

Full Length Research Article
Reserve Carbohydrate Metabolism in Crabtree-Negative and –Positive Yeasts at Different Carbon Sources
Tülay Turgut Genç
Adv. life sci., vol. 8, no. 1, pp. 47-51, November 2020
*- Corresponding Author: Tülay Turgut Genç (Email: tturgutgenc@comu.edu.tr)
Authors' Affiliations
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: The fermentation of sugars into ethanol even in the presence of oxygen is referred to as the Crabtree effect. The yeast cells displaying Crabtree effect are indicated as Crabtree-positive yeast. Saccharomyces cerevisiae is Crabtree positive and Debaryomyces occidentalis is Crabtree-negative yeast which does not have Crabtree effect. The reserve carbohydrate metabolism is different in Crabtree-positive and Crabtree-negative yeast cells. The present study aimed to determine the trehalose and glycogen accumulation patterns both in Crabtree-positive and Crabtree-negative yeast species.
Methods: In this research, trehalose and glycogen contents of S. cerevisiae and D. occidentalis yeast species were examined in a time course manner in three different carbon sources: glucose, galactose and glycerol. Firstly, yeast cells were grown in rich media supplemented with glucose then all washed and switched to fresh cultures including glucose, galactose and glycerol.
Results: In S. cerevisiae yeast cells the overnight accumulated trehalose degraded very rapidly after non-fermentable carbon source replenishment, but this took place in a long time, nearly two days, in D. occidentalis yeast cells. However, whenever D. occidentalis yeast cells shifted to glycerol, all the accumulated trehalose degraded within the twelve hours. Glycogen accumulation in D. occidentalis yeast cells is lower than S. cerevisiae yeast cells both in fermentable and non-fermentable carbon sources.
Conclusion: Results indicated that glycogen and trehalose accumulation patterns are completely different in D. occidentalis than S. cerevisiae. Crabtree-negative yeast cells generally, prefer to accumulate glycogen instead of trehalose as reserve carbohydrate. But in our research we proved that Crabtree-negative yeast D. occidentalis, accumulates more trehalose than S. cerevisiae yeast cells in non-fermentable carbon sources.
Keywords: Trehalose; Glycogen; Debaryomyces occidentalis; Saccharomyces cerevisiae; Crabtree effect
Introduction![]()
Saccharomyces cerevisiae is the most commonly used species in biotechnological applications and yeast research. But the genetic and metabolic properties of S. cerevisiae is not similar to other yeast species belonging to other genera. Among the nearly 1500 identified yeast species, a few yeast species are used in biotechnological applications. The physiological and metabolic properties of many non-saccharomyces yeast species have not yet been characterized [1]. D. occidentalis can grow on unusual carbon sources and some strains produce killer toxin that is lethal to sensitive S. cerevisiae strains [2]. D. occidentalis do not produce ethanol at aerobic conditions so it is Crabtree-negative, and S. cerevisiae can produce ethanol at aerobic conditions after glucose pulse, so it is Crabtree-positive [3]. Crabtree-positive yeast shows high glucose consumption and so they have high glycolytic rate than Crabtree-negative yeasts. In addition, Crabtree-negative yeasts generally produce and accumulate more glycogen as a reserve carbohydrate [4].
Trehalose and glycogen are the two reserve carbohydrates of the yeast S. cerevisiae [5]. Both carbohydrates are accumulated upon nutrient deprivation and external stress [6]. These non-optimal growth conditions result in a decrease in the growth rate and elongation of the cell cycle [5, 7, 8]. In fed-batch culture trehalose and glycogen are accumulated in the G1 phase [9]. There is no detailed information on the biosynthesis of trehalose and glycogen accumulation in D. occidentalis. Therefore, in this study, we make a comparable analysis of glycogen and trehalose accumulation in D. occidentalis and S. cerevisiae yeast strains in a time course manner in fermentable and non-fermentable carbon sources.
Methods![]()
Yeast strains and growth conditions
D. occidentalis yeast species (DBVPG-6722) is obtained from DBVPG collection (Perugia University, Italy) and S. cerevisiae strain BY4741 (BY4741 (Mat a, his3Δ1; leu2Δ0; met15Δ0; ura3Δ0) is obtained from EUROSCARF (Frankfurt, Germany).
For stating the doubling times (dt) and specific growth rates (μ) of yeast strains, yeast cells were grown in rich medium (YP) containing 1% yeast extract, 2% peptone and the appropriate carbon sources: 2 % glucose, 2 % galactose and 2 % glycerol. The optical densities of yeast cultures (OD600) were measured spectrophotometrically every 2 h and used for calculation.
In order to determine the time-course dependent accumulation of trehalose and glycogen, the yeast strains were pre-cultured in YP medium supplemented with 2% dextrose (YPD), 2% galactose (YPGAL) and 2 % glycerol (YPGL) up to the stationary stage. Then, these cultures were used to inoculate the yeast strains to the fresh YPD, YPGAL and YPGL media. The yeast samples were removed at every 6 h and the accumulation of trehalose and glycogen were determined enzymatically. Cultivation of the yeast strains were done in duplicates and the experiments were repeated three times. Hence glycogen or trehalose contents given in the tables and figures are the average values of the six independent measurements.
Trehalose and glycogen assay
Trehalose and glycogen accumulation of yeast cells were determined as described previously [10]. Yeast cells (50-60 mg cell wet mass) were removed and washed with ice-cold water and then resuspended in 250 µl of 0.25 M Na2CO3 and boiled for 2 h. Then 150 µl of 1M acetic acid and 600 µl of 0.2 M sodium acetate pH 5.2 were added and divided into two parts. One half of the cell mixture was incubated at 37 °C for 18 h in the presence of 3mU trehalase enzyme (Sigma, T8778, 0.25U ml-1) for trehalose assay. The second half of the suspension was incubated at 57 °C for 18 h with continuous shaking in the presence of 1 U/ml amyloglucosidase enzyme from Aspergillus niger (Sigma, 10115) for the glycogen assay. The amount of the liberated glucose was determined enzymatically via the glucose oxidase-peroxidase system (GOD-POD assay) using a commercial kit (Fluitest®- GLU, Biocon, Germany) [11]. The determined trehalose and glycogen contents of yeast cells were given as milligram of glucose equivalent per gram of wet mass (mg/g) of the yeast cells in all figures. The standard error values are less than 10%, so the error bars were not displayed in the graphs.
Results![]()
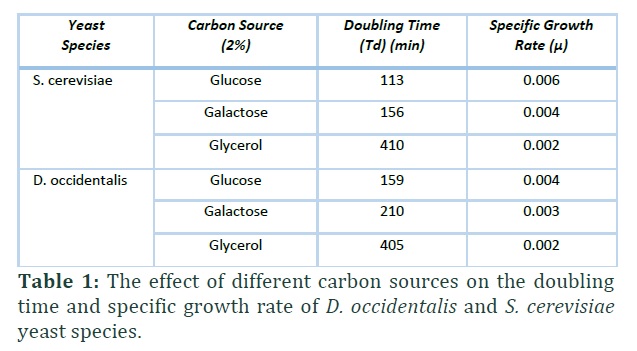
Several factors have been implied as initiator of trehalose and glycogen synthesis, like the glucose flux, the growth rate, the intracellular glucose-6-phosphate levels and glucose concentration in the medium. In order to understand the effect of different carbon sources on the glycogen and trehalose accumulation, growth rate of yeast species in these carbon sources were determined. As shown Table 1, doubling time decrease, that means yeast cells enter the stationary phase very rapidly, so the specific growth rate increase. In fermentable carbon sources, glucose and galactose, specific growth rates of S. cerevisiae were greater than D. occidentalis. In both yeast species specific growth rate decreased to 0.002 when the carbon sources became glycerol.
In order to determine time-course dependent accumulation of reserve carbohydrates and the fluctuations in accumulations depending on the carbon source, yeast cells were pre-cultured in rich medium supplemented with appropriate carbon sources.
The degradation of previously accumulated trehalose (15.52±4.3 mg glucose/gr cell wet mass) took place very rapidly (less than 12 h) when the overnight grown yeast cells were inoculated into fresh YPD medium in S. cerevisiae cells (Figure 1). The accumulation of trehalose both in fermentable and non-fermentable carbon sources started after 24 h that is late-logarithmic phase. The accumulated trehalose level in glucose grown yeast cells (1.1±0.2 mg glucose/gr cell wet mass) was three- and eight-fold lower than galactose (3.4 ±0.4 mg glucose/gr cell wet mass) and glycerol (9.13±1.6 mg glucose/gr cell wet mass) grown cells at 24 h, respectively. In glycerol grown yeast cells the accumulation of trehalose reached to top level (12.4 ±2.3 mg glucose/gr cell wet mass) at 36 h and started to decrease rest of the growth period, but the level of trehalose in glucose and galactose continued to increase after 24 h (Figure 1). The trehalose levels became 9.4±0.2 and 12.19±1.7 mg glucose/gr cell wet mass at the end of 72 h glucose and galactose growth, respectively.
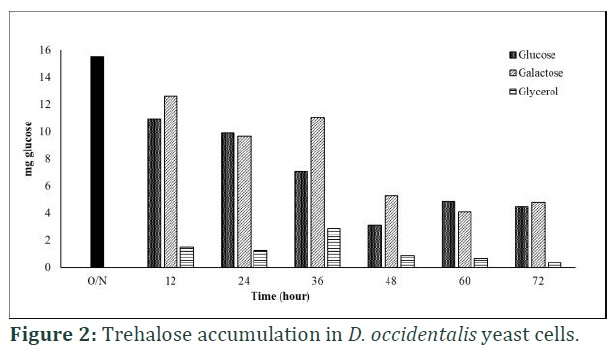
In D. occidentalis yeast cells, the previously accumulated trehalose (15.52±2.8 mg glucose/gr cell wet mass) degraded very slowly. It decreased to 3.15±0.3 mg glucose/gr cell wet mass in glucose at 48 h later and 4.1±0.5 mg glucose/gr cell wet mass in galactose at 60 h later (Figure 2). The yeast cells started to accumulate new trehalose at 60 h and 72 h after glucose and galactose replenishment, respectively. But the overnight accumulated trehalose degraded very rapidly into 1.5±0.2 mg glucose/gr cell wet mass after inoculation to fresh glycerol supplemented culture. Trehalose level increased to maximum level (2.9±0.3 mg glucose/gr cell wet mass) at 36 h and then started to decrease during 72 h growth period.
The overnight accumulation level of glycogen in S. cerevisiae yeast cells was nearly two times more than D. occidentalis yeast cells that is 4.23±0.3 and 2.59±0.4 mg glucose/gr cell wet mass, respectively. The previously accumulated glycogen was broken down into 0.44±0.1 mg glucose/gr cell wet mass in glucose and 0.95±0.2 mg glucose/gr cell wet mass in glycerol after 12 h from switching. However, in galactose grown cells degrade glycogen with in the 36 h (Figure 3). Accumulation of newly synthesized glycogen was started in glycerol grown S. cerevisiae yeast cells after 12 h and reached to maximum level, 10.45±1,9 mg glucose/gr cell wet mass, within 24 h. The accumulated glycogen in post-diauxic stage, immediately was degraded completely after 36 h later (Figure 3).
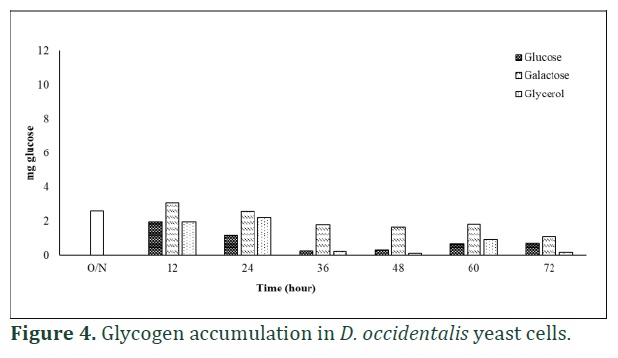
In D. occidentalis yeast cells, the previously accumulated glycogen degraded slowly in all used carbon sources except galactose. The level of glycogen in galactose grown yeast cells became nearly constant during 72 h growth period (Figure 4). Glycogen accumulation were not detected both in S. cerevisiae and D. occidentalis yeast cells during stationary phase (after 48 h), but at the end of 60 h, glycogen was started to accumulate both in yeast cells.
Figures & Tables


Discussion![]()
It is known that, S. cerevisiae yeast cells accumulates trehalose when the environmental conditions become unfavorable. When the nutrients become depleted yeast cells enter to stationary phase and start to accumulate trehalose. As in the case, in our research trehalose accumulated in the stationary phase both in glucose and galactose carbon supplies. However, in glycerol, trehalose level increased during 36 h and then dropped to low levels. The level of reserve carbohydrates can be regulated by growth rate of the yeast cells and nutrient flux [12]. Specific growth rate of S. cerevisiae yeast cells was too low in glycerol (µ=0.002). In addition, glycerol itself cannot activate the glycolytic flux. Glucose and galactose transported by means of the hexose transporters that play an important role in efficient Crabtree effect [13]. Glycerol transporters, Stl1p (Sugar transporter-like 1) are Glycerol proton symporter of the plasma membrane and similar to hexose transporters but not activated by glucose flux [14]. Glycerol also synthesized by yeast cells under different stress conditions, especially osmotic stress via HOG pathway [15]. At the late logarithmic stage both trehalose and glycogen accumulated in high levels S. cerevisiae yeast cells, and then degraded. This may be because of the synthesis of glycerol within the cells and stop the utilization of external glycerol as a respond to entrance in stationary phase. D. occidentalis yeast cells are resistant to osmotic stress, so they are known as osmotolerant yeasts. In our research we showed that the accumulated trehalose degraded in a very long time. Trehalose level in the cells maintained it in a certain level, not all trehalose was degraded. This may be the one way of D. occidentalis yeast cells to resist the stressful conditions.
Türkel (2006) was reported that glycogen accumulation starts after 48 h later, at mid-stationary phase, in glucose grown S. cerevisiae cells [16]. In our research the glycogen accumulation recorded after 24 h later. This may be related with the cell densities at that time. In our research, the pattern of glycogen and trehalose accumulation in S. cerevisiae yeast cells in fermentable carbon sources was more or less similar. However, in D. occidentalis, especially the trehalose accumulation pattern, was different in glucose and galactose growing cells. In non-fermentable carbon source, glycerol, D. occidentalis did not synthesize glycogen and trehalose during logarithmic and stationary phase. It may accumulate another metabolite, such as mannitol or something else. For example, in some Pichia species accumulates cererosides and glycosides instead of trehalose and glycogen, under unfavorable growth conditions These results shown that the reserve carbohydrate metabolism and regulation in Crabtree positive and Crabtree negative yeast cells are completely different.
Different factors such as decrease in the glucose-6-phospahate level, decrease in the glucose flux and the depletion of glucose in the growth medium, cause decrease in the growth rate and therefore the accumulation of reserve carbohydrates in Crabtree positive S. cerevisiae yeast cells [17, 18]. Crabtree-negative yeast cells generally accumulate glycogen instead of trehalose as reserve carbohydrate [4]. The depletion of glucose caused the accumulation of trehalose in S. cerevisiae yeast cells but not in D. occidentalis yeast cell. Also S. cerevisiae yeast cells started to accumulate trehalose when the galactose in the growth medium consumed that means concentration of glucose-6-phosphate decreased with in the cell. However, in Crabtree negative yeast, D. occidentalis, trehalose accumulation was not effected from the galactose concentration and glucose-6-phospahate concentration. These results indicate that in D. occidentalis neither glucose depletion in the growth medium nor drop in the glucose-6-phosphate level with in the cell can create a signal for trehalose accumulation. There may be another intrinsic factor with in the either signal transduction pathway or trehalose metabolism in D. occidentalis yeast cells. This unknown factor also may inhibit the synthesis of both trehalose and glycogen in the D. occidentalis in nonfermentable growth conditions.
Acknowledgment
This work was supported by Çanakkale Onsekiz Mart University the Scientific Research Coordination Unit, Project number: 2008-44.
The author declares that there is no conflict of interest regarding the publication of this paper.
References![]()
- Kregiel D. Physiology and metabolism of Crabtree-negative yeast Debaryomyces occidentalis. Food Chemistry and Biotechnology, (2008); 72(1029): 35–43.
- Chen KC., Csikasz-Nagy A, Gyorffy B, Val J, Novak B, Tyson JJ. Kinetic analysis of a molecular model of the budding yeast cell cycle. Molecular Biology of the Cell, (2000); 11: 369–91.
- De Deken RH. The Crabtree effect: A regulatory system in yeast. Journal of General Microbiology, (1966); 44: 149–56.
- Van Urk H, Voll WSL, Dcheffers WA, van Dijken J. Transient-state analysis of metabolic fluxes in Crabtree-positive and Crabtree-negative yeasts. Applied and Environmental Microbiology, (1990); 56: 281–87
- Lillie SH, Pringle JR. Reserve carbohydrate metabolism in Saccharomyces cerevisiae: Responses to nutrient limitation, Journal of Bacteriology, (1980); 143: 1384–94.
- Parrou JL, Teste MA, François J. Effects of various types of stress on the metabolism of reserve carbohydrates in Saccharomyces cerevisiae: genetic evidence for a stress-induced recycling of glycogen and trehalose. Microbiology, (1997); 143(6): 1891–1900.
- Carter BL, Jagadish MN. Control of cell division in the yeast Saccharomyces cerevisiae cultured at different growth rates. Experimental Cell Research, (1978); 112: 373–83.
- Johnston GC, Singer RA. Ribosomal precursor RNA metabolism and cell division in the yeast Saccharomyces cerevisiae. Molecular General Genetics, (1980); 178: 357–60.
- Sillje HHW, Ter Schure EG, Rommens AJM, Huls PG, Woldringh CL, Verkleij AJ, Boonstra J, Verrips CT. Effects of different carbon fluxes on G1 phase duration, cyclin expression and reserve carbohydrate metabolism in Saccharomyces cerevisiae. Journal of Bacteriology, (1997); 179: 6560–65.
- Parrou JL, François J. A simplified procedure for a rapid and reliable assay of both glycogen and trehalose in whole yeast cells. Analytical Biochemistry, (1997), 248: 186–88.
- Goldstein A, Lampen JO. Beta-D-fructofuranoside Fructohydrolase from Yeast. Methods in Enzymology, (1975); 42: 504–11.
- Paalman JWG, Verwaal S, Slofstra SH, Verkleij AJ, Boonstra J, Verrips CT. Trehalose and glycogen accumulation is related to the duration of the G1 phase of Saccharomyces cerevisiae. FEMS Yeast Research, (2003); 3(3): 261–68.
- Lin Z, Li WH. Expansion of hexose transporter genes was associated with the evolution of aerobic fermentation in yeasts. Molecular Biology and Evolution, (2011); 28(1): 131–42.
- Ferreira C, van Voorst F, Martins A, Nevels L, Oliveria R, Kielland-Brandt MC, Lucas C, Brandt A. A member of the sugar transporter family, Stl1p is the glycerol/H+ symporter in Saccharomyces cerevisiae. Molecular Biology of the Cell, (2005); 16(4): 2068–76.
- Hohmann S. Osmotic stress signaling and osmoadaptation in yeasts. Microbiology and Molecular Biology Reviews, (2002); 66(2): 300–72.
- Türkel S. Comparative analysis of glycogen and trehalose accumulation in Methylotrophic and nonmethylotrophic yeasts. Microbiology, (2006), 75(6): 737-41.
- Enjalbert B, Parrou JL, Vincent O, Francois J. Mitochondrial respiratory mutants of Saccharomyces cerevisiae accumulate glycogen and readily mobilize it in a glucose-depleted medium. Microbiology, (2000); 146: 2685–94.
- François J, Parrou J. Reserve carbohydrates metabolism in the yeast Saccharomyces cerevisiae. FEMS Microbiology Reviews, (2001); 25: 125–45.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0


