

Full Length Research Article
Male predominant association with Apolipoprotein B mRNA-editing enzyme, catalytic polypeptide-like 3G variants (rs6001417, rs35228531, rs8177832) predict protection against HIV-1 infection
Qaisar Ali1, Arshad Jamal2, Sajjad Ullah1, Ahmed Bilal Waqar1*
Adv. life sci., vol. 7, no. 2, pp. 91-97, February 2020
*– Corresponding Author: Ahmed Bilal Waqar (Email: drabwaqar@yahoo.com)
Authors' Affiliations
2. College of Science (Biology) University of Hail – Kingdom of Saudi Arabia
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Human immunodeficiency virus (HIV) infection, it is a global health concern mainly lead to acquired immune deficiency syndrome (AIDS). There are numerous limitations of this infection particularly in the form of host factors which may limit and interfere HIV-1 replication. The most notable host factors which hinder HIV-1 DNA propagation is the apolipoprotein B mRNA-editing enzyme, catalytic polypeptide- like 3G (APOBEC3G). Any genetic polymorphism of this substantial host factor may impact the host susceptibility pattern to HIV viral infection in different part of the world. The aim of this study to examine genetic variants (rs6001417, rs35228531, rs8177832) effecting HIV-1 infection.
Method: Three variants of APOBEC3G gene polymorphism were genotyped while using RT-PCR method. Frequency distribution of these genotypes was evaluated in both the HIV-1 and healthy group.
Results: The rs6001417 CG (p = 0.03) and rs35228531 CT (p = 0.01) genotypes were found as protective elements, while rs35228531 TT (p = 0.02) and rs8177832 AA (p = 0.03) genotypes had shown susceptibility against the HIV-1 infection. Our data suggest, rs35228531 CT (p = 0.003) and rs8177832 AA (P = <0.001) genotypes have predominant incidences in HIV-1 male population than healthy control.
Conclusion: We predict rs6001417 CG, rs35228531 CT as protective and rs35228531 TT, rs8177832 AA genotypes as a predisposing tool, against the HIV-1 infection in a section of Pakistani population. In addition, male gender was found predominantly high in both protective genotype rs35228531 CT (p = 0.003) and predisposing genotype rs8177832 AA (p = <0.001). The predominant contribution may help the patient to be predict about the status of HIV infection, however, extra efforts are required to study larger cohort of patients to better elucidate the association.
Keywords: HIV; AIDS; APOBEC3G; Polymorphism
Introduction![]()
HIV-1 infection is a complex and global health dilemma. Due to its fatal nature, approximately 37 million individuals worldwide are infected with HIV, with each passing year 3.1 million new cases emerge [1-3]. Recent reports suggest that HIV infection is highly prevalent in Sub-Saharan countries [4], however currently in Pakistan it is the leading cause of morbidity and mortality as it hits approximately 83,468 individuals. Major causes of HIV transmission are contaminated blood and sexual contacts [5,6].
The human immune system provides resistance against foreign objects through numerous immune strategies, leading to hindrance and elimination of foreign objects. In the process, advance and novel immune mechanisms are evolved [7-11]. Advancements in human genetic studies elucidate that both viral and host genetic factors are critically involved in genetic susceptibility or protection toward the infectious agents [12-14]. Moreover, the individuals with homozygousD32 allelic variant of the CCR5 protein have significant and influential outcome, however it does not exhibit complete resistance to HIV infection. Interestingly, truncated CCR5 is not a known co-receptor for HIV virus, yet it provides protection against HIV infection [15,16]. Also, the ethnic differences have crucial role in host predisposition and/or protection against the disease [17-19]. Subsequently, relevance of these genetic and ethnic differences to a particular disease has started to fascinate the researchers worldwide and thus manifesting the better understanding of their potential role in HIV infection or disease progression.
Mounting body of evidence suggests that multiple host factors affect pathogenesis of HIV-1/AIDS. Likely factors include; Apolipoprotein B mRNA editing enzyme catalytic polypeptide-like 3G (APOBEC3G), Chemokine Receptor 5 (CCR-5), Dendritic Cell-Specific Intercellular adhesion molecule-3-Grabbing Non-integrin (DC-SIGN), Tripartite motif 5a (TRIM5a), Tetherin, and SAM-domain HD-domain containing protein (SAMHD1) [5,20-23]. The possible defense mechanisms exhibited by these elements are antagonized by accessory viral proteins [24,25].
The association of gender with spontaneous clearance of HCV has already been reported showing link between female gender and rs12979860 CT genotype and facilitation in the spontaneous clearance of hepatitis C virus potentially [26]. It has been also reported that TNFAIP3 rs2230926G may be preliminary predictor of systemic lupus erythematous onset in males [27].
APOBEC3G (antiviral Apolipoprotein B mRNA-editing enzyme, catalytic polypeptide-like 3G) located on chromosome #22q13 [28] is a host intrinsic element, which combats HIV-1 infection [29]. Virion infectivity factor (vif), a structural part of HIV-1 is able to counteract APOBEC3G antiviral activity by targeting it via proteasomes [30]. The vif protein derived from different subtypes, A, B, CRF01_AE, and CRF02_AG, portray comparable anti APOBEC3G activity. On contrary subtype C vif protein depicts maximal anti APOBEC3G activity [31]. The APOBEC3G protein demonstrates its antiviral action by incorporating itself into newly synthesized viral particles, in the absence of the vif, triggering deamination of cytosine (C) to uracil (U) in newly synthesized mutant viral particles. APOBEC3G polymorphisms, such as (H186R) rs8177832 are associated with pathogenesis in different ethnic groups and various HIV-1 subtypes [32,33], however this association is not reported in other populations [33]. The previous studies did not take concern with circulating recombinant forms of HIV-1, nor examine the effect of APOBEC3G polymorphisms in Pakistani ethnic groups.
The emphasis of present study is to understand and evaluate the role of rs6001417, rs35228531, rs8177832 variants of APOBEC3G polymorphism in HIV-1 infection with circulating recombinant in Pakistani population.
Methods![]()
This study included a total of 206 individuals involving 100 HIV-1 seropositive and 106 HIV-1 sero-negative individuals. Both groups were obtained in the Imperial Diagnostic & Research Center-Imperial College of Business Studies Lahore, Pakistan. This study was approved by the Institutional Ethical Review Board (IERB), Faculty of Health and Allied Sciences (FHAS), Imperial College of Business Studies (ICBS), Lahore. After obtaining the written consent, whole blood was collected from each participant in EDTA container.
HIV-1 Detection
Plasma samples obtained by centrifugation at 3500 rpm for 5 minutes and were tested for HIV–1 infection, using Alere determine HIV-1 or 2 tests (Product code: 7D2346 CE Chicago, Illinois, United States) as per previously reported protocol [34].
Genotypic determination
The extraction of genomic DNA was performed from whole blood leukocytes, using a non-enzymatic salting-out method [35]. APO-BEC-3G (rs6001417, rs35228531, rs8177832) loci were determined using Fast Real-Time PCR (RT-PCR) Systems (Applied Bio-system Step One TM).
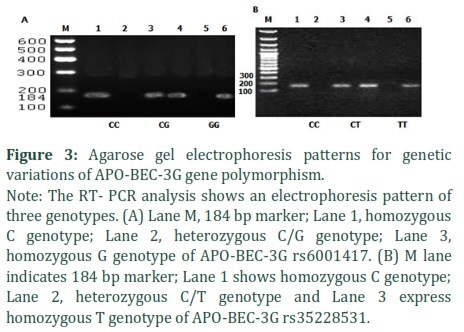
The 184bp DNA fragment of three variants APO-BEC-3G (rs6001417, rs35228531, rs8177832) was amplified by using (SYBR GreenER qPCR super MIX Universal kit Invitrogen USA). The primer sequences used are given in Table 1. The total volume of PCR mixture was 25ul, containing DNA template (1 ul), each primer (1 ul) and 10 ul commercial super Mix containing Taq-polymerase, MgCl2, dNTPs, Syber green fluorescent dye and PCR buffer. Thermocycling was performed as; initial denaturation at 95°C for 5 minutes, followed by 35 cycles of denaturation at 95°C for 30s, annealing at 55°C for 30s and extension at 68°C for 30s.The final extension was performed at 72°C for 07 minutes.
The amplified products were run on 2% agarose gel along with 50 bp ladder (Fermentas USA) and visualized under Wealtec Gel doc system (Fermentas USA).
Statistical Analysis:
The data was shown as Mean ± SD. The frequencies of different genotypes were calculated by direct counting. The Variables in the category were compared in different groups by using the Chi – square test. All statistical analyses were done by using SPSS 22. The independent t –test was applied for the demonstration of quantitative variation in both the patient and control groups. P-value < 0.05 was considered statistically significant.
Results![]()
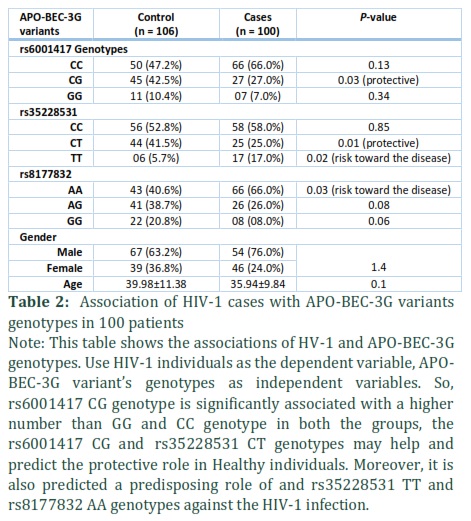
Generally, in 2018, a total of 129,079 new A total of 206 individuals were recruited in current study. The population investigated consisted of 100 HIV-1 infected patients and 106 healthy controls. Age and gender wise distribution of the positive cases and healthy groups are shown in Table 2. The mean ages were 39.98±11.38 and 35.94±9.84 in the control and HIV-1 groups respectively. Genotype frequencies of all the three APOBEC3G loci (rs6001417, rs35228531, rs8177832) for both groups are shown (Table 2).
Association between APOBEC3G rs6001417 variant and HIV-1 status
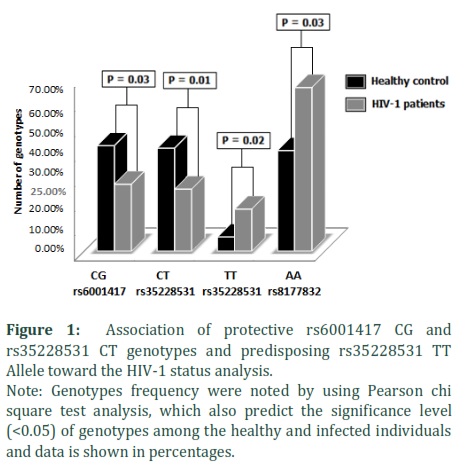
We found that the frequencies of APOBEC3G (rs6001417) CC, CG, and GG genotypes were 66.0%, 27.0%, 7.0% in HIV-1 and 47.2%, 42.5% and 10.4% in the control group (Table 2). Individuals reported positive for HIV-1 infection had lower frequencies of the APOBEC3G (rs6001417) CG, GG genotypes, whereas, CC genotype was increased in HIV-1 patients compared to controls. Thus, higher prevalence of CG genotype in the control group might indicate its possible protective role against HIV-1 infection (45 (42.5%) vs 27 (27.0%), p = 0.03) (Figure 1).
Association between APOBEC3G rs35228531 variant and HIV-1 status
APOBEG3G (rs35228531) CC, CT and TT genotypes were found to be 52.8%, 41.5%, 5.7% and 58.0%, 25.0%, 17.0% in control and HIV-1 positive individuals, respectively (Table 2). The levels of CC genotype were comparable in both control and infected group (p = 0.85), however, CT genotype was significantly higher in control compared to the infected group (41.5% vs 25.0%, p = 0.01).
Furthermore, our data suggest that TT genotype frequency was significantly increased in HIV-1 positive cases compared to control group (17.0% vs 5.7%, p = 0.02). (Figure 1).
Association between APOBEC3G rs8177832 variant and HIV-1 status
APOBEG3G (rs8177832) AA, AG and GG genotypes were found as 40.6%, 38.7%. 20.8% and 66.0%, 26.0%, 08.0% in control and HIV-1 positive individuals, respectively (Table 2). The levels of AG and GG genotypes were comparable in both control and infected groups (p = 0.08 and p = 0.06, respectively), however, AA genotype was significantly lower in control compared to the infected group (40.6% vs 66.0%, p = 0.03) (Figure 1).
Furthermore, we performed correlation analysis in between all the genotypes and the three studied variants of APOBEC3G in both groups but did not find any significant correlation (Table 2).
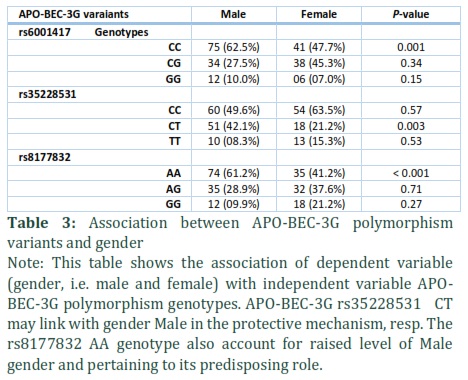
Analysis of gender with Protective (rs6001417) CG and (rs35228531) CT genotype
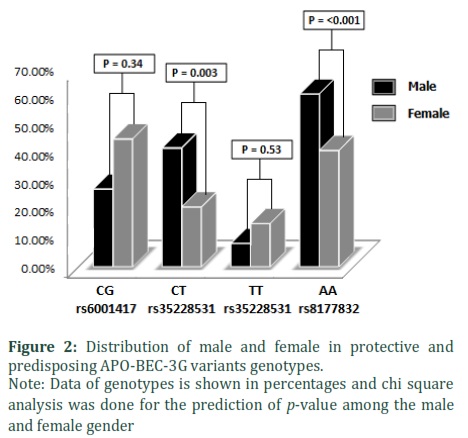
The frequency of protective genotypes of the above-mentioned variants are shown in Table 3. The frequency of rs6001417 CG genotype was slightly higher in the males but did not show statically significance (27.5% vs 45.3%), p = 0.34) (Figure 2). However, rs35228531 CT genotype prevalence was significantly higher in male group, comparatively 42.1%) vs 21.2%; p = 0.003 (Figure 2).
Analysis of gender with Predisposing (rs35228531) TT and (rs8177832) AA genotypes
We found that the distribution of TT genotype was similar in male and female groups [male (08.3%) vs female (15.3%); p = 0.53] however, rs8177832 AA genotype levels were higher in male than female [male (61.2%) vs female (41.2%); p = < 0.001] (Figure 2).
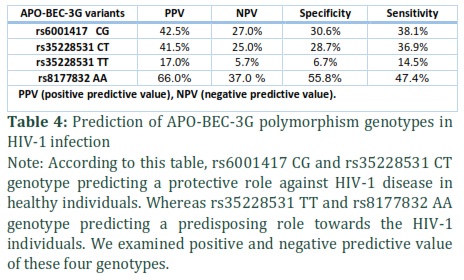
Predictive value of protecting and predispose features associated with APOBEG3G variants genotypes
As we have mentioned in our earlier results sections that the protective features are linked with rs6001417 CG, rs35228531 CT, and predisposing with rs35228531 TT, rs8177832 AA genotype against HIV-1 infection. In addition, we examined the predictive value of these four genotypes as well by doing statistical analysis, using SPSS version 22.0. Both the rs6001417 CG and rs35228531 CT genotypes were appeared as protective genotypes, we also explored positive predictive value (PPV) which was 42.5% and 41.5%, respectively (Table 4), which predicts the chance in percentage to reduce the risk of being infected with HIV-1 virus. Moreover, the rs35228531 TT and rs8177832 AA genotype had shown susceptibility towards the onset of HIV-1 infection. The PPV calculated was 17.0%, 66.0%, which predicts the role of these genotypes regarding the increased risk of being infected with HIV-1 virus (Table 4).
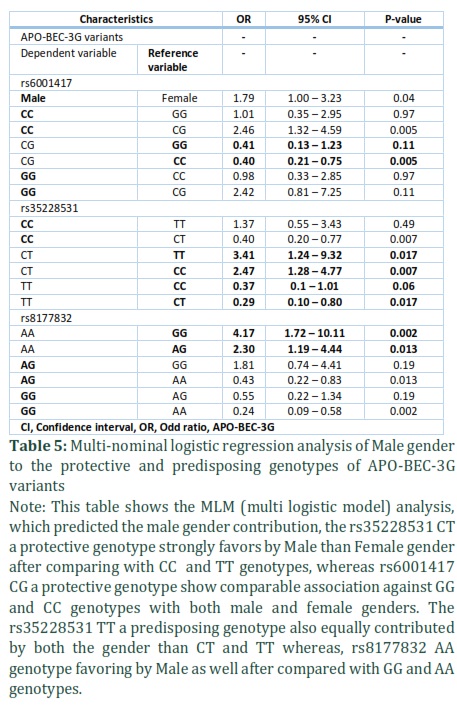
Multivariate logistic regression analysis of Male gender with protective and predisposing genotypes
We employed multi-nominal logistic regression to interpret the association between APOBEG3G variants (protective and predisposing) genotypes and gender in the healthy control and HIV-1 group and analyzed the following factors: APOBEG3G rs35228531 CT (CC and TT were taken as reference genotypes) and gender (male vs female). These factors independently and significantly contributed to protection against HIV-1 infection, rs35228531 CT vs CC (reference genotype) (OR, 2.47; CI, 1.28 – 04.77; p = 0.007) and CT vs TT (reference genotype) (OR, 3.41; CI, 1.24 – 9.32; P = 0.017). The CT (protective genotype) (51 (42.1%) vs 18 (21.2%); (p = < 0.001) was more prevalent in male than female group (Table 5) Factor involving in the predisposition of the HIV-1 infection in rs35228531 TT (predisposed genotype) had a comparable association with male and female gender in the disease progression (effecting by CC and CT genotypes; OR, 0.37; CI, 0.1 – 1.01; P = 0.06 and OR, 0.29; CI, 0.10 – 0.80; p = 0.017, respectively) (Table 5).
Moreover, our analysis also revealed the significant effect of male gender on rs8177832 AA a predisposed genotype towards HIV-1 infection (effecting by AG and GG genotypes; OR, 4.17; CI, 1.72 – 10.11; p = 0.002 and OR, 2.30; CI, 1.19 – 4.44; p = 0.013, respectively) (Table 5). HIV-1 studied individuals with rs6001417 CG (protective genotype) almost didn’t show any distinct contribution for both male and female ((effecting by GG and CC genotypes; OR, 0.41; CI, 0.13 – 1.23; P = 0.11 and OR, 0.40; CI, 0.21 – 0.75; P = 0.005, respectively) (Table 5).
Figures & Tables

Discussion![]()
Continual exposure to HIV infection may not certainly result in AIDS occurrence [36]. Multiple genetic and immune factors help in HIV acquirement, pathogenesis and AIDS progression at various stages of HIV life-cycle. So, HIV infection activates multiple intrinsic host factors that confer resistance to HIV pathogenesis. One of the important factors that substantiate intrinsic inhibition to HIV infection is APOBEC3G genetic host factor [24,37]. APOBEC3G single nucleotide polymorphisms (SNPs) are of particular importance. Its numerous SNPs have been studied in American [38] and European [39] populations in relation to its influence on AIDS development and progression. To our knowledge, the role of APOBEC3G in relation to HIV-1 infection has not been reported before in Pakistani population. Therefore, in current study we have investigated the frequency distribution of the three variants of APOBEC3G gene polymorphism in residents of Pakistan [40] and association of these variants with HIV-1 pathogenesis.
In current study, we have reported the possible biological role of APOBEC3G gene variants (rs35228531, rs35228531 and rs6001417) in promoting and/or repressing the HIV-1 infection between male and female in Pakistani population. As far as we know, this is the first study examining the role of APOBEC3G gene variants on HIV-1 infection in a Pakistani population in both gender, where we found male a possible predominant . In short, male gender was found predominatly high in both protective gentotype rs35228531 CT (p = 0.003) and predisposing genotype rs8177832 AA (p = <0.001). This predomiant contribution may help the patient to be predict about the status of HIV infection. However, the major limitation, is that the current study was carried out on a limited sample size. The findings, based on such a sample size / population fraction are not enough to infer to the whole population. In order to have a better understanding of the above-mentioned association further investigations are needed for a substantial part of the population.
Authors' Contribution
The conduction and designing of this study was done by Qaisar Ali and Arshad Jamal and lab support provided by Sajjad Ullah whereas, Ahmed Bilal Waqar supervised this research.
The authors declare that there is no conflict of interest regarding the publication of this paper.
References![]()
- Barré-Sinoussi F, Chermann J-C, Rey F, Nugeyre MT, Chamaret S, et al. Isolation of a T-lymphotropic retrovirus from a patient at risk for acquired immune deficiency syndrome (AIDS). Science, (1983); 220(4599): 868-871.
- Tang JW, Chan PK. The basics of HIV medicine. SS Lee, JCY, (2007).
- Murray CJ, Ortblad KF, Guinovart C, Lim SS, Wolock TM, et al. Global, regional, and national incidence and mortality for HIV, tuberculosis, and malaria during 1990–2013: a systematic analysis for the Global Burden of Disease Study 2013. The Lancet, (2014); 384(9947): 1005-1070.
- Kharsany AB, Karim QA. HIV infection and AIDS in Sub-Saharan Africa: current status, challenges and opportunities. The open AIDS journal, (2016); 1034.
- Imran M, Manzoor S, Saalim M, Resham S, Ashraf J, et al. HIV‐1 and hijacking of the host immune system: the current scenario. Apmis, (2016); 124(10): 817-831.
- Oberle CS, Joos B, Rusert P, Campbell NK, Beauparlant D, et al. Tracing HIV-1 transmission: envelope traits of HIV-1 transmitter and recipient pairs. Retrovirology, (2016); 13(1): 62.
- An P, Bleiber G, Duggal P, Nelson G, May M, et al. APOBEC3G genetic variants and their influence on the progression to AIDS. Journal of virology, (2004); 78(20): 11070-11076.
- An P, Penugonda S, Thorball CW, Bartha I, Goedert JJ, et al. Role of APOBEC3F gene variation in HIV-1 disease progression and pneumocystis pneumonia. PLoS genetics, (2016); 12(3): e1005921.
- Dunn GP, Bruce AT, Ikeda H, Old LJ, Schreiber RD. Cancer immunoediting: from immunosurveillance to tumor escape. Nature immunology, (2002); 3(11): 991.
- van Kooyk Y, Appelmelk B, Geijtenbeek TB. A fatal attraction: Mycobacterium tuberculosis and HIV-1 target DC-SIGN to escape immune surveillance. Trends in molecular medicine, (2003); 9(4): 153-159.
- D'Urbano V, De EC, Re M. Host restriction factors and Human immunodeficiency Virus (HIV-1): a dynamic interplay involving all phases of the viral life cycle. Current HIV research, (2018).
- Bataller R, North KE, Brenner DA. Genetic polymorphisms and the progression of liver fibrosis: a critical appraisal. Hepatology, (2003); 37(3): 493-503.
- Janssen R, Bont L, Siezen CL, Hodemaekers HM, Ermers MJ, et al. Genetic susceptibility to respiratory syncytial virus bronchiolitis is predominantly associated with innate immune genes. Journal of Infectious Diseases, (2007); 196(6): 826-834.
- Powell EE, Edwards‐Smith CJ, Hay JL, Clouston AD, Crawford DH, et al. Host genetic factors influence disease progression in chronic hepatitis C. Hepatology, (2000); 31(4): 828-833.
- Piacentini L, Biasin M, Fenizia C, Clerici M. Genetic correlates of protection against HIV infection: the ally within. J Intern Med, (2009); 265(1): 110-124.
- A Estrada-Aguirre J, G Cazarez-Salazar S, A Ochoa-Ramirez L, de J Acosta-Cota S, Zamora-Gómez R, et al. Protective effect of CCR5 Delta-32 allele against HIV-1 in Mexican women. Current HIV research, (2013); 11(6): 506-510.
- Kenny-Walsh E. The natural history of hepatitis C virus infection. Clinics in liver disease, (2001); 5(4): 969-977.
- Dolo A, Modiano D, Maiga B, Daou M, Dolo G, et al. Difference in susceptibility to malaria between two sympatric ethnic groups in Mali. The American journal of tropical medicine and hygiene, (2005); 72(3): 243-248.
- Perera FP. Molecular epidemiology: insights into cancer susceptibility, risk assessment, and prevention. JNCI: Journal of the National Cancer Institute, (1996); 88(8): 496-509.
- Duggal NK, Emerman M. Evolutionary conflicts between viruses and restriction factors shape immunity. Nature Reviews Immunology, (2012); 12(10): 687.
- Kagoné TS, Bisseye C, Méda N, Testa J, Pietra V, et al. A variant of DC-SIGN gene promoter associated with resistance to HIV-1 in serodiscordant couples in Burkina Faso. Asian Pacific journal of tropical medicine, (2014); 7S93-S96.
- Harris RS, Liddament MT. Retroviral restriction by APOBEC proteins. Nature Reviews Immunology, (2004); 4(11): 868.
- Merindol N, Berthoux L. Restriction factors in HIV-1 disease progression. Current HIV research, (2015); 13(6): 448-461.
- Imran M, Manzoor S, Saalim M, Resham S, Ashraf J, et al. HIV-1 and hijacking of the host immune system: the current scenario. APMIS, (2016); 124(10): 817-831.
- Ayinde D, Casartelli N, Schwartz O. Restricting HIV the SAMHD1 way: through nucleotide starvation. Nature Reviews Microbiology, (2012); 10(10): 675.
- Ali Q, Jamal A, Imran M, Ullah S, Kalam I, et al. Correlation of IL28B rs12979860 genotype and gender with spontaneous clearance of HCV infection: a Pakistani cross-section study. Personalized medicine, (2018); 15(6).
- Kadota K, Mori M, Yanagimachi M, Miyamae T, Hara T, et al. Analysis of gender differences in genetic risk: association of TNFAIP3 polymorphism with male childhood-onset systemic lupus erythematosus in the Japanese population. PLoS One, (2013); 8(8): e72551.
- An P, Johnson R, Phair J, Kirk GD, Yu X-F, et al. APOBEC3B deletion and risk of HIV-1 acquisition. The Journal of infectious diseases, (2009); 200(7): 1054-1058.
- Svarovskaia ES, Xu H, Mbisa JL, Barr R, Gorelick RJ, et al. Human apolipoprotein B mRNA-editing enzyme-catalytic polypeptide-like 3G (APOBEC3G) is incorporated into HIV-1 virions through interactions with viral and nonviral RNAs. Journal of Biological Chemistry, (2004); 279(34): 35822-35828.
- Feng Y, Love RP, Chelico L. HIV-1 viral infectivity factor (Vif) alters processive single-stranded DNA scanning of the retroviral restriction factor APOBEC3G. Journal of Biological Chemistry, (2013); 288(9): 6083-6094.
- Iwabu Y, Kinomoto M, Tatsumi M, Fujita H, Shimura M, et al. Differential anti-APOBEC3G activity of HIV-1 Vif proteins derived from different subtypes. Journal of Biological Chemistry, (2010); 285(46): 35350-35358.
- Bishop KN, Holmes RK, Malim MH. Antiviral potency of APOBEC proteins does not correlate with cytidine deamination. Journal of virology, (2006); 80(17): 8450-8458.
- Sheehy AM, Erthal J. APOBEC3 versus retroviruses, immunity versus invasion: clash of the titans. Molecular biology international, (2012); 2012.
- Eller L, Manak M, Shutt A, Malia J, Trichavaroj R, et al. Performance of the Determine HIV 1/2 Ag/Ab Combo Rapid Test on Serial Samples from an Acute Infection Study (RV217) in East Africa and Thailand; 2013. MARY ANN LIEBERT, INC 140 HUGUENOT STREET, 3RD FL, NEW ROCHELLE, NY 10801 USA. pp. A73-A74.
- Suguna S, Nandal D, Kamble S, Bharatha A, Kunkulol R. Genomic DNA isolation from human whole blood samples by non enzymatic salting out method. Int J pharm pharm sci, (2014); 6198-199.
- Dean M, Carrington M, Winkler C, Huttley GA, Smith MW, et al. Genetic restriction of HIV-1 infection and progression to AIDS by a deletion allele of the CKR5 structural gene. Science, (1996); 273(5283): 1856-1862.
- McNab F, Mayer-Barber K, Sher A, Wack A, O'garra A. Type I interferons in infectious disease. Nature Reviews Immunology, (2015); 15(2): 87.
- An P, Bleiber G, Duggal P, Nelson G, May M, et al. APOBEC3G genetic variants and their influence on the progression to AIDS. J Virol, (2004); 78(20): 11070-11076.
- Do H, Vasilescu A, Diop G, Hirtzig T, Heath SC, et al. Exhaustive genotyping of the CEM15 (APOBEC3G) gene and absence of association with AIDS progression in a French cohort. J Infect Dis, (2005); 191(2): 159-163.
- Nadembega W, Giannella S, Simporé J, Ceccherini‐Silberstein F, Pietra V, et al. Characterization of drug‐resistance mutations in HIV‐1 isolates from non‐HAART and HAART treated patients in Burkina Faso. Journal of medical virology, (2006); 78(11): 1385-1391.
- Saeed MA, Ahmed N, Ali Q. Frequency of Autoantibodies to Thyroid Peroxidase (TPO) in Patients with Type 1 Diabetes Mellitus. (2018).
- Compaore TR, Soubeiga ST, Ouattara AK, Obiri-Yeboah D, Tchelougou D, et al. APOBEC3G Variants and Protection against HIV-1 Infection in Burkina Faso. PloS one, (2016); 11(1).
- Do H, Vasilescu A, Diop G, Hirtzig T, Heath SC, et al. Exhaustive genotyping of the CEM15 (APOBEC3G) gene and absence of association with AIDS progression in a French cohort. Journal of Infectious Diseases, (2005); 191(2): 159-163.
- Iqbal K, Imran M, Ullah S, Jamal M, Waheed Y, et al. Correlation of apolipoprotein B mRNA-editing enzyme, catalytic polypeptide-like 3G genetic variant rs8177832 with HIV-1 predisposition in Pakistani population. Current HIV research, (2018); 16(4): 297-301.
- De Maio FA, Rocco CA, Aulicino PC, Bologna R, Mangano A, et al. Effect of HIV-1 Vif variability on progression to pediatric AIDS and its association with APOBEC3G and CUL5 polymorphisms. Infection, Genetics and Evolution, (2011); 11(6): 1256-1262.
- Ali Q, Kalam I, Ullah S, Jamal A, Imran M, et al. Predictive value of IL-28B rs12979860 variants for peg-IFN, sofosbuvir plus ribavirin treatment of HCV infection in Pakistani population. Personalized medicine, (2018); 15(6).
- Kalam I, Ullah S, Ali Q, Jamal A, Waqar AB. Impact of IL28B gene variants (rs12979860) in peg-IFN therapy against Chronic Hepatitis B Pakistani patients. Advancements in Life Sciences, (2018); 6(1): 11-18.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0
![]()