiology, (2008); 39: 143-150.
- Sarwar MK, Anwer F, Rashda A, Azam I, Iqbal T, Nosheen A, Azam WI. Isolation and characterization of halophilic bacterial strains from saline waters of Khewra salt mines on the basis of 16S rRNA gene sequence. International Journal of Applied Biology and Pharmaceutical Technology, (2014); 5: 142-149.
- Mukhtar S, Mehnaz S, Mirza MS, Malik KA. Isolation and characterization of bacteria associated with the rhizosphere of halophytes (Salsolastocksii and Atriplexamnicola) for production of hydrolytic enzymes. Brazilian Journal of Microbiology, (2019);50(1): 85-97.
- Roohi A, Ahmed I, Khalid N, Iqbal M, Jamil M. Isolation and phylogenetic identification of halotolerant/halophilic bacteria from the salt mines of Karak, Pakistan. International Journal of Agriculture and Biology, (2014); 16(3): 564-70.
- Vahed SZ, Forouhandeh H, Hassanzadeh S, Klenk HP, Hejazi MA, Hejazi MS. Isolation and characterization of halophilic bacteria from Urmia Lake in Iran. Microbiology, (2011); 80(6): 834-41.
- Ravi RK, Patel P. Isolation and characterization of hydrolytic enzyme producing halophilic bacteria Salinicoccus roseus from Okha. International Journal of Microbiology Research, (2015); 7(2): 616-622.
- Laraib, F, Rauf, R, Dilshad, R, Jameel, N, Batool, R. Characterization of Halophilic Bacteria Isolated from Khewra Salt Mines. Lahore Garrison University Journal of Life Sciences, (2022); 6(02): 133-147.
- Akcay K, Kaya Y. Isolation, characterization and molecular identification of a halotolerantBacillus megaterium CTBmeg1 able to grow on halogenated compounds. Biotechnology and Biotechnological Equipment, (2019); 33(1): 945-53.
- Diba H, Cohan RA, Salimian M, Mirjani R, Soleimani M, Khodabakhsh F. Isolation and characterization of halophilic bacteria with the ability of heavy metal bioremediation and nanoparticle synthesis from Khara salt lake in Iran. Archives of Microbiology, (2021); 7: 3893-3903.
- Flores MR, Ordonez OF, Maldonado MJ, Farias ME. Isolation of UV-B resistant bacteria from two high altitude Andean lakes (4,400 m) with saline and non-saline conditions. The Journal of General and Applied Microbiology, (2009); 55(6): 447-58.
- Ventosa A, Marquez MC, Ruiz-Berraquero F, Kocur M. Salinicoccus roseus gen. nov., sp. nov., a new moderately halophilic Gram-positive coccus. Systematic and Applied Microbiology, (1990); 13(1): 29-33.
- Haroon U, Liaquat F, Khizar M, Akbar M, Saleem H, Arif S, Ali W, Chaudhary HJ, Munis M FH. Isolation of halotolerant bacteria from rhizosphere of Khewra salt mine halophytes and their application to induce salt tolerance in wheat. Geomicrobiology Journal, (2021); 38(9): 768-775.
- Bangash A, Ahmed I, Abbas S, Kudo T, Shahzad A, Fujiwara T, Ohkuma M. Kushneria pakistanensissp. nov., a novel moderately halophilic bacterium isolated from rhizosphere of a plant (Saccharumspontaneum) growing in salt mines of the Karak area in Pakistan. Antonie van Leeuwenhoek, (2015); 107(4): 991-1000.
- Roohi A, Ahmed I, Paek J, Sin Y, Abbas S, Jamil M, Chang YH. Bacillus pakistanensis sp. nov., a halotolerant bacterium isolated from salt mines of the Karak Area in Pakistan. Antonie van Leeuwenhoek, (2014); 105(6), 1163-1172.
- Leena MC, Aamer AS, Abdul H, Fariha H. Physiological, biochemical and phylogenetic characterization of extremely halophilic bacteria isolated from Khewra mine, Pakistan. Applied Ecology and Environmental Research, (2018); 16(2): 1243-1256.
- Karray F, Ben Abdallah M, Kallel N, Hamza M, Fakhfakh M, Sayadi, S. Extracellular hydrolytic enzymes produced by halophilic bacteria and archaea isolated from hypersaline lake. Molecular Biology Reports, (2018); 45(5): 1297-1309.
- Li X, Yu HY. Characterization of an organic solvent-tolerant α-amylase from a halophilic isolate, Thalassobacillus sp. LY18. Folia Microbiologica, (2012); 57(5): 447-53.
- Rathakrishnan, D, Gopalan, A. K. Isolation and characterization of halophilic isolates from Indian salterns and their screening for production of hydrolytic enzymes. Environmental Challenges, (2022); 6: 100426.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0