

Full Length Research Article
The Association of Nine Novel Variants in the ZEB2 Gene with Serum Levels of TNF-α, TGF-β and IFN-γ
Zahraa Hashim Rabae, Jawdat N. Gaaib*
Adv. life sci., vol. 9, no. 2, pp. 202-206, July 2022
*– Corresponding Author: Jawdat N. Gaaib (Email: Jawdat.noori@uokerbala.edu.iq)
Authors' Affiliations
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: ZEB1 and ZEB2, are expressed by a variety of immune cells of both myeloid and lymphoid lineages, they act to regulate important transcriptional networks necessary for cell differentiation, maintenance, and function. The study was carried out in the Department of Clinical Laboratories / College of Applied Medical Sciences / University of Kerbala during the period from January 2021 to September 2021. The study aimed to shed light on the association between the variation of the ZEB2 gene and serum levels of some cytokines (TNF-α, TGF-β, INF-γ) involved in immune cell differentiation in the Iraqi population.
Methods: A total of 80 healthy volunteers, randomly chosen, were enrolled in this study, with an age ranged 20-50 years at the time of the investigation. The variation of the ZEB2 gene was investigated by the Sanger sequencing method. The serum levels of TNF-α, TGF-β and INF-γ were measured by ELISA.
Results: The results identified presence of nine novel non-registered variants in 27 samples, 5 out of 9 variants were detected in exon1 region, while the other 4 were detected in the 5`UTR region. These variants might be of interest to the ZEB2 gene studies due to their presence in the coding and regulatory regions that regulate the gene expression.
Conclusion: The two variants, 144520053 G/A and 144520162 T/A, among the nine novel variants might be the most important variants because both showed significant effects on some study parameters.
Keywords: Novel variants; ZEB2; TNF-alpha; TGF- beta; IFN-gamma
Introduction![]()
The ZEB protein, the zinc-finger E-box–binding family of transcription factors (TFs), is an essential modulator of the epithelial-to-mesenchymal transition (EMT) during embryonic development. The ZEB2 gene was formerly thought to be responsible for (EMT via repressing epithelial genes [1-3]. The ZEB gene family consists of two types: ZEB1 and ZEB2. Dendritic, macrophages, monocytes, B cells, T cells, and Natural Killer (NK) cells, belonging to both the myeloid and lymphoid lineages, are among the immune cells that express these genes [4]. The ZEBs regulate critical transcriptional networks required for cell development, maintenance, and function in these cells. In humans, the ZEBs typically serve as transcriptional repressors by collaborating with stimulated Small Mothers Against Decapentaplegic (SMAD) proteins [5-8].
The ZEB2 gene has an important role in the immune system, especially on cytokines. It encodes for low molecular weight non-structural proteins that regulate inflammatory response and hematopoiesis in a complex way [9, 10]. The encoded proteins include interleukins, interferons, mesenchymal growth factors, the chemokine family, and the tumor necrosis factor family [10, 11]. Every cell, with the exception of RBCs, produces cytokines and responds to them [12]. The ZEB2 gene has been characterized as a novel player in the transcriptional control of lymphocyte differentiation [13]. This study aimed to shed light on the association between the variation of the ZEB2 gene and serum levels of some cytokines (TNF-α, TGF-β, INF-γ) involved in immune cell differentiation in the Iraqi population.
Methods![]()
Blood Samples
A total of 80 healthy volunteers, randomly chosen, were enrolled in this study, with an age range of 20-50 years. Individuals who suffered from any history of disease or conditions that affect the immune system, such as malnutrition, infections (HIV, CMV, measles, HBV, HCV, HAV, HEV, etc.), chronic diseases (hepatic, renal, pulmonary, etc.), malignancies, surgery, trauma, chemotherapy, wounds, any medication influencing the immune system, infections of upper or lower respiratory tract, gastrointestinal tract or urinary tract infections, were excluded. The blood samples were collected from the individuals in Al-Imam Al-Sadiq hospital in Babylon city in Iraq, an ethical consent was signed by each volunteer. Two milliliters of the venous blood sample were obtained from each participant in a gel tube, and the blood was drawn by disposable syringes in a sterile condition. The collected blood was centrifuged to separate serum to be used later for TNF-α, IFN-γ and TGF-β assessment.
DNA extraction and molecular detection
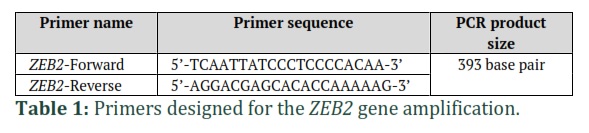
The DNA was extracted from the blood samples by using the gSYNC™ DNA Extraction Kit (Geneaid Company) The primers used in this study were designed by Primer3plus bioinformatics tool (https://www.bioinformatics.nl/cgi-bin/primer3plus/primer3plus.cgi). Conventional PCR was used to amplify the DNA fragments that contain the targeted sequence (393 bp) of the ZEB2 gene located on the long arm of chromosome 2. A total volume of 25 μl was used in the PCR reaction (3 µl DNA, 1µl from each primer, 12.5 µl master mix, and 7.5 µl nuclease free water). The PCR program used to amplify the target sequence consisted of 35 cycles, each cycle included denaturation for 30 seconds at 94ºC, annealing for 30 seconds at 59ºC and extension for 30 seconds at 72ºC . Agarose gel electrophoresis was used to separate PCR product bands on 2% agarose gel stained by fluorescent Redsafe dye. The gel electrophoresis system was set at 70 volts for 60 minutes, then the gel was displayed under UV transilluminator.
Nucleotides sequencing and analysis
All the 80 PCR product samples were sent to the Macrogen public biotechnology corporation, South Korea, to perform nucleotide sequencing. Sanger sequencing method was applied using an automated DNA sequencer. The results of sequencing were received by email, analyzed manually by using bioinformatics tools and aligned with human reference gene sequences already deposited on the National Center for Biotechnology Information (NCBI). The alignments were done by using the Basic Local Alignment Search Tool (BLAST) provided by NCBI. The sequenced region of the target gene was analyzed by using Molecular Evolutionary Genetics Analysis X (MEGAX), in which the multiple alignments of the sequences were performed by the CLUSTALW program to confirm the presence of variations detected by the BLAST tool [14]. Locations of all variants detected in the current study were reported and checked to identify the type of variants and predict their functional consequences by using tools provided by Ensemble Genome Browser. In addition, the aforementioned browser was also used to identify presence of known, unknown, and novel variants in the current study.
Enzyme Linked Immunosorbent Assay (ELISA)
The ELISA kits (Elabscience company) were used in this study to determine the serum levels of human TNF-α, TGF-β and IFN-γ.
Statistical analysis
To determine the effect of different groups on the study parameters, the Statistical Analysis System (SAS, 2012) was utilized. To make comparison between means, the Least Significant Difference –LSD test (ANOVA) was performed. The Chi-square test was employed to make comparisons between percentages.
Results
![]()
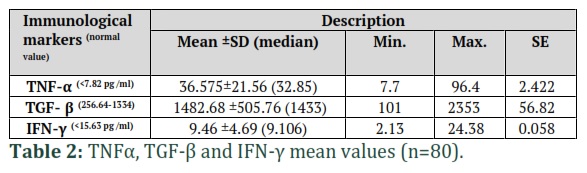
The age of included 80 individuals ranged from 20-50 years old. Of the participants 46% were between 20–29 years old, 38 % were between 30–39 years old, and 16% were 40–49 years old. Apparently healthy individuals aged 20 to 50 years old were selected randomly to be enrolled in this study because they had an optimal immune system compared to individuals over 50 years old. The sample mean± SD for age was 31.2±6.87 years (ranged 20-48 years). The number of males was 60 (75%), and the number of females was 20 (25%) among the individuals. Results of the TNF-α serum level showed that its mean was 36.575, the standard deviation recorded 21.56, the median was 32.85, the standard error was 2.422, the minimum and maximum values were 7.7 pg/ml and 96.4 pg/ml, respectively. While the normal range value of TNF-α was <7.82 pg /ml (Table 2). The IFN-γ level revealed a mean of 9.46, the standard deviation recorded 4.69, the median was 9.106, the minimum and maximum values were 2.13 pg/ml, and 24.38 pg/ml, respectively. The standard error was 0.058, while the normal range value of IFN-γ was <15.63 pg/ml (Table 2). The findings also showed that the mean of TGF-β was 1482.68, the standard deviation recorded 505.76, the median was 1433, the minimum and maximum values were 101 and 2353, respectively, and the standard error was 56.82. While normal range values of TGF-β was 256.64 -1334 (Table 2).
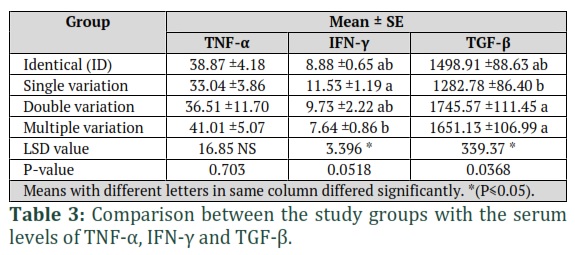
Conventional PCR was used to amplify the DNA target region in the ZEB2 gene, including 5`UTR region (166 bp) and exon 1 region (181 bp). The PCR product (393 bp) was detected in all of the study samples, this indicates the presence of the target region in the ZEB2 gene. The genetic analysis of the 80 samples showed that 34 samples were identical to the reference gene sequence without any variations detected. While the other 46 samples showed different types of variations in the ZEB2 gene. The number of variants present in the samples was different from one sample to another, 23 out of 46 samples showed single variation in their sequences, seven samples showed double variations, the remaining 16 samples showed multiple variations. According to the effect of the ZEB2 gene variations on TNF-α, TGF-β, and IFN-γ serum levels, the results revealed a significant difference between Identical (ID) group (42.5%) (Identical: no variations were detected), single variation group (28.75%) (One variation was detected), double variation group (8.75%) (Two variations were detected) and multiple variation group (20%) (More than two variations were detected) in both IFN-γ and TGF-β with a p-value of (0.0518) and (0.0368), respectively. While the results displayed non-significant differences between the same groups in the level of TNF-α (p value= 0.703) (Table 3).
The current study also showed an increase in the serum level of TGF-β with an increase in the number of variations in the ZEB2 gene (double and multiple variation groups). These results were similar to the ones which were reported by Li et al., (2017) [14] who found that the mutation and deletion in the ZEB2 gene can cause overexpression in TGF-β and demonstrated that the ZEB2 gene inactivation within the HSPCs (hematopoietic stem/progenitor cells) correlates with modifications in expression of genes involved in cytokine receptor signaling. These alterations might affect the differentiation of numerous hematopoietic lineages by modulating signal transmission. In this study, a positive relationship between TNF-α and TGF- β was found in volunteers who had multiple genetic variants, as they had a higher rate than volunteers with identical genetic sequences.
While in the case of IFN-γ, the relationship was inverse, the results showed that identical genetic volunteers had a higher mean value than volunteers who had multiple genetic mutations.
Figures & Tables

Discussion![]()
The variations in the ZEB2 gene are likely to have a negative impact on the development of organs and systems. Severe syndromic abnormalities are caused by mutations in ZEB encoding genes, and evidence is growing that these factors contribute to the growth of malignant tumors [15]. The ZEB2 gene codes for a protein that aids in the creation of various organs and tissues prior to birth. This protein is a transcription factor (TF), which means it attaches to certain sections of DNA and aids in the control of gene activity [4]. The epithelial–mesenchymal transition, which is critical in embryonic development, wound healing, and cancer progression, is predominantly related with this TF (ZEB2) [16].
The ZEB2 protein has been involved in chemical signaling pathways that govern prenatal development and growth. The different activities of the ZEB2 gene in nervous system evolution are characterized by a variety of severe neurological disorders, such as those seen in Mowat-Wilson syndrome (MWS), which is caused by a mutation in the ZEB2 gene [17].
The ZEB2 gene is also considered as a cancer biomarker, in which the relationship of the ZEB2 gene to several types of cancers has well studied by numerous researchers. The outcome of these research projects suggests presence of a relationship between the expression level of this transcription factor (ZEB2) and the grade of malignancy, fast cell proliferation, and deprived patient survival in various tumors. Through transcriptional regulation of epithelial-to-mesenchymal transition, the ZEB2 gene plays a key role in solid cancer metastasis, allowing cancer cells to infiltrate and spread . Dominguez et al., (2015) [18] proved that the deletion, insertion or alteration in the nitrogenous bases that make up the ZEB2 gene have caused a decrease in the immune system.
Zou et al., [19] studied 333 patients, and discovered four ZEB2 gene mutations (two recurrent and two novel) that were linked to MWS symptom, implying that not all MWS ZEB2 alleles fully lost their functions. Hu et al., (2020) [20], in a case report, investigated a 14 years old girl with MWS, within the third exon of the ZEB2 gene they detected a novel nonsense mutation resulted in a termination codon at position 84 amino acid. They suggested that this amino acid alteration was probably the pathogenic mutation that caused MWS in the proband. The MWS protein carries novel heterozygous ZEB2 mutations, and most of these produce premature stop codon (85%) or deletion encompassing the gene (15%). Various mutations in the MWS proteins result in either full ZEB2 protein ablation or nonfunctional ZEB2 protein synthesis [21].
Dendritic cells called plasmacytoid dendritic cells (pDCs) are key producers of type 1 interferons and play a role in controlling immunological responses to viruses [22]. In 2012, the ImmGen consortium first reported that ZEB2 was expressed by pDCs [23]. The loss of function and pathogenic variations of the ZEB2 gene might affect the function of these types of cells and the IFN production is one of these functions.
Genetic variants in a gene's coding sequences (exons) have often been given priority because they are easier to read, despite the fact that it has long been evident that coding sequence variants alone are insufficient for mapping complicated illnesses. Variants in the intervening sequences (introns) or untranslated regions (UTRs), while not affecting the predicted protein sequence, may play a key role in gene expression regulation.
The RNA sequence immediately upstream of the coding RNA is known as the 5′UTR region. It is rarely translated, though there are notable cases in which a portion of the 5′ UTR becomes translated [24]. Its length varies from a few nucleotides (nt) to several thousands in eukaryotes, with an average of about 200 nt in humans [25]. The Kozak consensus sequence (ACCAUGG), which contains the translation initiation codon, should be present in the 5′ UTR. Furthermore, secondary structures such as hairpin loops, which are frequently found inside the 5′UTR, may play a role in translation regulation. By influencing RNA transcription, translation and stability, genetic variations impacting these regulatory elements can have a significant impact on overall protein output [25-27].
Finally, we believed that this study finding, especially the nine novel detected variants in exon 1 and 5`UTR regions of the ZEB2 gene, might obtain importance in the field of the ZEB2 gene studies due to the presence of these variants in the important coding and regulatory regions that regulate expression of the ZEB2 gene. Furthermore, the two variants, 144520053G/A and 144520162T/A, among the nine novel variants might be the most important variants because both showed significant effect on some study parameters. These variants need to be further studied in the future to identify precisely their role in regulating the expression of TGF-α, TNF-β and IFN-γ that are involved in immune cell differentiation.
Acknowledgement
The authors would like to express special thanks and gratitude to the staff of Clinical Laboratories department at the College of Applied Medical Sciences/ University of Kerbala, for their support.
Author Contributions
The author declares that there is no conflict of interest regarding the publication of this paper.
References
- Vandewalle C, Comijn J, De Craene B, Vermassen P, Bruyneel E, et al. SIP1/ZEB2 induces EMT by repressing genes of different epithelial cell–cell junctions. Nucleic acids research, (2005); 33(20): 6566-6578.
- Wiles ET, Bell R, Thomas D, Beckerle M, Lessnick SL. ZEB2 represses the epithelial phenotype and facilitates metastasis in Ewing sarcoma. Genes & cancer, (2013); 4(11-12): 486-500.
- Wang J, Farkas C, Benyoucef A, Carmichael C, Haigh K, et al. Interplay between the EMT transcription factors ZEB1 and ZEB2 regulates hematopoietic stem and progenitor cell differentiation and hematopoietic lineage fidelity. PLoS biology, (2021); 19(9): e3001394.
- Scott CL, Omilusik KD. ZEBs: novel players in immune cell development and function. Trends in immunology, (2019); 40(5): 431-446.
- Peinado H, Olmeda D, Cano A. Snail, Zeb and bHLH factors in tumour progression: an alliance against the epithelial phenotype? Nature reviews cancer, (2007); 7(6): 415-428.
- Wang J, Lee S, Teh CE-Y, Bunting K, Ma L, et al. The transcription repressor, ZEB1, cooperates with CtBP2 and HDAC1 to suppress IL-2 gene activation in T cells. International immunology, (2009); 21(3): 227-235.
- Sánchez-Tilló E, Siles L, De Barrios O, Cuatrecasas M, Vaquero EC, et al. Expanding roles of ZEB factors in tumorigenesis and tumor progression. American journal of cancer research, (2011); 1(7): 897.
- Stankiewicz TR, Gray JJ, Winter AN, Linseman DA. C-terminal binding proteins: central players in development and disease. Biomolecular concepts, (2014); 5(6): 489-511.
- Deverman BE, Patterson PH. Cytokines and CNS development. Neuron, (2009); 64(1): 61-78.
- Gulati K, Guhathakurta S, Joshi J, Rai N, Ray A. Cytokines and their role in health and disease: a brief overview. MOJ Immunology, (2016); 4(2): 1-9.
- Zidek Z, Anzenbacher P, Kmoníčková E. Current status and challenges of cytokine pharmacology. British journal of pharmacology, (2009); 157(3): 342-361.
- Tato CM, Cua DJ. SnapShot: cytokines II. Cell, (2008); 132(3): 500. e501-500. e502.
- Hamilton SE, Jameson SC (2015) Effective effector generation of CD8+ T cells and NK cells: a need for T-bet and ZEB-too. The Rockefeller University Press.
- Li J, Riedt T, Goossens S, Carrillo García C, Szczepanski S, et al. The EMT transcription factor ZEB2 controls adult murine hematopoietic differentiation by regulating cytokine signaling. Blood, The Journal of the American Society of Hematology, (2017); 129(4): 460-472.
- Vandewalle C, Van Roy F, Berx G. The role of the ZEB family of transcription factors in development and disease. Cellular and molecular life sciences, (2009); 66(5): 773-787.
- Craene BD, Berx G. Regulatory networks defining EMT during cancer initiation and progression. Nature Reviews Cancer, (2013); 13(2): 97-110.
- Becker C, Fantini MC, Schramm C, Lehr HA, Wirtz S, et al. TGF-β suppresses tumor progression in colon cancer by inhibition of IL-6 trans-signaling. Immunity, (2004); 21(4): 491-501.
- Dominguez CX, Amezquita RA, Guan T, Marshall HD, Joshi NS, et al. The transcription factors ZEB2 and T-bet cooperate to program cytotoxic T cell terminal differentiation in response to LCMV viral infection. Journal of Experimental Medicine, (2015); 212(12): 2041-2056.
- Zou D, Wang L, Wen F, Xiao H, Duan J, et al. Genotype‑phenotype analysis in Mowat‑Wilson syndrome associated with two novel and two recurrent ZEB2 variants. Experimental and therapeutic medicine, (2020); 20(6): 1-1.
- Hu Y, Peng Q, Ma K, Li S, Rao C, et al. A novel nonsense mutation of ZEB2 gene in a Chinese patient with Mowat‐Wilson syndrome. Journal of Clinical Laboratory Analysis, (2020); 34(9): e23413.
- Ghoumid J, Drevillon L, Alavi-Naini SM, Bondurand N, Rio M, et al. ZEB2 zinc-finger missense mutations lead to hypomorphic alleles and a mild Mowat–Wilson syndrome. Human molecular genetics, (2013); 22(13): 2652-2661.
- McKenna K, Beignon A-S, Bhardwaj N. Plasmacytoid dendritic cells: linking innate and adaptive immunity. Journal of virology, (2005); 79(1): 17-27.
- Miller JC, Brown BD, Shay T, Gautier EL, Jojic V, et al. Deciphering the transcriptional network of the dendritic cell lineage. Nature immunology, (2012); 13(9): 888-899.
- Cenik C, Derti A, Mellor JC, Berriz GF, Roth FP. Genome-wide functional analysis of human 5'untranslated region introns. Genome biology, (2010); 11(3): 1-17.
- Mignone F, Gissi C, Liuni S, Pesole G. Untranslated regions of mRNAs. Genome biology, (2002); 3(3): 1-10.
- De Angioletti M, Lacerra G, Sabato V, Carestia C. β+ 45 G→ C: a novel silent β‐thalassaemia mutation, the first in the Kozak sequence. British journal of haematology, (2004); 124(2): 224-231.
- Pichon X, A Wilson L, Stoneley M, Bastide A, A King H, et al. RNA binding protein/RNA element interactions and the control of translation. Current Protein and Peptide Science, (2012); 13(4): 294-304.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0