

Full Length Research Article
Biocontrol of MRSA and E. coli using bacteriophages from cow manure
Nimerta Kumari, Bushra Bano Patoli*, Atif A Patoli, Shagufta Jabeen
Adv. life sci., vol. 7, no. 4, pp. 264-269, August 2020
*– Corresponding Author: Bushra Bano Patoli (Email: bushrapatoli@gmail.com)
Authors' Affiliations
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Bacteriophages can be used as biocontrol agents to eliminate the undesirable and targeted bacteria in human beings, environment and industries. We aimed to isolate phages from cow manure and investigate their antibacterial and anti-biofilm formation effect against MRSA and E. coli species.
Method: E. coli and S. aureus isolates of clinical origin were sub-cultured on MacConkey’s and Mannitol salt agar media, respectively. To confirm the Methicillin resistance in S. aureus, mecA gene was detected by the PCR using gene specific forward and reverse primers. The bacteriophages were isolated using enrichment procedure from cow manure. Their antibacterial and anti-biofilm formation activity was determined through the inhibition of bacterial growth and decrease in the absorbance of Crystal Violet through biofilm inhibition assay, respectively.
Results: The isolated phages showed clear plaque formation against MRSA and E. coli species. The time-dependent lytic assay showed a sharp decline in OD600 of MRSA and E. coli after one hour incubation with the phages. Biofilm inhibition was indicated by a decrease in the absorbance (OD595) of CV in a microtiter well plate
Conclusion: This is perhaps the first study to isolate phages from cow manure at local region and demonstrate their lytic efficiency against MRSA and E. coli, with the ultimate aim to use them as a promising biocontrol agent against antibiotic resistant pathogens.
Keywords: Phages; MRSA; Plaque formation; Biofilm inhibition; E. coli; Biocontrol
Introduction![]()
Bacteriophages simply called as “phages” are viruses that infect and multiply within the host bacterial cell. The discovery of viruses in the early 20th century [1] has led us to some interesting findings, that is, they are ubiquitous in nature, capable of infecting the cells in all three domains of life including Archaea, Bacteria and Eukarya. The unique structure of viruses and their ability to infect bacterial cells unearthed various fundamental biological processes. The genetic makeup of viruses was harnessed as a molecular tool and led to the development of recent field of molecular biology. Based on the study of coliphages (T series, Lambda, Mu, PhiX174) and others, the molecular mechanisms of virus-host interactions, genetic recombination (transduction), genetic engineering and CRISPR were discovered [2,3]. Various genetic tools (recombinase/integrase and restriction enzymes) were discovered from the earliest studied phages like lambda phage, a temperate phage capable of both lytic and lysogenic cycle in E. coli [4]. The specificity to the host cell and the lytic capability of bacteriophages makes them an ideal tool as an antimicrobial agent [5] to combat bacterial infections and contamination on the non-living surfaces. Besides, the lytic property of bacteriophages is exploited to treat bacterial infections and the phage therapy is now considered again an alternate to the antibiotics due to the alarming increase in the antibiotic resistant bacteria. The natural occurrence of bacteriophages in the environment makes them a suitable alternate to the chemical to control the growth of potential pathogens and the bacterial load on the surfaces, wastewater and the sludge volume without posing a risk to the other flora of natural ecology. Several studies and clinical trials are now conducted to suggest phages a favorable substitute to the antibiotics [6]. Previous studies suggests that the phages can cure ~ 90% of the chronic suppurative bacterial infections, peritonitis, osteomyelitis, dysentery in humans caused by antibiotic-resistant bacteria such as, Staphylococcus aureus, Pseudomonas aeruginosa, Klebsiella pneumoniae and E. coli [5].
The wide spread prevalence of Methicillin-Resistant Staphylococcus aureus (MRSA) is major threat to the health care, causing ~12,000 deaths annually in the US only. Animal to human transmission of MRSA is also reported [7,8]. There are several reports of multi drug resistant E. coli causing multiple infections in human being [9]. These bacterial pathogen are also well documented to produce biofilms, an important survival strategy, on surfaces and medical devices [10,11]. The dense microbial growth in biofilms allows bacteria adhere to the surfaces and get protected in the self-secreted glycocalyx from the antimicrobial effects. It is reported that surfaces exposed to phages prevented the biofilm formation by bacteria [12]. The phage associated depolymerase enzymes helps them to break the exo-polysaccharide and interrupt the biofilm matrix and ultimately results in the lysis of cell [13].
In the present study we isolated the phages from livestock manure using S. aureus and E. coli as a specific host to the phages from the environment. To the best of our knowledge, this is the first study to isolate and test the antibacterial activity of phages from the cow manure at local region against the MRSA and E. coli. The isolated phages have demonstrated themselves a promising anti-biofilm agent against these pathogenic organisms.
Methods![]()
The present study was conducted in HEC funded Molecular Microbiology and Genetics Laboratory at the Institute of Microbiology, University of Sindh, Jamshoro. The study does not involve any human participant so it does not require any ethical approval.
Bacterial culture and growth condition
Non-clinical E. coli and S. aureus strains were isolated from water and the door knobs at the Institute of Microbiology, University of Sindh, and Jamshoro. The clinical strains, MRSA and E. coli, were obtained from the diagnostic and research laboratory of Liaquat University Medical Hospital, Jamshoro, Sindh. The growth of S. aureus and E. coli was maintained by using Mannitol Salt Agar and MacConkey’s agar media (Oxoid), respectively. The identification of the isolates was performed using their cultural and biochemical characteristics according to the standard methods. To confirm the S. aureus species, coagulase test was performed. To detect the presence of mecA gene in MRSA strains Polymerase Chain Reaction (PCR) was performed by using the specific primers: mecA1 381 [5’] GTA GAA ATG ACT GAA CGT CCG ATA A- 342 and mecA2 603 [3’] CCA ATT CCA CAT TGT TTC GGT CTA A-627. S. aureus, ATCC® 33591M (MediMark® Europe) strain was used as a positive control strain for the detection of mecA gene. PCR amplified product was observed at ~310 bp regions using agarose gel electrophoresis in the gel doc system (BioBasic). For long term storage, the strains were stock preserved at -5°C using 15% glycerol in Nutrient Broth (NB) (Oxoid UK CM0003). For all the tests the bacterial strains were cultured using NB medium at 37°C in shaking incubator or on Nutrient agar (NA Oxoid UK) plates (Lab-Lemco’ powder 0.1%, Yeast extract 0.2%, Peptone 0.5%, Sodium chloride 0.5%, Agar 1.5% pH 7.4 ± 0.2).
Isolation and enrichment of phages from environment
The phages were isolated from four different fresh cow manure samples in September 2019 according to the method described earlier [14] with some modifications. To recover the phages, 1 gm of fresh cow manure was diluted ten times (1:10) in sterile distilled water, vortexed to mix and centrifuged at 5000 rpm for 10 minutes at room temperature. The clear supernatant was collected in a fresh Eppendorf tube and mixed with chloroform at 1:10 dilution. After the incubation of 30 minutes, the samples were filtered using 0.45 µm pore size filter by the Millipore filter assembly.
To enrich the prospective phages, two hours liquid cultures of test strains (S. aureus and E. coli) at an OD600 < 1 were supplemented with 2 mM final concentration of CaCl2 and incubated for 10 minutes to facilitate the attachment of the isolated phages with the host strains [15]. One milliliter of the obtained sample filtrate (presumably containing the phages) was then mixed with CaCl2 treated culture and kept for two hours in the shaking incubator at 37°C to achieve the phage propagation and cell lysis. The phage lysate was then purified from the cell debris by adding 0.5 ml of chloroform to kill any intact cells. The lysate was then centrifuged at 5000 rpm for ten minutes to obtain the clear supernatant which was filtered again using 0.45µm pore size filter. The enriched phage lysates were then immediately used or kept in refrigerator at 4°C for further tests.
Lysis of S. aureus and E. coli by lytic phages
The obtained phage lysate was used as the source for the screening of phages capable of lysing S. aureus by the method described previously with some modifications [14]. For this a fresh liquid culture of the test strains (clinical strains MRSA and E. coli) were prepared. To grow the MRSA strain, a single colony from MSA plate was inoculated in nutrient broth medium containing 5 % NaCl and 0.5 % glucose and kept in the shaking incubator at 37°C for two hours or more until the OD600 of 0.5 is achieved. The liquid cultures were then used to form the bacterial lawn with the help of sterile cotton swab on nutrient agar plates. A fixed amount (10 µl) of the two different phage lysates was spot inoculated over the bacterial lawn and allowed to settle down to prevent the dripping of liquid from the nutrient agar plates. The plates were incubated at 37°C for 24 hours. The same procedure was adopted to test the E. coli strain with the exception of adding extra 5 % NaCl in the nutrient broth medium. To ensure the maximum purity of the phage lysate, the individual plaques were repeatedly collected and concentrated by growing with the test bacterial strains (Figure 1).
Qualitative assay of phages by PFU/ml
Plaque formation assay was performed to enumerate the phages as described earlier [16]. For this overnight cultures of E. coli and MRSA were diluted (1:100) in nutrient broth, with the addition of 5% NaCl for MRSA culture. The cultures were incubated at 37°C with shaking for one hour. The cells were treated with 2 mM concentration of CaCl2 and incubated for further 15 minutes. The treated cell culture (100 µl) was mixed with the phage lysate (100 µl) for few minutes and dispensed into 3 ml of soft top-agar (melted nutrient broth with 0.7% agar) in a sterile test tube. The mixture was poured onto solid nutrient agar plates immediately and incubated for 16 hours at 37°C. After the incubation period the Plaque Forming Units per milliliter (PFU/ml) were counted for the qualitative analysis of the virus (Figure 2 a & b). The culture with no phage lysate served as a control (Figure 2 c & d).
Time dependent killing of bacteria by phages
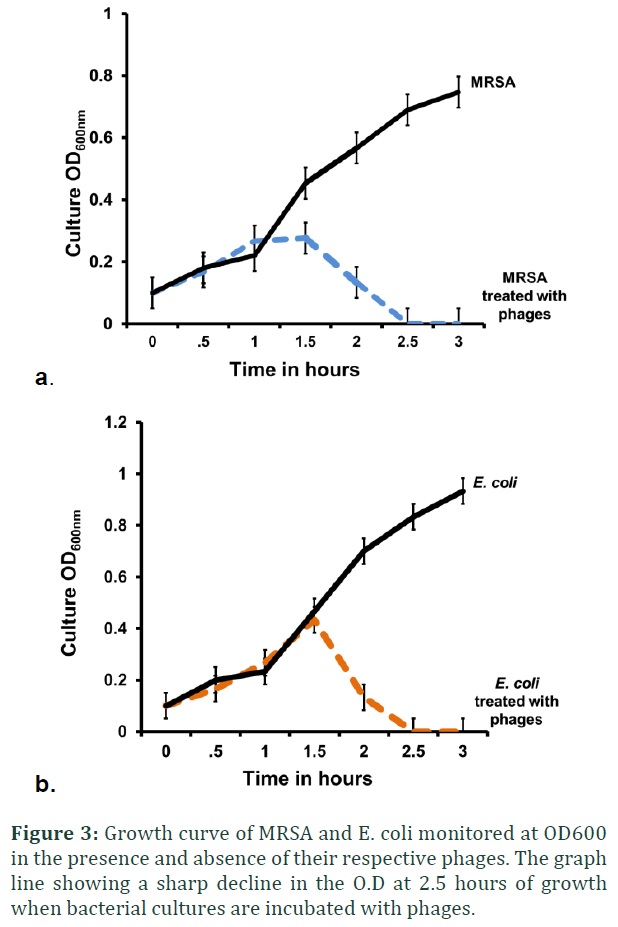
The overnight broth culture of E. coli and MRSA were diluted 1:100 in 2x nutrient broth (equals to about 107CFU/ml). The filtered solution of CaCl2 is added in bacterial culture to make final concentration of 2mM. One milliliter of bacterial culture was mixed with 100µl respective phage lysate and kept in gentle shaking at 37°C. No phage lysate is added in control culture flasks. The optical density (OD600) of each culture was monitored every 30 minutes to determine the lysis efficacy of phages for three hours (Figure 3).
Inhibition of bacterial biofilm formation by phages
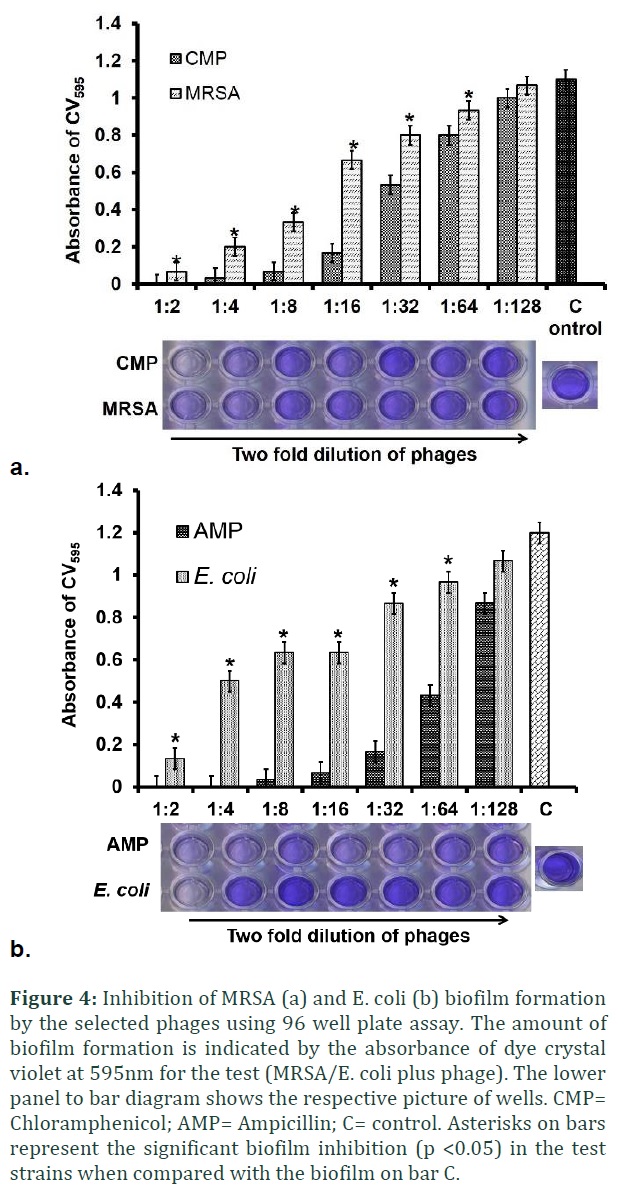
Inhibition of biofilm formation was determined for those phages which showed no clear lysis on spot assay plate (Figure 1). Briefly, the overnight broth culture of E. coli and MRSA were diluted 1:100 in nutrient broth medium containing 0.5% glucose and 2 mM CaCl2 final concentration. In a flat-bottomed polystyrene 96-well microtiter plates (Corning NY), a total of seven two-fold dilutions (1:2, 1:4, 1:8, 1:16, 1:32, 1:64 and 1:128) of the phage lysate were prepared in NB medium containing 0.5% glucose and dispensed as 100µl volume in each well. For positive control, half MIC (1/2) of Chloramphenicol (5 µg/ml) was used for S. aureus and half MIC of Ampicillin (100µg/ml) was used for E. coli in the control well to inhibit the biofilms formation [17]. Two-fold dilutions of the antibiotics were made as described for phage lysate.
A fixed amount of the broth culture (100 µl) of each bacterial strain (MRSA and E. coli) was added to their respective wells. The microtiter plates were incubated for 24 h at 37°C. After incubation, the planktonic cells were gently discarded; wells were washed three times with phosphate buffer saline (PBS). The plates were blotted on tissue paper to dry. The cells adhered to the wells were stained with 200µl of 0.1% Crystal Violet (CV) dye for 15 min. The plates were washed four times with distilled water to remove the excess stain and dried. To extract the stain, wells were treated with 200 µl of ethanol (90%) for 5 min. The absorbance of each was measured at 595 nm. The amount of biofilm inhibition was calculated relative to the amount of biofilm that was developed in the presence of the phage lysate and antibiotics and compared with the controls containing biofilms by untreated bacterial cultures (Student’s t-test was performed). Un-inoculated media was taken as a blank (Figure 4). And values of the blank measurement were deducted from the test values to remove the background.
Data analysis: All of the experiment for growth and biofilm inhibition were performed in triplicate. Data was analyzed and values of mean, standard deviation and standard error were calculated in Microsoft Office Excell 2010. To calculate the significant difference between two independent means Student’s t-test was performed at p <0.05.
Results![]()
Screening of the phages against MRSA and E. coli strains
The different cow manure samples (n=4) prospectively containing the phages were initially enriched with S. aureus and the environmental isolate E. coli. The phages isolated (enriched phage lysate) showed a various lysis pattern against the clinical MRSA and E. coli. From a total of four phage lysate samples, two showed the inhibition of MRSA growth demonstrated through either clear or turbid plaque formation on bacterial lawn (Figure 1 a). One of them has shown complete inhibition E. coli growth by forming clear plaques whereas a turbid plaque formation was observed in the fourth phage lysate sample against E. coli (Figure 1 b).
The phage lysate samples were diluted to 1:100 and allowed to infect clinical bacterial cultures (E. coli and MRSA) at an OD600 of 0.5 (~ 106 CFU/ml) on nutrient agar double layer agar plate. Numerous plaques were observed after incubation which were counted and the enriched phage lysate showed 5X107 PFU /ml (Figure 2 a, b). The control plate (un-treated bacterial culture) showed a confluent growth over the surface of nutrient agar plate (Figure 2 c & d).
Time-dependent lytic assay of bacteriophages against MRSA and E. coli
The lytic efficacy of the phage lysates was tested against clinical MRSA and E. coli strains in the specified period of time. After the enrichment and incubation of phages with their specific strains, the OD600 was measured every 30 minutes for 3 hours. The controls contained cultures of bacteria without phages to monitor their normal growth pattern. After the growth of one hour a slight increase in turbidity was observed but started to decline in the next hour and reached to the OD600 observed at the initial hour of growth. The cultures in the control (un-treated bacterial culture) showed steady rise in OD600 indicating the normal growth pattern of bacteria in the absence of phage (Figure 3 a & b).
Less virulent phages have anti-biofilm competency
Lytic phages are known to reduce the biofilm formation or destroy the pre-formed biofilms. Here in our study, we tested the phage lysate samples from the turbid plaques (indicated by dashed red arrows in Figure 1) to demonstrate the inhibition of biofilms formed by MRSA and E. coli in a micro-titer well plate. As the concentration of the phages increased, a decrease in the CV absorbance was noticed at 595 nm, indicating the phages can successfully prevent the biofilm formation at their lowest dilutions (Figure 4). A significant inhibition of biofilm was observed when bacteria were incubated with the controls (antibiotics), ½ of the minimum inhibitory concentration (MIC) of Chloramphenicol (CMP) and Ampicillin (AMP) (Figure 4). The sensitivity and MIC of the selected antibiotics were determined for MRSA and E. coli before their test to inhibit the biofilm formation. The un-treated bacterial culture did not contain any anti-bacterial compounds and showed no growth inhibition when compared with the growth of bacterial culture incubated with the phages. Student’s t-test was performed to check the significance of biofilm inhibition. A significant difference in the biofilm inhibition was observed between the bacterial cells treated with phages and untreated bacterial culture, indicated by asterisks in figure 4 bar diagrams. The control (C) showed full biofilm formation as indicated by high CV absorbance.
Figures & Tables


Discussion![]()
Bacteriophages efficiently inhibit the growth of MRSA and E. coli
The transmission of S. aureus and E. coli infections from animals to human is reported worldwide [7,8]. S. aureus is a common colonizer of human that could results in various infections and can also cause mastitis in cows [8]. Based on the abundant presence of the host bacteria, we tried to hunt their parasites (bacteriophages) in the cow manure samples. In the present study, the presence of phages was screened in four different cow manure samples. For that purpose the filtrates of the cow manure samples (presumably containing the phages) were enriched with the specific host strains i.e. S. aureus and E. coli isolated from environment and later were checked for their lytic efficacy against the clinical strains, MRSA and E. coli. From a total of four phage lysate samples, two showed the inhibition of MRSA and E. coli growth demonstrated through clear plaque formation on bacterial lawn (Figure 1 a & b). Two of phage lysate showed incomplete inhibition of growth by forming turbid plaques (Figure 1 a & b). Our results support the previously described studies which showed the isolation and abundance of specific phages from the cattle samples against the gram positive and gram negative bacteria [16,18,19]. A varied pattern of lysis was demonstrated that phage lysates can be further assayed for their anti-bacterial activity.
Lytic abilities of phages and prospective use in phage therapy
According to the WHO 2017 report, the antibiotic resistant bacteria like E. coli and MRSA are placed among the top priority pathogen and considered as a threat to healthcare by causing different diseases in humans and cattle [20]. The development of new antibiotics is commonly used strategy to control the antibiotic resistance and prevent its spread. However, the alternates of antibiotics like natural herbal or homeopathic medicines, anti-microbial peptides and phage therapy can serve as the best therapeutic options to eliminate the antibiotic resistant pathogens. In this study, when the selected phage lysate was incubated with the cultures of MRSA and E. coli, a significant reduction on the optical density/growth was monitored indicating the isolated phages encompass the lytic effect on the selected clinical strains (Figure 3). Several mechanisms are known for the bacterial lysis by phages. They all ultimately results in the degradation of peptidoglycan by phage encoded endolysins which mimic glycosylase (lysozyme), endopeptidase, amidase, and transglycosylase enzymes. Endolysin along with membrane degrading holins results in the lysis of bacterial cells [21].
Anti-biofilm phages have industrial and medicinal applications
Bacteria in biofilms are often resistant to antibiotics due to thick glycocylax and persisting cells [22]. Biofilms in medical implants and industries are nuisance for the production and difficult to get removed [23]. The use of phages as an anti-biofilm agents in the medical inserts and food industries is described in previous studies [24]. In the present study the phages selected from the turbid plaques have demonstrated the complete inhibition of the biofilm formation by MRSA and E. coli in polystyrene micro-well plate (Figure 4). We postulate here that although phages in the turbid plaques have less inhibitory effect on the growth of the bacteria but they can successfully inhibit the biofilm formation perhaps by enzymes like polysaccharide de-polymerase which actually degrades the matrix of biofilms. The biofilm formation needs a certain threshold of cell population to make biofilm [25]. From the obtained results we can also presume that phages can halt the bacterial growth at the level that they cannot achieve their required density to establish quorum-sensing derived biofilm.
This study has successfully led to the isolation of the lytic phages from the different cow manure samples and successfully demonstrated their antibacterial activity against MRSA and E. coli strains from clinical origin. The inhibition of biofilm formation by the isolated phages indicates that they can be used a suitable biocontrol agents against targeted pathogens like MRSA and E. coli both in both the industrial and medical fields. However further studies in details are required to exploit the potential of phages as a controlling agent against the selected bacterial pathogens.
Authors' Contribution
Nimerta Kumari designed, executed and wrote this project as a principal investigator. Bushra Patoli did the PCR amplification of mecA gene and helped in writing up this manuscript. Atif Patoli did the analysis of results and execution of the laboratory work. Shagufta Jabeen helped in biofilm inhibition assay and reviewed the manuscript.
The authors declare that they have no competing interests.
References![]()
- Twort FW. An investigation on the nature of ultra-microscopic viruses. The Lancet, (1915); 186(4814): 1241-1243.
- Barrangou R, Fremaux, Christophe, Deveau, Hélène, Richards, Melissa, Boyaval, Patrick, Moineau, Sylvain, Romero, Dennis A., Horvath, Philippe. CRISPR Provides Acquired Resistance Against Viruses in Prokaryotes. Science, (2007); 315(5819): 1709-1712.
- Henry M, Debarbieux L. Tools from viruses: bacteriophage successes and beyond. Virology, (2012); 434(2): 151-161.
- Lederberg EM, Lederberg J. Genetic studies of lysogenicity in Eschericia coli. Genetics, (1953); 38(1): 51-64.
- Sulakvelidze A, Alavidze Z, Morris JG, Jr. Bacteriophage therapy. Antimicrob Agents Chemother, (2001); 45(3): 649-659.
- Joerger RD. Alternatives to antibiotics: bacteriocins, antimicrobial peptides and bacteriophages. Poultry science, (2003); 82(4): 640-647.
- Juhász-Kaszanyitzky E, Jánosi S, Somogyi P, Dán A, van der Graaf-van Bloois L, et al. MRSA transmission between cows and humans. Emerging infectious diseases, (2007); 13(4): 630-632.
- Cuny C, Wieler LH, Witte W. Livestock-Associated MRSA: The Impact on Humans. Antibiotics (Basel, Switzerland), (2015); 4(4): 521-543.
- Ramírez-Castillo FY, Moreno-Flores AC, Avelar-González FJ, Márquez-Díaz F, Harel J, et al. An evaluation of multidrug-resistant Escherichia coli isolates in urinary tract infections from Aguascalientes, Mexico: cross-sectional study. Annals of clinical microbiology and antimicrobials, (2018); 17(1): 34-34.
- Azizian Reza AJF, Razieh Amini5, Karimi Sajedeh,Sadeghifard Norkhoda, Sekawi Zamberi, Nasser Ahmad, Taherikalani Morovat, Askari Hassanand Pakzad Iraj. Dynamics of Bacteriophages as a Promising Antibiofilm Agents. JOURNAL OF PURE AND APPLIED MICROBIOLOGY,, (2014); 8(2): p. 1015-1019.
- O'Toole G, Kaplan HB, Kolter R. Biofilm formation as microbial development. Annual review of microbiology, (2000); 5449-79.
- Curtin JJ, Donlan, R. M. Using bacteriophages to reduce formation of catheter-associated biofilms by Staphylococcus epidermidis. Antimicrob Agents Chemother, (2006); 50(4): 1268-1275.
- Kevin A. Hughes IWSaMVJ. Biofilm susceptibility to bacteriophage attack :the role of phage-borne polysaccharide depol yrnerase. Microbiology, (1998); 1443039-3047.
- Sambrook JR, D. W. . Molecular Cloning: A Laboratory Manual,NY, USA (2001). Cold Spring Harbor Laboratory Press: New York,, (2012); 4th ed.
- Chhibber S, Kaur T, Kaur S. Essential role of calcium in the infection process of broad-spectrum methicillin-resistant Staphylococcus aureus bacteriophage. Journal of Basic Microbiology, (2014); 54(8): 775-780.
- Jurczak-Kurek A, Gasior T, Nejman-Falenczyk B, Bloch S, Dydecka A, et al. Biodiversity of bacteriophages: morphological and biological properties of a large group of phages isolated from urban sewage. Sci Rep, (2016); 634338.
- S.A. DOSS GSTaSGBA. Effect of sub-inhibitory concentrations of antibiotics on the virulence of Staphylococcus aureus. Journal of Applied Bacteriology, (1993); 75123-128.
- Callaway TR, Edrington TS, Brabban AD, Keen JE, Anderson RC, et al. Fecal Prevalence of Escherichia coli O157, Salmonella,Listeria, and Bacteriophage Infecting E. coli O157:H7 inFeedlot Cattle in the Southern Plains Region of theUnited States. FOODBORNE PATHOGENS AND DISEASE, (2006); 3(3).
- Synnott AJ, Kuang Y, Kurimoto M, Yamamichi K, Iwano H, et al. Isolation from sewage influent and characterization of novel Staphylococcus aureus bacteriophages with wide host ranges and potent lytic capabilities. Appl Environ Microbiol, (2009); 75(13): 4483-4490.
- Rodrigo-Troyano A, Sibila O. The respiratory threat posed by multidrug resistant Gram-negative bacteria. Respirology, (2017); 22(7): 1288-1299.
- Cahill J, Young, R. Phage Lysis: Multiple Genes for Multiple Barriers. Advances in Virus Research, (2019); 10333-70.
- Stewart PS. Mechanisms of antibiotic resistance in bacterial biofilms. International journal of medical microbiology : IJMM, (2002); 292(2): 107-113.
- Hall-Stoodley L, Costerton JW, Stoodley P. Bacterial biofilms: from the Natural environment to infectious diseases. Nature Reviews Microbiology, (2004); 2(2): 95-108.
- Neguţ AC, Chifiriuc M-C, Săndulescu O, Streinu-Cercel A, Oprea M, et al. Bacteriophage-driven inhibition of biofilm formation in Staphylococcus strains from patients attending a Romanian reference center for infectious diseases. FEMS Microbiology Letters, (2016); 363(18).
- Kjelleberg S, Molin S. Is there a role for quorum sensing signals in bacterial biofilms? Current Opinion in Microbiology, (2002); 5(3): 254-258.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0