

![]()
Gene Profiling for Invertase Activity: Assessment of Potato Varieties for Resistance towards Cold Induced Sweetening
Arfan Ali1*, Mazhar Iqbal2, Qurban Ali3, Abdul Razzaq4, Idrees Ahmad Nasir3
Adv. life sci., vol. 3, no. 2, pp. 63-70, February 2016
*- Corresponding Author: Arfan Ali (Email: Arfan.Ali@cemb.edu.pk)
Authors' Affiliation
2- Department of Environmental Sciences, Faculty of Biological Sciences, Quaid-i-Azam University, Islamabad – Pakistan
3- Centre of Excellence in Molecular Biology, University of the Punjab – Pakistan
4- University of Central Punjab – Pakistan
Abstract![]()
Introduction
Methods
Results
Discussion
Supplementary Data
References
Abstract
Background: Potato is the most important staple food in the world. Cold-induced sweetening occurs when potatoes are stored at low temperature for longer period of time. Due to non- enzymatic Millard reaction it causes unwanted changes in colour, taste and in flavor when fried and roasted at high temperature. However, long-term cold storage is mandatory to keep an adequate supply of potatoes throughout the year. The cause of cold-induced sweetening is invertase enzyme.
Methods: Five potato varieties (Hermes (A) Lady Rosetta (B) Oscar (C) Kuroda (D) and Multa (E)) were investigated for invertase activity during two month cold storage at 4°C. Crude protein was extracted by PD Midi Trap G25 column technique. Quantification of mRNA expression was employed through QPCR. Determination of sucrose, reducing sugars and organic acids was simply done by 80% ethanol method and concentration were find out by using HPLC with already set standards. The correlation between invertase enzyme, sugar content and mRNA expression was calculated through Statistical methods.
Results: Significant activity of invertase was observed at 4ºC with up to 6.3 nmol/min/mg of protein in the type-1 & 4 (cv. Hermes and Kuroda); 2.5 times less in type 2 (Rosetta) and 3.5 times less in type 3 (Multa) when compared with same at 4ºC. In addition, malic acid concentration was found positively correlated with invertase activity at 4ºC as compared with its concentration at harvesting time. However, citric acid and oxalic acid concentrations were independent of invertase enzyme activity. The transcript level of invertase enzyme was found significantly high in potato tubers stored at 4ºC in result 1 & 4 type, less in result type 2(C) and negligible in result 3(E) potato variety when revealed through reverse transcription PCR.
Conclusions: In conclusion, Oscar (C) and Multa (E) were found more resistant to CIS at 4ºC storage and may be used for future variety improvement programs for CIS resistant through breeding and molecular approaches.
Keywords: Potato, Cold induced sweetening, Invertase, reducing sugars, Solanum tuberosum, Metabolism
Introduction
Potato has turned into a very important crop for both consumers and farmers in Pakistan. All over the world Pakistan stands fourth in production of potato. It is extremely nutritious and high yielding crop. Potatoes are usually harvested at rest period or even after the end of that period. Poor post-harvest handling including improper storage practices and transport that cause abounding damage and abridgement of burning quality. The potentially damaging factor among above listed is the improper storage. Potato needs metabolic stability with respect to its carbohydrate contents for un-interrupted supply to the industry all the year around. Low temperature storage is mandatory as it diminishes bacterial soft rots, decrease water, dry matter loss and prevents sprouting [1]. Sowokinos reported that storage at low temperature triggers increase in reducing sugar content of the potato termed as “low temperature sweetening” [2]. It is reported that reducing sugars react with free amino acids by the Millard reaction and a neurotoxin acrylamide accumulates during this process [3]. This undesired increase in reducing sugars of potato severely affects its market demand by end user.
Accumulation of hexose sugars in potato during cold storage at 4ºC for two months is done by the enzyme invertase. Correlation exists between hexose accumulation, acid invertase activity and sucrose ratio in cultivars exhibiting varying resistance to cold-induced sweetening [4-7]. At low temperature storage, the increase in activity of invertase is cultivar dependent [8]. Presumably differences in mRNA expression patterns and post-transcriptional events determine the degree of particular cultivar’s resistance to cold- induced sweetening [7,8]. Certain reports exist where hexose accumulation was prevented during cold storage in transgenic plants with suppressed acid invertase activity [9]. In this process, the activity of vacuolar invertase (StvacINV1) and the role of invertase inhibitors are considered equally important [10,11].
The current study focused on identification of elite potato varieties from Pakistan with resistance towards low temperature induced sweetening based on vacoular acid invertase enzyme activity. Here we used five potato varieties; Hermes (A) Lady Rosetta (B) Oscar (C) Kuroda (D) and Multa (E). Vacuolar invertase activity and sugar contents of each variety’s tubers were recorded during storage period at 4ºC, 10ºC and at 20ºC. Similar parameters were also tabulated during growing stage for each variety under study. The correlation between invertase enzyme activity, sucrose and reducing sugar content is highlighted. This study is one of its kinds regarding determination of relevance of Invertase mRNA expression level and reducing sugars.
Methods
Plant Material
Five local potato varieties; Hermes (A) Lady Rosetta (B) Oscar (C) Kuroda (D) and Multa (E) were kindly provided by Potato Research Institute Sahiwal, Pakistan. Potato varieties with significant agronomical potential were selected. All-inclusive five potato varieties were harvested through three successive years (2010-12) in the Centre of Excellence in Molecular Biology, University of the Punjab, Lahore fields. All the required agronomic practices including fungicide, herbicide, insecticides and fertilizers were applied to obtain better yield output. The harvested tubers were cured at 20ºC for 15 days in the dark. Fresh, good skinned and healthy looking tubers weighing between 100-120 g were selected and were stored at 20ºC, 10ºC and at 4ºC in the dark with 90% relative humidity. For analysis of sugar content at growing stage, three immature tubers weighing between 75-100 g were selected while post harvest analyses were performed on five randomly selected potatoes.
Sample preparation & Extraction of Crude Protein
Longitudinal sections of subjected potato tubers were made to measure amount of sucrose, reducing sugar, organic acid and invertase enzyme activity assay. The samples were snap frozen with liquid nitrogen. For invertase enzyme activity assay, total tuber protein was isolated. Approximately 100 g of the grinded tuber sample was used for crude protein extraction. PD Midi Trap G25 column cat#45-001-530 was used for this task. Extraction buffer was prepared by using 50mM HEPES-KOH, pH7.5, 5mM MgCl2, H2O, 1mM EDTA, 1mM EGTA, 0.1% W/V Triton X-100 and 10% W/V glycerol. PMSF was added just prior to use at the final concentration of 1mM. Further extraction of crude protein was done as per manufacturer’s instructions. Total crude protein was estimated by Bradford assay [12].
Vacoular acid invertase enzyme activity
Invertase enzyme activity was measured with and without invertase inhibitor as originally described by Liu [13]. The assay was based on the principle that the crude protein extract of potato has an endogenous proteinaceous inhibitor which has an inhibitory effect on soluble invertase activity [14]. Each sample comprised of 20 µg of total protein. Negative control samples were also processed in parallel. Final absorbance was taken at 560 nm by spectrophotometer after putting the reaction mix plate in dark for 30 minutes. Activity was calculated as nmol glucose formed per minute per mg of protein.
Determination of sucrose, reducing sugars and organic acids
Potato tuber samples were subjected to lyophilisation for two days prior to snap freezing in liquid nitrogen. After centrifugation, samples were resuspended in 4 mL of 80% ethanol followed by vortexing for 1 min. Further, samples were incubated at 60oC with continuous shaking overnight. Subsequently, supernatant was transferred into new tube and centrifuged at 4000 rpm and 20oC for 10 min. The previous step was repeated once. Further, 80% ethanol was added in the supernatant to make volume up-to 10ml. Subsequently, the samples were subjected to vacuum-fuge for drying without heat for overnight. For HPLC analysis, 1ml of mQ H2O was added and incubated at room temperature for 30 min. The pellet was dissolved through vortex and centrifuged at 13000 rpm for 10 minutes. Filtration of samples was done with the help of tuberculin syringe, by using 0.2 µm regenerated cellulose syringe filter. Glucose, fructose, and sucrose were calculated in mg/ml along with Malic acid, Citric acid and Oxalic acid through High Performance Liquid Chromatography (HPLC).
Real Time PCR analysis
Total cellular RNA was extracted from snap frozen tuber slices by using Agilent Plant RNA isolation Mini Kit (Cat#5155-2780). RNA was treated with DNase (Ambion DNased treatment kit Cat#AM1906). The cDNA was prepared by using Superscript III (Invitrogen, cat18080-051) as per instructions. This cDNA was used as template for real-time PCR assay. Relative quantification of mRNA (invertase gene) was done with Bio-Rad PCR system. SYBR Green/fluorescein qPCR Master mix (Cat#k0241) was used. Data were normalized by actin as internal control. Cycling profile was 95oC for 10 min, followed by 95oC for 15s, 55oC for 30s and 72oC for 30s.
Statistical analysis
The deduced data was statistically analysed by using analysis of Variance technique [15] whereas heritability [16] and genetic advance was analysed according to Falconer [17].
Results
Estimation of sugar content in potato tuber at growing period
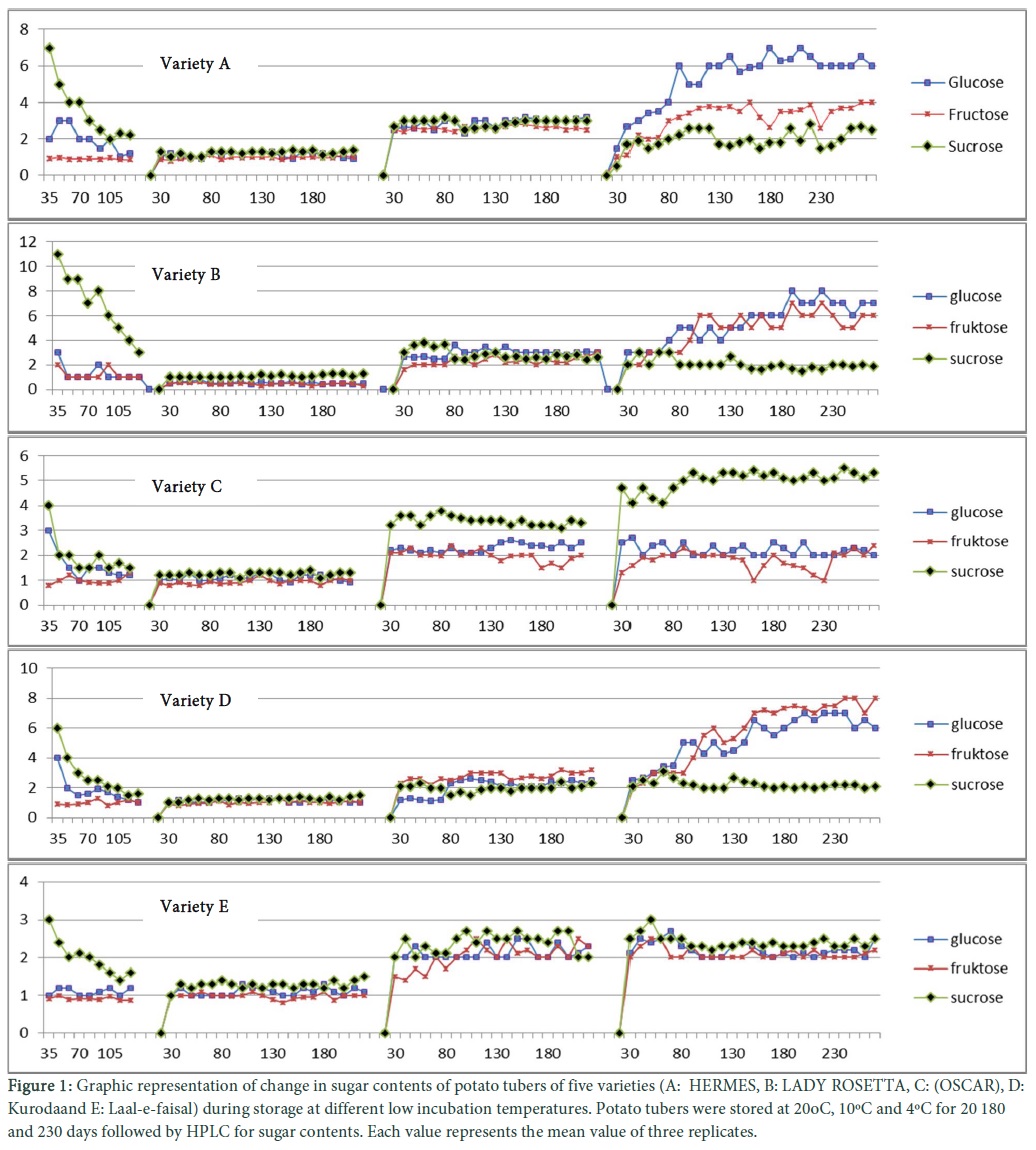
Maximum 5 nmol/mL of glucose content was obtained in 35 days old immature potato tubers. Fructose was present less than 1 nmol/mL and it was found to be almost constant in all the cultivars. Sucrose content ranged from 4-11 nmol/mg of fresh biomass of tuber which was higher than glucose at early growing stages. However, amount of glucose and sucrose was almost same at harvesting stage. Before the end of the growing season the minimum contents of reducing sugars in tubers were reached and deliberate addition occurred hereafter [18]. Similar changing trend in sugars were obtained in this research (Fig. 1 extreme left).
Estimation of sugar content at different storage time and temperatures
Less than 1.5 nmol/mg of sugar contents of fresh weight were obtained in almost all tubers among all varieties when analysed during dormancy stage, depicting low metabolic activities during this period. Beside this a little increase in the sucrose contents was observed in all varieties during sprouting. Sugar contents of both varieties A and B were same with low glucose and sucrose contents initially. But after 35 days of first analysis, glucose content were increased to 3 nmol/mg as compared to sucrose and fructose which were 2.7 nmol/mg and 2.4 nmol/mg, respectively at 10ºC. While variety C showed high sucrose contents (3.5 mg/g) throughout sampling period. Results of variety D were interesting in a sense that instead of sucrose (2 nmol/mg) and glucose (2.7 nmol/mg), fructose (4 nmol/mg) was high throughout sampling dates from initial to ending period. Variety E showed no prominent fluctuation from normal as its sucrose was high (2.6 nmol/mg) but it was low as compared to other varieties (4 nmol/mg). Four different patterns of glucose, fructose and sucrose contents were observed in all five potato varieties when subjected to storage at 4oC i.e. (1) increased reducing sugars level during storage; (2) increase in sucrose level but not reducing sugars; (3) increase fructose level as compared to glucose and sucrose; (4) low level of sucrose as well as reducing sugars. In result-1 category (variety A and D) sucrose and reducing sugars were 2 mg/g Fresh weight (FW) initially during storage period of 30-80 days. There was observed an increase in reducing sugar level up to 6.5 mg/g/ FW in the same varieties during storage period of 180 days. The levels of reducing sugars increased gradually up to 7 mg/g FW in variety A while 8 mg/g FW in variety D after storage period of 230 days. However in result-2 category, in variety (Oscar C) sucrose contents were high in initial sampling that is almost 5 mg/g of FW. It reached at maximum level of 5.8 mg/g FW after 180 days of storages and then remained stable. Interestingly, levels of reducing sugar in this cultivar (C) remained stable. In variety D the pattern was quite similar to type 1, but the only difference which was noticed is that instead of glucose, fructose was high (8 mg/g FW) until last days of storage and remained stable.
In result-3 category, variety E showed unique pattern. Sucrose contents increased rapidly and reached to its maximum level within 5 weeks (about 3 mg/g FW), but then decreased. The reducing sugar were also increased during that period, but later on decrease in reducing sugars was also observed until 2 mg/g fresh weight. A slight increase in sucrose was also observed at this stage and after that it became stable. The cultivar E is the progeny of ND 860-2, which is a so-called, “low sugar-accumulating, cold-tolerant” breeding line. These results are well correspond to Espen [19]. Corresponding with the results described by (Burton. 1989) type1 & 4, results are resembling with Hill [20]. The results of cultivar E are similar to those observed in studies of cold-tolerant lines, ND 860-2 (Fig. 1) [6].
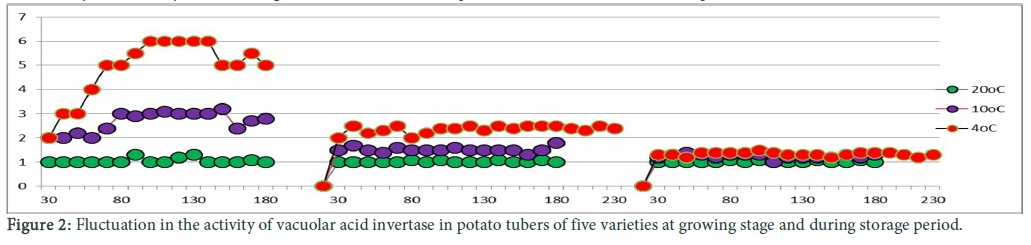
Variation in enzyme activity
Three types of vacoular acid invertase activities were observed which represents four types of sugar contents of five varieties. During the growing season the activity of acid invertase was observed to be 2.5 to 3 nmol glucose/min/mg. Decrease in acid invertase activity was observed which attained its lowest level till the harvesting time. Similar behaviour of all the varieties was seen during growing season. In the Fig. 2 activity of acid invertase enzyme represents the sugar contents behaviours of all the varieties at curing period which demonstrate that the activity of acid invertase enzyme was low as in the growing season but fluctuation in acid invertase enzyme activity was observed at 10oC in the similar pattern as was the case with sugar contents at this stage for all these varieties. Vacoular acid invertase activity was 3.5 nmol glucose/min/mg at 10oC storage period, but at 4oC their gradual increase in the activity of vacuolar acid invertase (VINV) was observed which reached to its maximum level of 6.3 nmol glucose /min/mg. Then it attained a constant level and slight decrease was observed after 180 days of storage after that stability was attained.
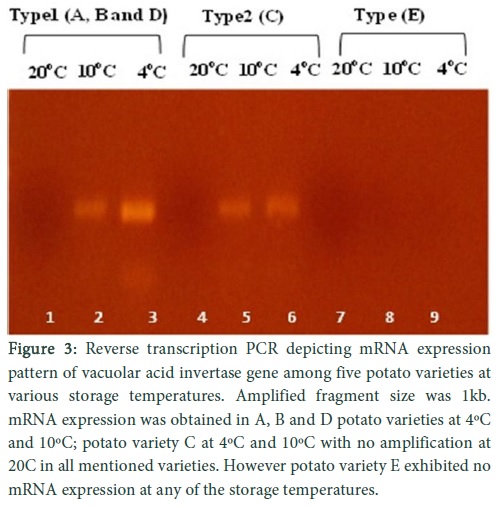
Relative transcript levels of vacuolar acid invertase in potato tubers
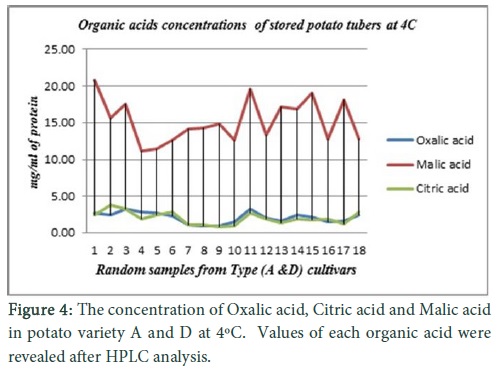
In cultivar (A , B and D), very low signals were observed during real time quantification of vacoular acid invertase gene in potato tubers stored at 10oC, but increase in transcriptional level was observed at 4ºC. A very medium to low transcriptional level for vacuolar acid invertase gene was observed for cultivar C at 10oC as well as at 4oC storage of potato tubers. Vacoular acid invertase gene activity was negligible for variety (E) when analysed through real time quantification for its transcriptional level at 10oC as well at 4oC (Fig. 3). Random samples from varieties (A & D) were picked for organic acid analysis because these varieties were mostly affected by invertase activity and protein was isolated from stored potato tubers (4oC). Protein samples were subjected to HPLC and concentrations of malic acid (20 mg/ml of protein) was found high as compared to citric acid (4 mg/mL) and oxalic acid (3 mg/mL).
Statistical analysis
Significant genetic differences were reported among potato genotypes at growth stage and under different temperature treatments (Table 1 in supplementary data). It was found that maximum amount of sucrose (4.38±0.8482 mg) was recorded at growth stage of potato as compared with glucose and fructose. Highest heritability (81.47%) and genetic advance (136.446%) was observed for sucrose while lower heritability and higher genetic advance was found for glucose and fructose. Heritability (58.665%, 62.679%) and genetic advance (54.137%, 16.984%) was higher for fructose in the potato tubers at 10°C and 4°C temperature storage, respectively. At 20°C higher amount of sucrose (1.1667±0.0586 mg) was found as compared with glucose and fructose amounts while higher heritability (82.852%, 85.259%) and genetic advance (84.050%, 89.679%) were found for fructose and glucose, respectively. Table 2 (in supplementary data) shows that higher amount of fructose (1.2222 mg) and sucrose (6.8889 mg) was recorded for Multa while lowest fructose (0.9087 mg) and sucrose (1.9889 mg) was recorded from Multa (E) at growth stage of potato. Higher glucose (1.9667 mg) was found for Hermes (A) while lowest was found for Multa (E) at growth stage of potato. It was persuaded from results that higher amount of fructose was found for Oscar (B) while lowest for Multa and Kuroda (D) at 20, 10 and 4°C temperature storage. Higher amount of glucose was found in Oscar (C) and Kuroda (D) at 4°C, while lowest glucose amount was reported for Multa (E) at 4°C temperature storage. Higher amount of sucrose was found for Multa at 4°C, 10°C while lowest for Oscar (C) and Kuroda (D). Principle component Bi-plot (Fig. 1) suggested that over all better performance for sucrose was observed by Multa at 4oC storage. Higher amount of fructose, glucose and low sucrose was reported for Lady Rosetta (B) and Hermes (A) (Fig. 2). It can be inferred from figure 3 that higher amount of sucrose was found for Multa, Glucose for kuroda (A & D) while the other genotype showed higher fructose contents.
Data and Tables
Discussion
An effort was made to monitor the sugar contents and vacoular acid invertase enzyme activity in young tubers in local potato cultivars as done by Endo [4] in Japanese potato cultivars. The contents of sucrose and glucose were estimated to be high in all the varieties in initial stages but a gradual decrease in sucrose and glucose contents was also observed with tuber growth. This may be due to the fact that youngest tubers were having relatively large connection with acid invertase activity. Vacoular acid invertase activity is being dominated by sucrose synthase activity in vegetative growing stages [4,21,22]. However fructose contents were estimated to be lower than glucose and sucrose in early stages. High ratio of glucose to fructose is due to increase in the activity of fructose specific hexokinase, which is higher than glucose specific hexokinase as reported by Davis and Opark [23]. Davis and Viola reported that the decline in activities of sucrose synthase, fructose kinase and glucose-phosphorylation might induce the reduction ratio of glucose to fructose from ca.80:1 to 2:1. Similar results were achieved in our study where values of glucose, sucrose and fructose contents were decreased toward tuber maturation but slight variations in the ratio might be due to genetic variability of the varieties and fluctuation in the enzyme activity (Fig. 2). In the current study, various values of sugars were obtained at varied storage temperatures. These results followed the same pattern of sugar accumulation as reported by Hammond and Zrener [8,9]. In varieties (A, B & D) similar changes of sugar contents and invertase activity was observed. In variety D instead of glucose, fructose values were high which indicate positive correlation of vacuolar acid invertase enzyme activity with glucose, fructose contents. A comparison of type 2 acid invertase activity i.e. 2.5 nm glucose/min/mg with cultivar (C) during storage period was same but a slight increase occur during 10oC while at 4 oC activity was low as compared to type 1. Cultivar (C) showed high sucrose contents and low reducing sugar throughout storage period. Cultivar E was most attention-able. Sucrose and reducing sugars were low along with low invertase activity pattern, similar during curing period and throughout the storage period.
Increase in sucrose contents of Multa might be due to the genetic background of Solanum tuberosum sub sp. Andigena [4]. Its yellowish orange colour was because of high carotenoid contents [24] so it can be referred to as highly resistant potato cultivar tolerant to cold. Variety E was called low sugar accumulating cold tolerant resistant cultivar due to its genetic background, ND 860-2. Thus the data obtained in cultivar E was similar to that observed in studies of cold-tolerant lines, ND 860-2 [10]. Sugar contents for result A, B & D were highly corresponding with the vacuolar acid activity observed in type 1 vacuolar acid activity. With increasing storage incubation period vacuolar acid activity increased. These findings are highly corresponding with the reported study of Endo [4]. Real Time PCR results expression corresponds to similar findings. The transcription level at 20oC was negligible, while at 10oC slight increase was observed, which continued at 4oC during different storage period. Statistical analysis suggested that the conversion of glucose and fructose into sucrose was higher at vegetative growth stage of potato genotypes. Higher genetic advance also revealed that the genotypes may be used to fix increase in sucrose at growing stage of potato [25]. Further findings suggested that potato storage at various temperature conditions resulted in the higher glucose contents at 4°C and 10°C as compared to sucrose and fructose contents [26]. Storage of potato tubers at 20°C found helpful to improve sucrose amount in potato and to develop good quality potato producing genotypes [27]. It was further concluded from results that there is significant change in the contents of fructose, glucose and sucrose at growth stage and under different temperature treatments [28]. Organic acids especially malic acid concentration was high during storage at 4°C as compared to harvesting stage [29]. Citric and oxalic acid concentrations were not affected during storage [30]. A positive correlation of malic acid was found with increased activity of vacuolar acid invertase. Initially at harvesting, malic acid concentration was low as compared to citric and oxalic as shown in Figure 4. The genotypes that showed higher sucrose at different temperatures (Multa) may be used for the development of higher yielding and good quality potato chipping varieties [29-34].
References
- Ali A, Jansky S. Fine screening for resistance to cold-induced sweetening in potato hybrids containing Solanum raphanifolium germplasm. Advances in Agriculture, (2015); 2015: 4.
- Sowokinos J, Belknap W, Vayda M, Park W. Post-harvest regulation of sucrose accumulation in transgenic potatoes: role and properties of potato tuber UDP-glucose pyrophosphorylase. The molecular and cellular biology of the potato, (1994); (Ed. 2): 81-106.
- Chuda Y, Ono H, Yada H, Ohara-Takada A, Matsuura-Endo C, et al. Effects of physiological changes in potato tubers (Solanum tuberosum L.) after low temperature storage on the level of acrylamide formed in potato chips. Bioscience, biotechnology, and biochemistry, (2003); 67(5): 1188-1190.
- Matsuura-Endo C, Kobayashi A, Noda T, Takigawa S, Yamauchi H, et al. Changes in sugar content and activity of vacuolar acid invertase during low-temperature storage of potato tubers from six Japanese cultivars. Journal of plant research, (2004); 117(2): 131-137.
- McKenzie MJ, Sowokinos JR, Shea IM, Gupta SK, Lindlauf RR, et al. Investigations on the role of acid invertase and UDP-glucose pyrophosphorylase in potato clones with varying resistance to cold-induced sweetening. American Journal of Potato Research, (2005); 82(3): 231-239.
- Richardson D, Davies H, Ross H, Mackay GR. Invertase activity and its relation to hexose accumulation in potato tubers. Journal of Experimental Botany, (1990); 41(1): 95-99.
- Zrenner R, Schüler K, Sonnewald U. Soluble acid invertase determines the hexose-to-sucrose ratio in cold-stored potato tubers. Planta, (1996); 198(2): 246-252.
- Hammond JB, Burrell MM, Kruger NJ. Effect of low temperature on the activity of phosphofructokinase from potato tubers. Planta, (1990); 180(4): 613-616.
- Blenkinsop R, Yada R, Marangoni A. Metabolic control of low-temperature sweetening in potato tubers during postharvest storage. Horticultural Reviews, (2004); 30317-354.
- Liu X, Cheng S, Liu J, Ou Y, Song B, et al. The potato protease inhibitor gene, St-Inh, plays roles in the cold-induced sweetening of potato tubers by modulating invertase activity. Postharvest Biology and Technology, (2013); 86265-271.
- Liu X, Lin Y, Liu J, Song B, Ou Y, et al. StInvInh2 as an inhibitor of StvacINV1 regulates the cold‐induced sweetening of potato tubers by specifically capping vacuolar invertase activity. Plant biotechnology journal, (2013); 11(5): 640-647.
- Bradford MM. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical biochemistry, (1976); 72(1-2): 248-254.
- Laude TP, Carena MJ. Diallel analysis among 16 maize populations adapted to the northern US Corn Belt for grain yield and grain quality traits. Euphytica, (2014); 200(1): 29-44.
- Bracho GE, Whitaker JR. Characteristics of the inhibition of potato (Solanum tuberosum) invertase by an endogenous proteinaceous inhibitor in potatoes. Plant physiology, (1990); 92(2): 381-385.
- Steel RG, JH Dickey D Principles and procedures of statistics a biometrical approach. Chapter: Book Name. 1997 of publication; 400-428. WCB/McGraw-Hill.
- Burton GW. Quantitative inheritance in pearl millet (Pennisetum glaucum). Agronomy Journal, (1951); 43(9): 409-417.
- Falconer D, Mackay T. I ntroduction to Q uantitative Genetics. Longman, (1995); 19(8): 1.
- Ballester A-R, Molthoff J, de Vos R, te Lintel Hekkert B, Orzaez D, et al. Biochemical and molecular analysis of pink tomatoes: deregulated expression of the gene encoding transcription factor SlMYB12 leads to pink tomato fruit color. Plant physiology, (2010); 152(1): 71-84.
- Espen L, Morgutti S, Abruzzese A, Negrini N, Rivetta A, et al. Changes in the potato (Solanum tuberosum L.) tuber at the onset of dormancy and during storage at 23 C and 3 CI Biochemical and physiological parameters. Potato research, (1999); 42(1): 189-201.
- Hill L, Reimholz R, Schröder R, Nielsen TH, Stitt M. The onset of sucrose accumulation in cold‐stored potato tubers is caused by an increased rate of sucrose synthesis and coincides with low levels of hexose‐phosphates, an activation of sucrose phosphate synthase and the appearance of a new form of amylase. Plant, Cell & Environment, (1996); 19(11): 1223-1237.
- Davies H, Viola R, Belknap W, Vayda M, Park W. Control of sugar balance in potato tubers. The molecular and cellular biology of the potato, (1994); (Ed. 2): 67-80.
- Pressey R. Potato sucrose synthetase: purification, properties, and changes in activity associated with maturation. Plant physiology, (1969); 44(5): 759-764.
- Davies H, Oparka K. Hexose metabolism in developing tubers of potato (Solanum tuberosum L.) cv Maris Piper. Journal of plant physiology, (1985); 119(4): 311-316.
- Ishii G, Mon M, Ohara A, Umemura V. Food chemical properties of a new potato with orange flesh. SPECIAL PUBLICATION-ROYAL SOCIETY OF CHEMISTRY, (1999); 229357-359.
- Hoehenwarter W, van Dongen JT, Wienkoop S, Steinfath M, Hummel J, et al. A rapid approach for phenotype‐screening and database independent detection of cSNP/protein polymorphism using mass accuracy precursor alignment. Proteomics, (2008); 8(20): 4214-4225.
- Cheng L, Zhang X, Zhao Q, Li H, Wang Y, et al. Comparative proteomic analysis of cold-induced sweetening in potato (Solanum tuberosum L.) tuber. Acta physiologiae plantarum, (2014); 36(5): 1197-1210.
- Xue H-L, Bi Y, Tang Y-M, Zhao Y, Wang Y. Effect of cultivars, Fusarium strains and storage temperature on trichothecenes production in inoculated potato tubers. Food chemistry, (2014); 151236-242.
- Ignat A, Manzocco L, Brunton NP, Nicoli MC, Lyng JG. The effect of pulsed electric field pre-treatments prior to deep-fat frying on quality aspects of potato fries. Innovative Food Science & Emerging Technologies, (2015); 2965-69.
- Aiello D, Abriano S, Cinquerrui A, Firrao G, Polizzi G et al. Ability and synergistic effects of antago-nistic pseudomonas and pantoea spp. To cause vascular discolouration and pith necrosis in tomato plants. Journal of Plant Pathology, (2013); 95(4): 35-S34.
- Carocho M, Barros L, Antonio AL, Barreira JC, Bento A, et al. Analysis of organic acids in electron beam irradiated chestnuts (Castanea sativa Mill.): Effects of radiation dose and storage time. Food and chemical toxicology, (2013); 55348-352.
- Bhaskar PB, Wu L, Busse JS, Whitty BR, Hamernik AJ, et al. Suppression of the vacuolar invertase gene prevents cold-induced sweetening in potato. Plant physiology, (2010); 154(2): 939-948.
- Sowokinos JR. Biochemical and molecular control of cold-induced sweetening in potatoes. American Journal of Potato Research, (2001); 78(3): 221-236.
- Pressey R. Role of invertase in the accumulation of sugars in cold-stored potatoes. American Potato Journal, (1969); 46(8): 291-297.
- Ali A, Muzaffar A, Awan MF, Din S, Nasir IA, et al. Genetically Modified Foods: Engineered tomato with extra advantages. Adv life sci, (2014); 1(3): 139-152.