

Full Length Research Article
Anatomical transition of trilaminar cerebellar cortex between reptiles and Aves
Naheed Khan1,2, Khalida Perveen1,2, Mushtaq Hussain2*, Rabia Qadeer Malik3, Shaheen Sharafat4
Adv. life sci., vol. 10, no. 2, pp. 259-264, June 2023
*– Corresponding Author: Mushtaq Hussain (mushtaq.hussain@duhs.edu.pk)
Authors' Affiliations
2. Bioinformatics and Molecular Medicine Research Group, Dow Research Institute of Biotechnology and Biomedical Sciences, Dow College of Biotechnology, Dow University of Health Sciences, Karachi – Pakistan
3. Department of Anatomy, University of Kent – UK
4. Department of Pathology, Liaquat National Hospital and Medical College, Karachi – Pakistan
[Date Received: 24/01/2023; Date Revised: 18/06/2023; Date Published Online: 30/06/2023; Date Updated:11/09/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Motor coordination in vertebrates is primarily regulated by cerebellum. Divergence of Aves from reptilian ancestors results in noticeable improvement in the motor coordination. This study aims to explore anatomical innovations in the cerebellar cortex during the course of evolution of reptiles and Aves.
Methods: Three representative species each from reptilian and avian lineages were selected to represent both vertebrate classes. Complete brain was dissected out from the cranial cavity of each specimen after radiological assessment of its extent. After gross examination, the brains were subjected to detailed histological investigation using conventional and special stains. Micrometry of layer and cellular architecture of cerebellar cortex were undertaken digitally using ImageJ and statistically compared using GraphPad Prism.
Results: Grossly, significant increase (p<0.0001) in brain mass, brain volume and cerebellar volume was observed in Aves compared to reptiles. Histomorphometric analyses of granular and molecular layers of cerebellum showed statistically significant decrease (p<0.0001) in the thickness of avian representatives compared to reptilian counterparts. Similarly significant decrease (p<0.0001) in the Inter-Purkinje neuronal distance was observed in Aves compared to reptiles. Conversely, increased cellular and neuronal count (p=0.0332 to <0.0001) was observed in all three layers of avian cerebellum in comparison to reptiles. This suggests increased cellular packaging and/or density in the avian cerebellum compared to reptiles.
Conclusion: In summary, significant increase in the cellular density and differentiation in the cerebellum of avian representatives may provide anatomical basis of increased motor coordination in Aves compared to reptiles.
Keywords: Cerebellum; Evolution; Granular Layer; Molecular Layer; Purkinje Layer
Introduction![]()
vertebrate species, involved in coordination and motor learning [1]. The characteristic organization of afferent and efferent projections in cerebellum, suggests its role in the integration of motor commands and sensory information required for the execution of coordinated movements [2]. More recently, non-motor functional domains such as cognition, emotions, language, working memory [1], and the coordination of errors in thought [3,4] are also linked with the biological roles of cerebellum.
Characteristic anatomy of cerebellum, with major subdivisions of corpus cerebellum and vestibulo-cerebellum, first appeared in gnathostomes and chondrichthyes [5,6,7] which by and large remained conserved in the vertebrate lineages during the course of evolution. Nevertheless, histological architecture of cerebellum is conserved with three cellular layers in all vertebrate lineages. This trilaminar form widely referred to as cerebellar cortex, is constituted by three layers: the outer molecular layer, an intermediate row of Purkinje neurons and an inner granular layer [8,9,10]. Typically, the basic organization of cerebellar cortex neural circuitry is also well conserved in vertebrates, which mainly comprises of two types of input systems: mossy and climbing fibers in addition to the granule cells, modulatory interneurons including stellate, golgi and basket cells which regulate the discharge of single output Purkinje neuron [9,11,12].
Reptiles and Aves are the two major taxonomic and/or evolutionary lineages of vertebrates, where reptiles are the antecedent representative that not only set the origin of mammals around 319 million years ago (MYA) but lately (261 MYA) separate sub-branch of reptiles lead to the origin and subsequent divergence of Aves [13,14]. Reptiles and Aves are significantly different from each other in relation to the motor activity, where by and large the later are considerably more agile and many have flight and supremely evolved predatory capability compared to former. This represents significant fundamental lead to the cerebellum to coordinate extensively evolved motor activities in Aves. Studies conducted in avian and reptilian species showed that despite much histological conservation, the gross anatomy of cerebellum varies greatly. Reptiles generally have unfolded cerebella, whereas Aves have variably folded cerebellar architecture [15,16]. Some microanatomical differences are also evident between two taxonomic groups. For example, few representatives of amphibians and reptiles present more than one layer or clustered groups of Purkinje neurons, not commonly observed in birds [11]. In a few reptilian species such as snakes, the Purkinje cells are also found scattered in the molecular layer [5]. Nevertheless, comparative investigations representing the important evolutionary transition of cerebellar microanatomy from reptiles to birds are limited.
The present study entails the detailed histomorphometric comparison of reptilian cerebellum with that of birds, an important event in the course of vertebrate evolution. The study is a continuation of our work related to the anatomical evolution of vertebrate cerebellum using representative species of each class of vertebrate in Pakistan [7]. The results provide interesting insights to the anatomical evolution of cerebellum which in turn reflects the sculpting forces that might have led to noticeable functional consequences in Aves as they diverged from reptiles.
Methods![]()
Institutional Review Board for Animal Research and Ethics at Dow University of Health Sciences (DUHS) provided the approval/endorsement to conduct the study. The study was conducted at Dow Research Institute of Biotechnology and Biomedical Sciences (DUHS) from 2017 to 2022.
Specie selection and procurement
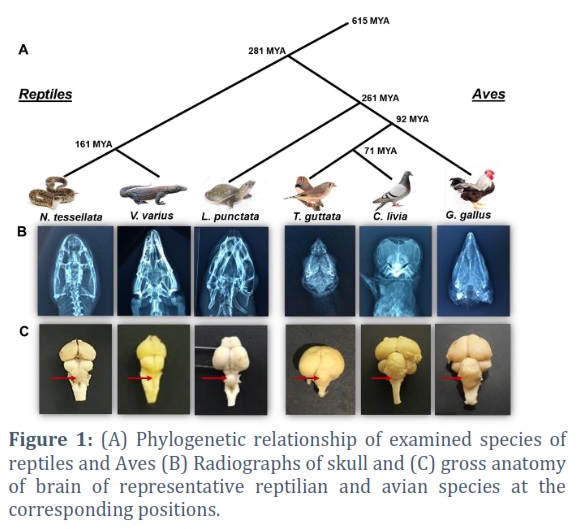
Three representative species of reptiles and Aves were recruited in the study considering their placement in the evolutionary tree (Figure 1A). Monitor lizard (Varanus varius), dice snake and turtle (Lissemys punctata) were studied as representatives of reptiles, whereas chicken (Gallus gallus), zebra finch (Taeniopygia guttata) and pigeon (Columba livia) were included to represent Aves. Three animals of each of the three specie of reptiles and Aves were procured alive from Dow Institute for Advanced Biological and Animal Research (D.U.H.S). Altogether, a total of 9 reptiles and 9 Aves were investigated. Time Tree database was used to estimate the Divergence times between any two species [17].
Radiological examination, dissection and specimen collection
Radiological examination of the head of at least one animal of all investigated species of reptiles and Aves was performed. Plain X-ray films were observed to trace the extent of brain in the intracranial space (Figure 1B). The animals were then sacrificed under appropriate anesthesia and their heads were detached just above the attachment of first cervical spinal nerve. Briefly, brain was exposed by approaching the dorsum followed by opening of the cranial cavity and introducing a cut at meninges. All cranial nerves and attachment of olfactory tract and bulb were severed to facilitate the extraction of whole brain. All the collected samples of brain were kept in 10% Buffered Neutral Formalin (BNF) for fixation.
Morphometric study
The brain was weighed on digital balance to determine the whole brain mass. The volume of whole brain was obtained by using the volume displacement method. The volume of cerebellum was deduced by using the formula for ellipsoid, as follows [18].
V= 0.167 x π x length x width x depth
The entire data points were neutralized against body mass and volume of whole brain of the investigated species.
Tissue processing and staining
The sample of brain and/or cerebellum was cut by a mid-sagittal section into 2 halves. One half of the tissue after processing in an automatic tissue processor were cut on a microtome to obtain 5-7 μm sections. The sections were stained by the conventional hematoxylin and eosin stain (H and E) [19]. The other half of the tissue blocks were subjected to staining and processing by modified impregnation technique of thionin staining [20].
Microscopy
Microscopy of the H and E-stained sections was done at 200x and 1600x magnifications to count the cells (neurons and neuroglia) of granular and molecular layers, Purkinje cell count and to measure the thickness of molecular and granular layers, and Inter-Purkinje distance in 50µm2 area. At least three sections and three different fields per section were taken for each animal for cellular counts and layer thickness, making total of 81 and 243 data points with each species and class, respectively. The sections obtained by formal-thionin block impregnation technique were observed in detail for counting the neurons in molecular and granular layer in 50µm2 area. The Purkinje cell diameter were also measured on formal-thionin stained sections. For Purkinje cell diameter and inter Purkinje distance, total of 90 cells from different section per species were considered. Microscopy was done on Optika B3 camera installed on Leica DM2500 LED microscope. The microphotographs were transferred to Image J v.1.50i software for micrometry.
Statistical Analysis
The normally distributed data were evaluated by Kolmogorov-Smirnov test. The normally distributed data were analyzed by student t-test and the skewed data by Mann-Whitney test. Graph Pad Prism v.8.0.1were used to graphically represent the analyzed data. P-values of 0.05 or less were regarded as statistically significant.
Results![]()
The animal species were recruited based on their evolutionary relationship, so as to include the major lineages of reptiles and Aves (Figure 1A). The radiographic images were observed to determine the expanse of brain in the cranial cavity (Figure 1B). The dissected brain in all reptilian and avian species showed dorsally situated cerebellum which lies caudal to telencephalon/cerebrum. The gross examination showed that reptilian cerebella were sheet like as observed in V. varius or dome shaped in L. punctata and N. tessellata, having smooth surfaces which were devoid of folds or folia. In comparison, the avian cerebella of the three examined species presented multi-lobed, convoluted anatomical configuration with numerous folia (Figure 1C).
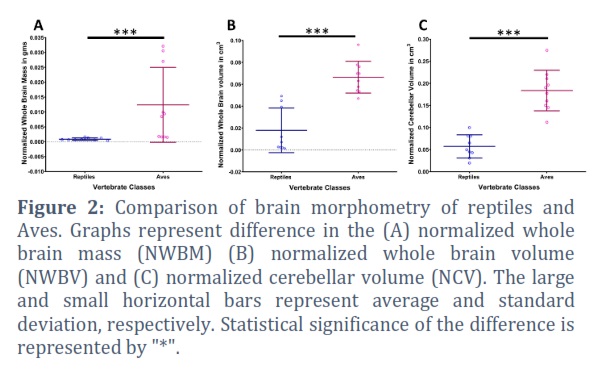
Brain mass was assessed in terms of normalized whole brain mass (NWBM) by dividing whole brain mass against body weight of respective animal. Avian species were found to have higher NWBM with high variability within the group. The reptiles showed lower NWBM with less variability among the species assessed. Comparison of two classes showed statistically significant difference in the NWBM (p=0.0002) (Figure 2A). Nevertheless, normalized whole brain volume (NWBV) of assessed avian species was also found significantly greater (p<0.0001) than reptilian species examined in the study (Figure 2B). The ratio of cerebellar volume and the volume of whole brain was calculated for the normalization of cerebellar volume (NCV). Consistent to the other gross variables, NCV in avian species were found significantly larger (p<0.0001) compared to the investigated reptiles (Figure 2C).
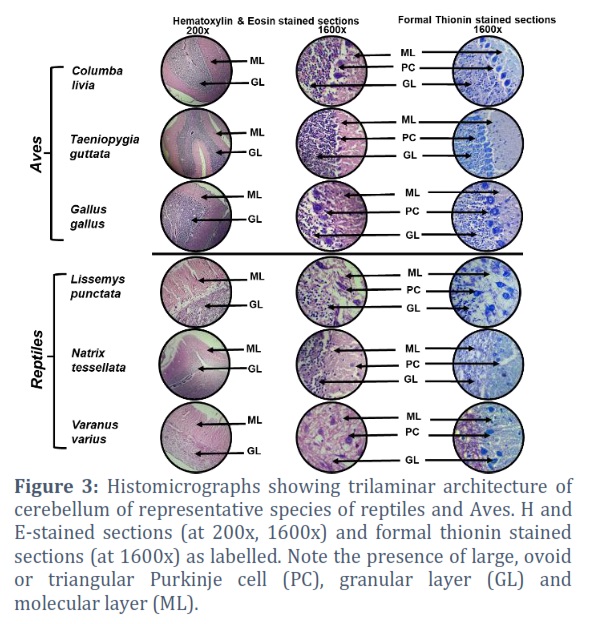
Histological examination of H and E-stained sections of the cerebellar cortices of investigated species showed a well-defined tri-laminar architecture, comprising a superficial molecular layer (ML), an intermediate Purkinje cell (PC) layer and a deeper granular layer (GL). The Purkinje cells are arranged in single (birds) and double (reptiles) layers, sandwiched between the superficial ML and deeper GL. The Purkinje cells showed large, ovoid or triangular soma with processes projecting into ML in both reptilian and avian cerebella. The molecular layer is primarily composed of neuronal fibers/processes with sparse cells (neurons and neuroglia). The granular layer appeared increasingly cellular comprising both neurons and neuroglia (Figure 3). The sections stained with formal-thionin stain exhibited dark blue neurons in all the three layers of cerebellar cortex (Figure 3).
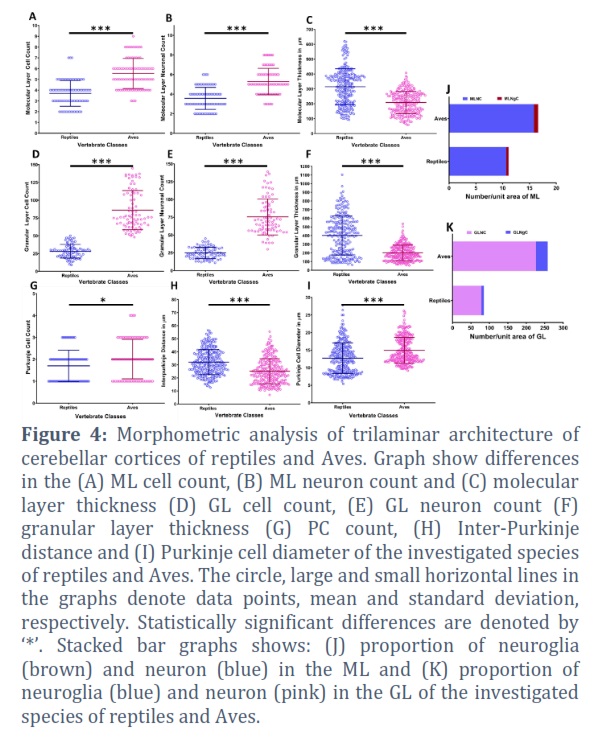
All three investigated species of birds showed high molecular layer cellular and neuronal counts (MLCC and MLNC, respectively) with statistically significant differences (p<0.0001) compared to reptilian species (Figure 4A, B). Conversely, molecular layer thickness (MLT) of Aves was found significantly less (p<0.0001) compared to reptilian species (Figure 4C). In combination this suggests high cellular packaging in the avian cerebellum compared to the reptilian species. Similarly, the granular layer cellular and neuronal counts (GLCC and GLNC, respectively) were also found significantly high (p<0.0001) in Aves compared to reptiles (Figure 4D, E). Like MLT, granular layer thickness (GLT) was also found to be significantly less (p<0.0001) in Aves Figure 4F). This further point to the increment in the cellular packaging in the avian cerebellum compared to reptiles.
The Purkinje cell count (PCC) was observed to be high in Aves with marginal statistical significance (p=0.0332) compared to reptiles (Figure 4G). Similar to PCC, diameter of Purkinje cell (PCD) in investigated avian species was noticeably higher (p<0.0001) than reptiles (Figure 4H). Moreover, Inter-Purkinje distance (IPD) was found maximum in reptiles compared to Aves. In combination this further augment the increased in the cellular concentration in the avian cerebellum compared to reptiles (Figure 4I).
The neuroglia count of molecular layer (MLNgC) was calculated by finding the difference between MLNC and MLCC. Similarly, neuroglia count of granular layer (GLNgC) was obtained by deducting GLNC from GLCC. The ratio between two cell type was obtained (Table 1), which suggest nearly one-fifth fold increment in proportion of neurons to neuroglia in the molecular layer (Figure 4J) and a 2-fold increment in the proportion of neurons to neuroglia in the granular layer of reptiles compared to Aves (Figure 4K).
Figures & Tables

Brain mass of vertebrates is positively aligned with the encephalization quotient (EQ) score, which in turn reflects increased cognition [21]. Humans have estimated EQ score of 6.56 [22], and calibration of NWBM in the examined species suggests noticeably higher EQ scores of 4.35 in Aves and 0.306 reptiles. Therefore, high NWBM and NWBV in Aves as observed in this study points to the increased cognition compared to reptiles. The expansion of avian brain is directly linked with the highly coordinated activity of flight, manipulative motor activity involved in building nests [16], tool manufacture, vocal learning [23] and many other high cognitive performances. Moreover, studies in humans have shown that cerebellar volume accounts for variance in cognitive performance [24]. In this regard it is worth mentioning that highly advanced motor abilities in Aves are linked to the increased cerebellar foliation, which in turn is attributed to the enhanced density of cerebellar neural circuitry and the resultant increment in processing capacity [16,25]. Therefore, the observed increase in NWBV, NWBM and NCV and presence of foliation in avian cerebellum form the anatomical basis of elevated cognitive functions in Aves compared to reptiles.
Previously, positive association between brain size, neuron numbers, and cognitive abilities have been established [26,27]. Consistent to the gross anatomical advancement in the avian cerebellum compared to reptiles, the increased cellular density as observed in this study in different layers of cerebellum further augment the cerebellum driven cognitive functions in Aves compared to reptilian representatives. Our observation of elevated cell and neuron count in molecular and granular layer of cerebellar cortex from reptiles to Aves points to the much-evolved information processing unit of neurons located at ML and GL, which connects the afferent and efferent neurons (PC) of cerebellar cortex [5]. A significant decrease in MLT and GLT from reptiles to Aves with the increased cell count suggests a well evolved cytoarchitecture compacted in the cerebellum of Aves compared to reptiles. Similarly, high PCC and PCD and low IPD in Aves implies to the relatively much evolved inhibitory efferent signals from PC [5]. Consequently, the processing capacity of the cerebellum is increased which can be correlated with the requirement for higher coordination during flight in birds [28].
Neuroglia/Neuron ratio in ML and GL suggests more enriched supportive elements in Aves compared to reptiles in relation to the neurons [29]. Our observation is also consistent with studies suggesting higher number of glial cells per neuron in advance taxa compared to lower taxa [30]. It is evident that neuroglia influences the neurons present in ML and GL by their property of myelination and by modulating the synapses [31]. This in turn modulates the cerebellar cortex output Purkinje neurons, thereby influencing the execution of complex motor tasks. Therefore, it is conceivable that the conundrum of observed deprivation of neuron in proportion to neuroglia in Aves may indeed supports the elevation of coordinated motor activities and cognition in Aves compared to reptiles.
The present study is the first in relation to the comparison of the gross and microanatomy of cerebellum of avian and reptilian species indigenous to Pakistan. In general, it provides interesting insights to the anatomical evolution of avian cerebellum from its ancestral lineage of reptiles. In summary, the findings demonstrate that all gross, trilaminar and cellular dimensions of the avian cerebellum may have played a major role in the improved coordination of motor activities and cognition in the lineage compared to reptiles. However, similar comparative studies especially including other parts of brain may further highlights the involvement of other neural circuitry in this connection.
Competing Interest
The authors declares that there is no conflict of interest regarding the publication of this paper.
![]()
References
- Ren Y, Guo L, Guo CC. A connectivity-based parcellation improved functional representation of the human cerebellum. Scientific Reports, (2019); 9: 1-12.
- Asan AS, McIntosh JR, Carmel JB. Targeting sensory and motor integration for recovery of movement after CNS injury. Frontiers in Neuroscience, (2022); 21;15:1895.
- Schmahmann JD. The cerebellum and cognition. Neuroscience letters, (2019); 688:62-75.
- Parasuram H, Nair B, Naldi G, D’Angelo E, Diwakar S. Understanding cerebellum granular layer network computations through mathematical reconstructions of evoked local field potentials. Annals of Neuroscience, (2018); 25: 11-24.
- Yopak KE, Pakan JMP, Wylie D. The cerebellum of nonmammalian vertebrates: Evolutionary Neuroscience. (2020); pp. 247-260. Academic Press.
- Montgomery JC, Bodznick D, Yopak KE. The cerebellum and cerebellum-like structures of cartilaginous fishes. Brain, Behavior and Evolution, (2012); 80: 152-165.
- Khan N, Perveen K, Hussain M, Qadeer-Malik R, Sharafat S. Comparative Histological Analysis of Cerebellum of Representative Species of Elasmobranchii. International Journal of Morphology, (2023) ;41(2):383-8.
- Fikry H, Saleh LA, Abdel Gawad S. Neuroprotective effects of curcumin on the cerebellum in a rotenone‐induced Parkinson’s disease Model. CNS Neuroscience & Therapeutics, (2022); 28(5):732-48.
- Brown AM, Arancillo M, Lin T, Catt DR, Zhou J, Lackey EP, Stay TL, Zuo Z, White JJ, Sillitoe RV. Molecular layer interneurons shape the spike activity of cerebellar Purkinje cells. Scientific reports, (2019);11;9(1):1-9.
- Zheng J, Yang Q, Makris N, Huang K, Liang J, Ye C, Yu X, Tian M, Ma T, Mou T, Guo W. Three-dimensional digital reconstruction of the cerebellar cortex: lobule thickness, surface area measurements, and layer architecture. The Cerebellum, (2023); 22(2): 249-60.
- Manzano AS, Herrel A, Fabre C, Abdala V. Variation in brain anatomy in frogs and its possible bearing on their locomotor ecology. Journal of Anatomy, (2017); 231: 38-58.
- Puelles L. Contributions to neuroembryology of Santiago Ramon Y Cajal (1852-1934) and Jorge F Tello (1880-1958). International Journal of Developmental Biology, (2009); 53: 1145.
- Pereira SL, Baker AJ. A mitogenomic timescale for birds detects variable phylogenetic rates of molecular evolution and refutes the standard molecular clock. Molecular biology and evolution, (2006); 23:1731-40.
- Waters PD, Patel HR, Ruiz-Herrera A, Álvarez-González L, Lister NC, Simakov O, Ezaz T, Kaur P, Frere C, Grützner F, Georges A. Microchromosomes are building blocks of bird, reptile, and mammal chromosomes. Proceedings of the National Academy of Sciences, (2021); 118:e2112494118.
- Voogd J, Glickstein M. The anatomy of the cerebellum. Trends in Cognitive Sciences, (1998); 2: 307-13.
- Hall ZJ, Street SE, Healy SD. The evolution of cerebellum structure correlates with nest complexity. Biology Letters, (2013); 9: 20130687.
- Kumar S, Stecher G, Suleski M, Hedges SB. TimeTree: a resource for timelines, timetrees, and divergence times. Molecular Biology and Evolution, (2017); 34: 1812-9.
- Lisney TJ, Collin SP. Brain morphology in large pelagic fishes: a comparison between sharks and teleosts. Journal of Fish Biology (2006); 68(2):532-54.
- Bancroft JD, Gamble M. Theory and practice of histological techniques. 2008. Churchill Livingstone, Elsevier, Philadelphia.
- Bancroft JD, Cook HC. Manual of histological techniques and their diagnostic application. 1994. Churchill Livingstone, London.
- Dick U, Roth G. Neuronal factors determining high intelligence. Philosophical Transactions of the Royal Society of London. Series B, Biological Sciences, (2016); 371: 20150180.
- Cairo O. External measures of cognition. Frontiers in Human Neuroscience, (2011); 5: 108.
- Gutierrez-Ibanez C, Iwaniuk AN, Wylie DR. Parrots have evolved a primate-like telencephalic-midbrain-cerebellar circuit. Scientific Reports, (2018); 8: 1-11.
- Hogan MJ, Staff RT, Bunting BP. Cerebellar brain volume accounts for variance in cognitive performance in older adults. Cortex, (2011); 47: 441-450.
- Cunha F, Gutiérrez-Ibáñez C, Racicot K, Wylie DR, Iwaniuk AN. A quantitative analysis of cerebellar anatomy in birds. Brain Structure and Function, (2021) 226:2561-83.
- Herculano-Houzel S. Coordinated scaling of cortical and cerebellar numbers of neurons. Frontiers in Neuroanatomy, (2010); 4: 12.
- Marhounova L, Kotrschal A, Kverkova K, Kolm N, Němec P. Artificial selection on brain size leads to matching changes in overall number of neurons. Evolution, (2019); 73: 2003-2012.
- Sur E, Oznurlu Y, OzaydinT, Çolakoglu F, Unsal S, Yener Y. Comparative histometrical study of the cerebellum and the determination of some AgNor parameters in different avian species. Bulletin of the Veterinary Institute in Pulawy, (2011); 55: 261-265
- Verkhratsky A, Ho MS, Zorec R, Parpura V. The concept of neuroglia. Neuroglia in Neurodegenerative Diseases, (2019); 1-3.
- Bullock TH. The natural history of neuroglia: an agenda for comparative studies. Neuron glia biology, (2004); 1: 97-100.
- Quan L, Uyeda A, Muramatsu R. Central nervous system regeneration: the roles of glial cells in the potential molecular mechanism underlying remyelination. Inflammation and Regeneration, (2022); 42:1-2.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0
![]()