

Full Length Research Article
Heritability of physiological traits of barley (Hordeum Vulgare L.) under salinity stress
Siavash Solhi Andarab1, Varahram Rashidi1*, Hossein Shahbazi2, Farhad Farahvash1, Alireza Ahmadzadeh1
Adv. life sci., vol. 9, no. 2, pp. 207-213, July 2022
*– Corresponding Author: Varahram Rashidi (Email: rashidi.varahram@gmail.com)
Authors' Affiliations
2. Department of agronomy and plant breeding, Ardabil branch, Islamic Azad University, Ardabil – Iran
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: In this paper, to determine the genetic parameters and heritability of physiological traits related to salinity stress in barley (Hordeum Vulgare L.), a 7×7 half diallel cross was conducted at Islamic Azad University, Ardabil, during 2016-2017.
Methods: F1 seeds are used in this experiment. These seeds have been grown under non-stress conditions, 8 and 12 dS/ m salinity stress levels, in a greenhouse alongside the parent plants. After exposure of plants to salinity, Leaf chlorophyll content, Chlorophyll Fluorescence and Stomatal Conductance were measured.
Results: Narrow sense heritability of chlorophyll content ranged from 0.29 to 0.40. These values for chlorophyll fluorescence ranged from 0.16 to 0.24 and for stomatal conductance ranged from 0.26 to 0.54. Broad sense heritability was high among the evaluated traits and ranged from 0.70 to 0.87. The degree of average dominance was higher than one in all of traits.
Conclusion: This observation indicated some degree of over dominance in control of the traits. Although dominant alleles were not observed in leaf chlorophyll content and chlorophyll fluorescence, yet they were favorable in stomatal conductance.
Keywords: Salinity tension; Diallel; Heritability; Physiological traits
Introduction![]()
Plants are constantly exposing to several stresses in their growth environment that somehow limit their development and survival. Salinity stress, which effects the biochemical, physiological and biosynthetic processes during crop production [1, 2], is one of the most important abiotic stresses [1, 3]. Nutritional imbalance, low osmotic potential in soil solution, and the effect of specific ions on salinity stress are some of the destructive effects of plant growth which are caused by salinity [4]. Salinity affects some of the main metabolic processes through two dehydration osmotic processes and toxicity of ion accumulation in plant cells [5]. Although barley is among the most tolerant crops to salinity, nevertheless, salinity has restricted the production of this crop around the world. This issue is mostly observed in arid and semi-arid regions [6]. Understanding the mechanisms of salt tolerance, is the key to improve salt tolerance of barley [7]. Most breeding programs, especially improvement of tolerance to environmental stresses, have been based on empirical selection of yield, which is not very efficient due to low heritability and genotype × environment interactions. Therefore, indirect selection, based on physiological traits has been proposed as a complementary to the selection of yield [8]. By combining information on the physiological basis of yield limits and using new selection tools, the genetic improvement of stress tolerance can be increased. From the perspective of plant breeding, each secondary physiological trait should have sufficient genetic diversity, a strong genetic correlation with yield and higher heritability than yield [9]. In addition, the evaluation of these traits should be quick, easy and cheap [10]. Among the physiological traits maximum quantum yield of photosystem II in dark adapted state (Fv/Fm) can be regarded as important indicator in the evaluation of salinity stress [11, 12]. Since salinity affects the photosynthesis process [13], it is possible to use the level of chlorophyll accumulation as a criterion for salinity tolerance. Since the decrease of chlorophyll concentration signifies toxicity in tissues due to accumulation of ions, it can act as an indicator of salinity tolerance [14]. Reduction of chlorophyll content under salinity was greater in salt sensitive cultivars comparing tolerant cultivars [15-17]. The results of Rahnama et al., [18] for wheat and other cereals indicate that in presence of salinity, the change in stomatal conductance can be used as a means of screening for salinity stress tolerance. Knowledge of the heritability of traits related to salinity tolerance is essential in designing selection methods to create superior genotypes. The appropriate breeding method requires adequate information on genetic control of traits. Hence, the main goal of this study is evaluating the gene action and inheritance of physiological traits related to salinity stress in barley using diallel cross.
Methods![]()
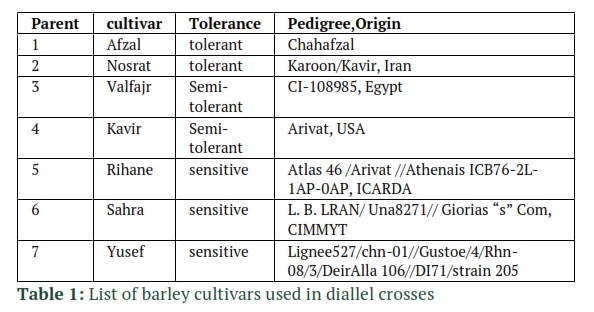
To determine the heritability of physiological traits under salinity stress a 7 × 7 half diallel cross was performed among 7 barley cultivars (table1) in agricultural research station of Islamic Azad University, Ardabil, Iran in 2016. F1 seeds are used in this experiment which was conducted under non-stress conditions, 8 and 12 dS/m salinity stress levels, in a greenhouse. The seeds were kept in the same environment as their parent plants. Five seeds were grown in plastic pots of size 25 × 30 cm which were filled with soil consisted of a mixture of sand, sterilized compost, and garden soil (1:1:1, v/v). Average of minimum and maximum temperature of greenhouse was 12 and 30°C. Relative humidity ranged from 62% to 75%. For uniformity of seed emergence especially in sensitive cultivars, application of salinity was started after plant growth and reaching to the 3rd Leaf (Z20 Zadoks scale). Saline water was gradually added to the pots with the desired salinity. The salinity was applied gradually in two weeks. We measured the electric conductivity of the drained water from pots to control the salinity level. Subsequent irrigations were done using tap water after reaching the desired EC. Also, to prevent salt washing and return the extra water to the pot, saucers were used. After the plants were exposed to salinity and flowered, leaf chlorophyll content, Chlorophyll fluorescence and stomatal conductance were measured.
Measurement of traits
Chlorophyll fluorescence: maximum quantum yield of photosystem II in dark adapted state (Fv/Fm) was measured in flag leaves of 3 plants per pot using OS30P Fluorometer, made by OPTI-Science.
Stomatal conductance: Decagon SC-1 leaf porometer was used to measure the Gs value of three plants in flag leaves. Measurements were done between 10 and 12 A.M. and the stomatal conductance was calculated in (cm/scm/mm).
Leaf chlorophyll content: CCM200, made by OPTI-Science, was used to measure the chlorophyll content of flag leaves of three plants per pot.
Statistical analysis
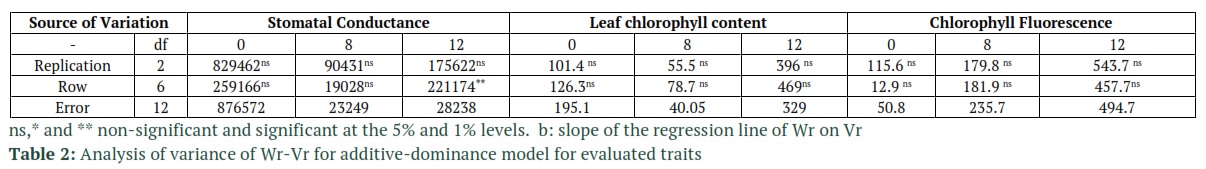
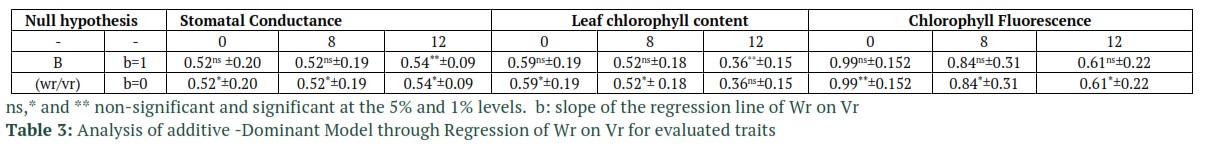
The analysis of the variance of the diallel was performed out by the Hayman’s method (1954b), which Walters and Morton later developed for half-diallel in [19]. Genetic components such as covariance between the additive and dominant effects (F), corrected dominance variance (H2), uncorrected dominance variance (H1), the dominance effects (h2) which is the algebraic sum of all heterozygote gene loci in all crosses, and additive genetic variance (D) were calculated using the method provided by Singh [20]. Significance of genetic components was evaluated using t-test. Average degree of dominance, narrow and broad sense heritability were calculated based on the method in [21]. The goodness of fit of the proposed model in [19] which is also the model that is used in this study, was tested using the analysis of variance on the difference of array of covariance between parent and offspring (Wr) and the array of variance (Vr). Additionally, to further check the closeness of the observed data to the expected data in the additive-dominant model, linear regression was used on Wr and Vr arrays (H0: b = 1 vs. H1: b ≠ 1). DIAL98 [22] software was used to calculate the diallel analysis of variance. Also, Microsoft’s Excel was used to estimate genetic components.
Also, the combining ability was analyzed based on [23] using the SAS 9.2 Software. This analysis was performed based on Model I and Method II of [23]. The relative importance of non-additive and additive gene effects was assessed using the Baker’s variance ratio (2MSgca/(2MSgca+MSsca) which was computed according to Baker (1978).
Results
![]()
Hayman’s graphical method
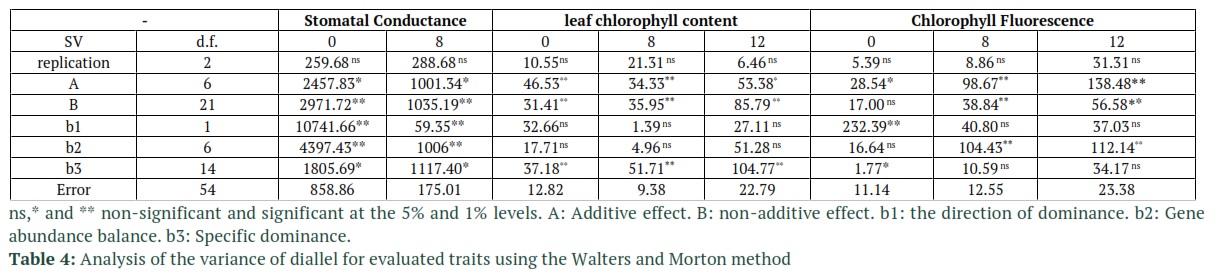
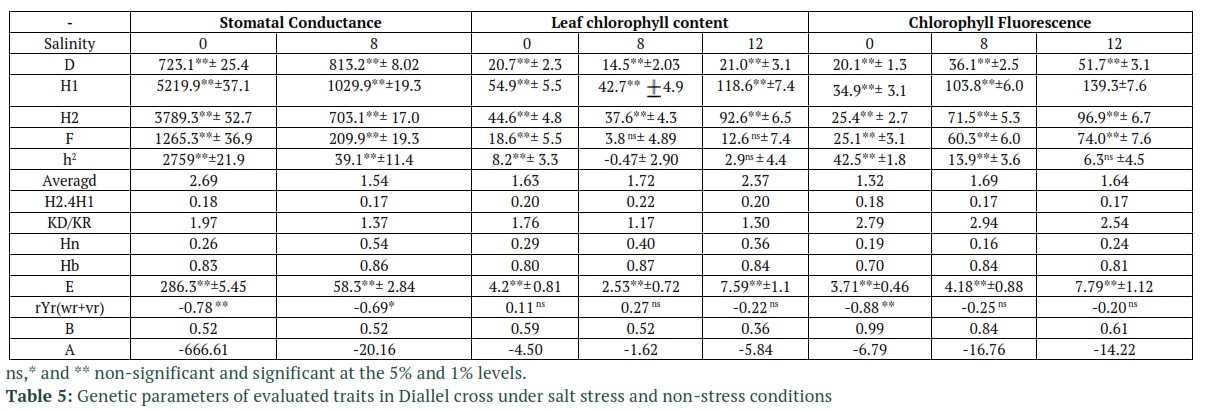
Tables 2 and 3 report the goodness of fit test results for the additive-dominant model. The accuracy of the tested model, based on the non-significant mean squares value of Wr-Vr for the treatment group, for all of the traits (except for stomatal conductance under 12 ds/m salinity) is shown to be adequate. The reported results from Dashti et al., [24] are also in compliance with the results from Tables 2 and 3. The slope of linear regression in stomatal conductance and leaf chlorophyll content (under 12 ds/m salinity) were also lower than 1 and the model was not fitted (Table3). Based on the significance of component A in Table 4 which represents additive variance, additive effects were detected in the control group for all of the traits. Table 5 also confirms the results of Table 4, especially for component A’s statistical significance in Table 4 which complies with D component’s significance (additive effects) in Table 5. The statistical significance of dominant genetic effects for all the tested traits, except for chlorophyll fluorescent (non-stress condition), shows how important these effects are in the tested traits. Based on the results in Table 4, the F1’s mean variations which are measured through component “b1” from the mid-parent values had statistical significance for stomatal conductance and chlorophyll fluorescent under non-stress conditions. For the most part, the dominance is in one direction due to the fact that component “b1” is statistically significant. Hence, it measures the average hybrid vigor [20]. Another results from Table 5 that confirms the effects of dominance on the traits, is the significance of H1 and H2 components which represent the variances of non-corrected and corrected dominance, respectively. Component “b2” was also statistically significant for stomatal conductance and chlorophyll fluorescent. This significance in component b2 shows that the difference between the Fls and their mid-parent values, which is referred to as the average dominance deviation, highly variates in the F1 arrays. These results also an indication of the fact that not all parents perform equally and some of them were significantly superior to other performance-wise. In table 5, H2/4H1 was calculated to estimate the proportions of positive and negative genes. The values of H2/4H1 which were below 0.25 for all the traits, indicated that positive and negative alleles were asymmetrically distributed in parents. The fact that H2 is lower than H1 in these traits also proves this conclusion. The variance of Specific Combining Ability (SCA) which is represented by “b3” is also reported to be significant in stomatal conductance and leaf chlorophyll content at all of stress levels and in chlorophyll fluorescence under non-stress condition. This was due to the significance of the estimate of component F which was reported for all cases except for leaf chlorophyll content under salinity. Dominant alleles in parents are expected to be more common in the case of positive values of b3. The KD/KR ratio in Table 5 is reported to be higher than 1 which shows the higher proportion of the dominant alleles compared to the proportion of the recessive alleles in the parents. These results are in accordance the values of F which are also reported in Table 5. In this table, the regression line has a negative intercept value which indicates over-dominance. Over-dominance is also observed in averagd parameter in Table 5 which determines the degree of average dominance. The fraction of phenotypic variance of the traits which is also known as narrow sense heritability
and is represented by Hn in Table 5 is relatively low to moderate, considering that its value is greater in stomatal conductance and leaf chlorophyll content than chlorophyll Fluorescence. Although the dominance of alleles was not observed in the case of leaf chlorophyll content and chlorophyll fluorescence, however, the negative values of rYr(Wr+Vr) in Table 5 which represents the correlation coefficients between the order of dominance and the parental averages in stomatal conductance showed some signs of this dominance.
Griffing’s analysis
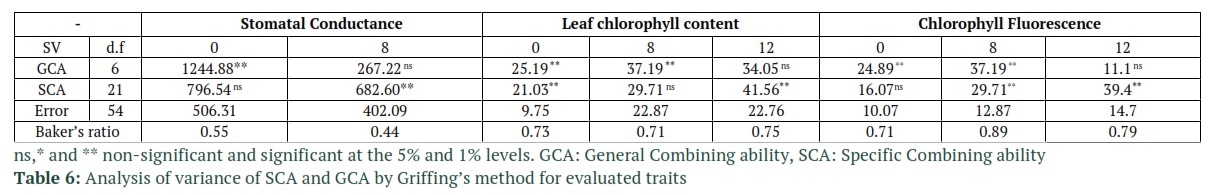
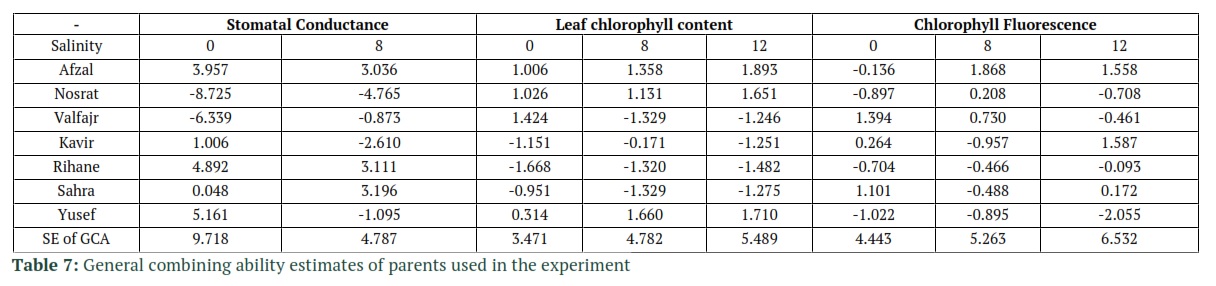
The significance of General Combining Ability (GCA) mean squares in stomatal conductance under non-stress, leaf chlorophyll content and chlorophyll fluorescence under non-stress and 8 ds/m salinity was shown by the Griffing’s combining ability analysis. Also, the values of SCA mean squares in stomatal conductance and chlorophyll fluorescence were statistically significant, yet under salinity stress. However, in leaf chlorophyll content it was significant under 12 ds/m salinity and non-stressed conditions. Based on these results it can be concluded that the importance of dominance effects had been increased with the increase of salinity (Table 6). Non-additive effects in the case of Chlorophyll fluorescence and chlorophyll content are less important than the additive effects base on the values of the Baker’s ratio in Table 6. It was concluded from the GCA effects results in Table 7 that under salinity, the tolerant cultivar Afzal, and sensitive cultivars, Rihane and Sahra had favorable alleles for stomatal conductance. High stomatal conductance in those sensitive cultivars indicates that the sensitivity of Rihane and Sahra to salinity can be attributed to ion toxicity rather than osmotic effect of salinity, since they could acquire adequate water and open their stomata under salinity. Afzal and Nosrat the tolerant cultivars along with sensitive cultivar Yusef had favorable alleles for leaf chlorophyll content. Under 8 ds/m salinity, tolerant cultivars (Afzal, Nosrat and Valfajr) and under 12 ds/m tolerant cultivars such as Afzal and Kavir had favorable alleles for Chlorophyll Fluorescence. To grow off-springs that have improved attributes and increase the crop yield, it is important to choose proper parents for crossing. So, identification of parental lines with good general combining ability could be useful for varietal improvement.
Figures & Tables
Discussion![]()
These variations mean that alleles are distributed asymmetrically among the parents [25]. Additive variance is not measured unambiguously through component “a” due to the fact that b2 is significant for stomatal conductance and chlorophyll fluorescent. However, this component would have non-additive variance as well [20]. The residual of dominance effects that are not estimated by b1 and b2 are considered in b3 which combines interaction effects such as Dominance × Dominance, Additive× Additive, and Additive × Dominance [26]. Also, asymmetrical distribution of recessive alleles as well as dominant ones was experienced in the parents. Over-dominance was also used in [27] to control chlorophyll a and b. On the other hand, the value of Hb which is the broad sense heritability of the traits is high. The dominance effects can also be deducted in the differences between the Hn and Hb values.
This paper focused on reporting and analyzing the results of a 7×7 half diallel cross which was conducted at Islamic Azad University, Ardabil, during 2016-2017. In most cases, inheriting the evaluated traits could be explained with a simple genetic model. This was concluded using the Hayman's analysis and its assumption which was fulfilled. The analysis of the generation means in an experiment revealed the contribution of epistasis and dominance gene interactions to control of K+, Na+ and K+/Na+, however, chlorophyll content was in the control of additive and dominant effects [28]. The broad sense heritability estimated values, which were reported to be high, for the tested traits indicated the larger effect genotype on these trains than the effect of the growing environment. These results indicated that salt tolerance might be improved by better selection. The study of genetic components of traits is one of the basic factors for the success of breeding programs. Hence, selection of a proper breeding technique is heavily impacted by the comprehensiveness and accuracy of the information on the genetic parameters that control the physiological traits. Additive and nonadditive gene effects in wheat has already been shown to control its salinity tolerance [20]. One of the practical methods to determine salt injury has shown to be evaluating the physiological traits to assess the plant material, despite the complexity of the expression of salt tolerance [29]. Over-dominance was experienced due to the fact that the average dominance had a degree value over 1, which also is a sign of the traits being controlled heavily by the dominance effects.
For some of the cases, delayed stage plant selection was more effective for lower estimated values of heritability and the estimated results of the dominance gene effects. However, early generation selection improved stomatal conductance based on the estimations for heritability, for which high values were estimated, and additive gene effects. Due to the importance of narrow sense heritability in selection of a proper breeding technique and based on the experimental results, the stomatal conductance can be considered as a proper feature evaluating the salinity tolerance in barley. Some of the other traits demonstrate lower values for narrow sense heritability. Hence, these traits can be used as selection criteria for salinity tolerance in advanced generations of barley breeding programs. GCA variations among the parents was indicated by the reported high values of the mean squares of GCAs of the evaluated traits. These results suggested that selection over a segregating population can lead to genetic gain. Based on the results of the combining ability analysis, SCA includes two types of epistatic gene actions, namely dominance × dominance and additive × dominance, while a specific part of variance is formed by additive × additive because of GCA [25]. The progeny performance of self-fertilizing species can be properly predicted based on the estimations of gca effects of individual lines. This is due to the fact that additive × additive epistasis and additive gene action are, based on [30], exploitable in homozygous genotypes. Based on physiological studies such as [31], a combination of sub-traits determined the overall trait of salt tolerance. Additive effects, dominance, and heterosis are influential in determining these sub-traits.
Although in breeding programs selection criteria such as physiological traits are important, yet, it is better that selection for large dominance effects not be performed before some inbreeding is carried out However, in cross pollinated crops, it is potentially possible to exploit these dominance effects during the process of developing F1 hybrids.
Author Contributions
Dr. Rashidi and Shahbazi designed the experiment and supervised the research. Dr. Farahvash, and Ahmadzadeh served as consultants and participated in the analysis. Dr. Solhi Andarab conducted the experiment.
The authors certify that they have no conflict of interest to declare in the subject matter or materials discussed in this manuscript.
References
- Assefa Woldemariam G. Efforts, successes and challenges of green feed production in Ethiopia. Online Journal of Animal and Feed Research, (2021); 11: 13-17.
- Karasakal A, Khayatnezhad M, Gholamin R. The Effect of Saline, Drought, and Presowing Salt Stress on Nitrate Reductase Activity in Varieties of Eleusine coracana (Gaertn). Bioscience Biotechnology Research Communications, (2020); 13(4): 2087-2091.
- Naheed R, Arfan M, Farhat F, Ijaz S, Khalid H. Acclimatization of drought tolerance with Somaclonal variants of sugarcane (Saccharum officinarum L.). Advancements in Life Sciences, (2020); 8(1): 57-62.
- Heidari A, Toorchi M, Bandehagh A, Shakiba MR. Effect of NaCl stress on growth, water relations, organic and inorganic osmolytes accumulation in sunflower (Helianthus a nnuus L.) lines. Universal journal of environmental research and technology, (2011); 1351-362.
- Parida AK, Das AB. Salt tolerance and salinity effects on plants: a review. Ecotoxicol Environ Safety, (2005); 60: 324-349.
- Kalaji M, Govindjee H, Bosa K, Koscielniak J, Zuk-Golaszewska K. Effects of salt stress on photosystem II efficiency and CO2 assimilation of two Syrian barley landraces. Environmental and Experimental Botany, (2015); 73: 64-72.
- Colmer TD, Munns R, Flowers TJ. Improving salt tolerance of wheat and barley; future prospects. Australian Journal of Experimental Agriculture, (2005); 45: 1425-1443.
- Reynolds MP, Singh RP, Ibrahim A, Ageeb OAA, Larque-Saavedra A, et al. Evaluating physiological traits to compliment empirical selection for wheat in warm environments. Euphytica, (1998); 100: 85-94.
- Jackson P, Robertson M, Cooper M, Hammer G. The role of physiological understanding in plant breeding; from a breeding perspective. Field Crops Research, (1996); 49: 11-39.
- Araus JL, Casadesus J, Bort J (2001). Recent tools for the screening of physiological traits determining yield. In: Reynolds MP, Ortiz-Monasterio JL, McNab A, editors. Application of physiology in wheat breeding. Mexico: D.F. CIMMYT. pp. 59-77.
- Misra AN, Misra M, Singh R (2012). Chlorophyll Fluorescence in Plant Biology. In book: Biophysics, Chapter: 7. In: Misra AN, editor. New York: InTech. pp. 171-192.
- Zhao X, Chang AY, Toh-E A, Arvan P. A role for Lte1p (a low temperature essential protein involved in mitosis) in proprotein processing in the yeast secretory pathway. Journal of Biological Chemistry, (2007); 282(3): 1670-1678.
- Udovenko GV (1997) Salt Tolerance of Cultivated Plants. In: Brezhnez DD, editor. Leningrad: Kolos.
- Khosravinejad F, Heydari R, Farboodnia T. Effects of Salinity on Photosynthetic Pigments, Respiration, and Water Content in Two Barley Varieties. Pakistan Journal of Biological Sciences, (2008); 11(20): 2438-2442.
- Ashraf M, Karim F, Rasul E. Interactive effects of gibberellic acid (GA3) and salt stress on growth, ion accumulation and photosynthetic capacity of two-spring wheat (Triticum aestivum L.) cultivars differing in Salt tolerance. Plant Growth Regulation, (2002); 36: 49-59.
- Lee G, Carrow RN, Duncan RR. Photosynthetic responses to salinity stress of halophytic seashore paspalum ecotypes. Plant Science, (2009); 166: 1417-1425.
- Qiu N, Lu C. Enhanced tolerance of photosynthesis against high temperature damage in salt-adapted halophyte Atriplex centralasiatica plants. Plant Cell Environ, (2003); 26: 1137-1145.
- Rahnama A, James RA, Poustini K, Munns R. Stomatal conductance as a screen for osmotic stress tolerance in durum wheat growing in saline soil. Functional Plant Biology, (2009); 37(3): 255-263.
- Walter DE, Morton R. On the analysis of variance of half diallel table. Biometerics, (1978); 3491-94.
- Singh M, Singh RK. A comparison of different methods of half-diallel analysis. Theoretical and Applied Genetics, (1984); 67: 323-326.
- Mather K, Jinks JL Biometrical Genetics. Chapter: Book Name. 1971 of publication; Chapman & Hall.
- Ukai Y. A microcomputer program DIALL for diallel analysis of quantitative characters. Japanese Journal of Breeding, (1989); 39: 107-109.
- Griffing B. Concept of general and specific combining ability in relation to diallel crossing systems. Australian Journal of Biological Science, (1956); 9: 463-493.
- Dashti H, Naghavi MR, Tajabadipour A. Genetic Analysis of Salinity Tolerance in a Bread Wheat Cross. Journal of Agricultural Science and Technology, (2010); 12: 347-356.
- Hayman BI. The theory and analysis of diallel crosses. Genetics, (1954); 39: 789-809.
- Choudhary S, Vadez V, Hash CT, Kishor PK. Pearl Millet Mapping Population Parents: Performance and Selection Under Salt Stress Across Environments Varying in Evaporative Demand. Proceedings of the National Academy of Sciences, India Section B: Biological Sciences, (2019); 89(1): 201-211.
- Metwali EMR, Abd-El-Haleem SHM, EL-Saeid R, Kadasa NMS. An Investigation of Gene Action on Different Traits of Barley (Hordeum Vulgare L.)Using Partial Diallel Crosses System. Life Science Journal, (2014); 11(1): 64-71.
- Farshadfar E, AghaieSarbarzah M, Sharifi M, Yaghotipour A. Assessment of Salt Tolerance Inheritance in Barley via generation Mean Analysis. Journal of Biological Sciences, (2008); 8(2): 461-465.
- Noble CL, Halloran GM, West DW. Identification and selection for salt tolerance in lucerne (Medicago sative L.). Aust. Journal of Agricultural Research, (1984); 35239-252.
- Baker R. Issues in diallel analysis. Crop Science, (1978); 18: 533-536.
- Flowers TJ. Improving crop salt tolerance. Journal of Experimental Botany, (2004); 55: 307-319.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0