

Full Length Research Article
Neuroprotective effects of ferulic acid and thymoquinone against deltamethrin-induced neurotoxicity in Drosophila melanogaster
Hala Abdulbaki, Mohammad Ali Al-Deeb*
Adv. life sci., vol. 10, no. 2, pp. 289-297, June 2023
*– Corresponding Author: Mohammad Ali Al-Deeb (m_aldeeb@uaeu.ac.ae)
Authors' Affiliations
[Date Received: 11/03/2023; Date Revised: 25/05/2023; Date Published Online: 30/06/2023; Date Updated: 11/09/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Recent research indicates a link between exposure to pyrethroid pesticides and negative neurodevelopmental outcomes such as neurodegenerative disorders, low intelligence quotient (IQ), and attention disorders. Thus this study aims (1) to investigate the effect of exposing Drosophila melanogaster adult male flies to 0.59 µM deltamethrin (DLM) for 24 h on their survival, climbing behavior, and acetylcholinesterase (AChE) activity, as well as the gene expression levels of ple, ddc, dat, aanat1, dop1r1, dop2r, and dopecr genes, and (2) to assess the protective effect of 250 µM Ferulic acid (FA), 25 µM Thymoquinone (TQ), and their combinations on the survival, climbing behavior, and AChE activity of D. melanogaster adult male flies exposed to 0.59 µM DLM for 72 h.
Methods: In the first experiment, adult male wild-type flies were exposed to DLM incorporated into a 10% sucrose solution for 24 h, whereas, in the second experiment, they were exposed to DLM and the individual neuroprotective agents, or their mixture for 72 h.
Results: Flies exposed to DLM exhibited higher mortality rates and shorter climbing distances in the negative geotaxis assay compared to control flies. Additionally, exposure to DLM upregulated the genes ddc, dat, dop1r1, dop2r, and dopecr significantly. Moreover, exposure to DLM for 72 h resulted in higher mortality rates and severe locomotor defects. However, the neuroprotective agents were effective in mitigating these toxic effects. While DLM inhibited AChE activity, concurrent exposure to FA ameliorated the inhibition.
Conclusion: This study demonstrates the efficacy of natural compounds in reducing DLM-induced toxicity in D. melanogaster and highlights the potential use of this model in studying and developing therapeutic strategies for movement disorders.
Keywords: Drosophila melanogaster; Neurotoxicity; Ferulic acid; Thymoquinone; Deltamethrin; Negative geotaxis
Introduction![]()
Deltamethrin (DLM) is a synthetic type II pyrethroid with broad-spectrum insecticide properties. Besides protecting crops and golf courses, it is also used to control animal ticks and vector-borne diseases. Due to its high efficiency, DLM has become one of the most widely used pesticides for household pest control. Thus, numerous individuals are being exposed to DLM, which can lead to serious health problems [1 , 2]. Based on epidemiological and animal studies, long-term exposure to low doses of DLM was associated with behavioral and molecular abnormalities that may lead to neurodegenerative disorders, developmental deficits, and learning impairments [3 , 4]. Similar to other pyrethroids, DLM primarily acts by prolonging voltage-gated sodium channel opening, triggering repetitive firing of neurons followed by depolarizing blockage of neurons, resulting in subsequent insect death. A variety of other functions can also be exerted by DLM on biological membranes, including voltage-gated calcium channels, chloride channels, and GABAA receptors [5 , 6]. It has been shown that DLM induces oxidative stress, which contributes to pathological cell death in numerous neurodegenerative diseases, such as Alzheimer’s disease and Parkinson’s disease (PD) [7 , 8 , 9]. It is likely that the general population is readily exposed to DLM and other chemicals, either through direct or indirect exposure to pesticide-treated food. Understanding how these pesticides are damaging the nervous system is critical to know the mechanisms by which they do so.
Phytochemicals are plant-based chemicals that exhibit significant health-promoting properties. Due to their antioxidant properties, phytochemicals have received considerable attention in recent years for their role in protecting cells against environmental toxins [10 , 11]. In several clinical trials, phytochemicals have shown promising results as neuroprotective agents targeting the treatment of neurodegenerative disorders [12 , 13]. Ferulic acid (FA), a naturally occurring phenolic compound that belongs to the hydroxycinnamic acid family, is found mainly in plants. It is a well-studied natural molecule with powerful neuroprotective effects [14]. The antioxidant property of FA is well known due to its free radical scavenging effect. In a recent study, FA attenuated neuroinflammation and improved behavioral deficits in the 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) mouse model of PD [15]. Through antioxidant and anti-inflammatory properties, FA has also been shown to have a promising neuroprotective effect against rotenone-induced neurodegeneration in rats as a model for PD [16]. Aside from its antioxidant and anti-inflammatory properties, FA also regulates various neuro-signaling pathways through interactions with multiple receptors or enzymes. Additionally, it regulates a variety of pro-inflammatory cytokines expression as well as pro-apoptotic signals, which may explain its neurotherapeutic action [17]. Furthermore, thymoquinone (TQ) is one of the major bioactive components of Nigella sativa essential oil commonly found in the Middle East, and Africa. Numerous studies have demonstrated that TQ has a variety of potent pharmacological effects, including antioxidant, and anti-inflammatory [18]. Moreover, TQ has been shown to reduce the levels of intracellular reactive oxygen species (ROS) in neuronal cells, and to downregulate pro-inflammatory cytokines [19 , 20]. A potential protective effect of TQ on dopaminergic neurons was reported against 1-methyl-4-phenylpyridinium ions and rotenone cytotoxicity in cell culture experiments, highlighting its antioxidant role in cells protection [21]. In rotenone-animal models of PD, TQ suppressed Parkinson’s symptoms, including dyskinesia [22]. Furthermore, TQ was shown to ameliorate the toxicity induced by 1-methyl-4-phenylpyridinium or rotenone in primary cultured dopaminergic neurons [23 , 24]. Combination therapy or multi-component therapy involves combining two or more therapeutic agents to enhance their biochemical activity by exploiting additive effects, or better yet, greater-than-additive effects, between the two agents. A clear understanding of the interactions within and between natural product mixture components is essential for achieving specific biological effects that can provide valuable resources for drug development [25].
Drosophila melanogaster (fruit fly) is a versatile model organism that has been used extensively for biological research to study diverse disciplines ranging from fundamental genetics to tissue and organ development [26 , 27]. Together, the Drosophila model system with the feasible genetic and molecular tools, made the fruit flies serve as a valuable in vivo model to study the effects of environmental toxicants and to screen potential therapeutic drugs [11 , 28 , 29, 30 , 31 , 32].
Since antioxidants have been shown to effectively alleviate DLM-induced toxicity in previous interventions [5], along with FA and TQ, they have also been widely reported to possess neuroprotective properties due to their antioxidant effects. Therefore, the objectives of this study are twofold: (1) to investigate the effect of exposing D. melanogaster adult male flies to 0.59 µM DLM for 24 h on their survival, climbing behavior, and AChE activity, as well as the gene expression levels of tyrosine hydroxylase (ple), DOPA decarboxylase (ddc), DA transporter (dat), D1-like receptor 1 (dop1r1), D2-like receptor (dop2r), DA/ecdysteroid receptor (dopecr), and DA arylalkylamine N-acetyltransferase (aanat1) and (2) to assess the protective effect of 250 µM FA, 25 µM TQ, and their combinations on the survival, climbing behavior, and AChE activity of D. melanogaster adult male flies exposed to 0.59 µM DLM for 72 h. However, gene expression analysis was not performed in the second objective due to time and resource constraints.
Methods![]()
Chemicals
The following chemicals were acquired from Sigma Aldrich (St. Louis, MO, USA): deltamethrin (PESTANAL®), trans-ferulic acid, thymoquinone, and acetylthiocholine iodide, as well as 5′,5′-dithiobis(2-nitrobenzoic acid) (DTNB). In addition, dimethyl sulfoxide (DMSO) with a purity of 99.9% was obtained from Heiltropfen, Germany.
Fly strain and rearing
Throughout all of our experiments, we used wild-type D. melanogaster fruit flies (Oregon R strain). For a more in-depth explanation of the rearing conditions, we refer to our prior research [33]. In summary, the flies were reared on instant Drosophila medium while being subjected to controlled environmental conditions, including a temperature of 23±1°C and a 12:12 h light-dark cycle.
Exposure to DLM for 24 h
In a previous publication [33], we described and used a feeding device to expose flies to chlorpyrifos, which was dissolved in a 10% sucrose solution. In short, male flies that were synchronized in age (4-6 days old) were briefly anesthetized with cold and transferred into each feeding device. All experiments were conducted in triplicate, and each experiment was replicated three times (n = 3) with new vials to prevent any contamination. Therefore, each experiment contained three feeding devices for the insecticide treatment (0.59 µM DLM) and three devices for the control (sucrose solution containing DMSO). The concentration of DMSO in the control group was equivalent to that of the DLM treatment group and did not result in any significant mortality or disruption in the flies' movement (data not shown).
Exposure to DLM and neuroprotective agents for 72 h
In this experiment, the flies were exposed to eight treatments incorporated into 10% sucrose solution using a feeding device for 72 h: (1) control (DMSO); (2) 0.59 μM DLM; (3) 250 μM FA; (4) 25 μM TQ; (5) FA+TQ; (6) DLM+FA; (7) DLM+TQ; and (8) DLM+FA+TQ. The DLM, FA, and TQ were first dissolved in DMSO and then diluted to the specific concentration using the sucrose solution. Each feeding device was charged with fresh chemicals every 24 h.
Assessment of fly survival
Survival rates of flies were recorded 24-72 h after introducing the flies to feeding devices. Dead flies were identified by lack of coordinated movement. Surviving flies were used for subsequent tests.
Negative geotaxis assay
The test used a climbing device made of two plastic vials held together with tape, and a digital camera was used to record fly behavior. Flies were collected with an aspirator and allowed to acclimate for 30 min before being tapped down to the bottom of the vial and timed for 8 s of climbing. The experiment was repeated five times for technical replicates and three times for biological replicates. Six climbing devices were used simultaneously, and the distances climbed by each fly were measured using PAST software [33].
RT-qPCR
We adhered to the same procedure utilized in our previous study [33]. To elaborate, we extracted RNA from the flies and then used 50 ng of total RNA in triplicate with the Luna® Universal One-Step RT-qPCR Kit (New England Biolabs Inc., Ipswich, USA) for RT-qPCR. The reaction was conducted in a QuantStudio 5 Real-Time PCR system (Applied Biosystems) following the manufacturer's instructions. We evaluated the relative transcript level of ple, ddc, dat, aanat1, dop1r1, dop2r, and dopecr genes through the 2(-ΔΔCt) method, where glyceraldehyde-3-phosphate dehydrogenase (GAPDH) served as the reference gene for normalization [34].
Determination of AChE activity
The method of Ellman et al. [38] was utilized to evaluate the activity of acetylcholinesterase. To start, fly samples were homogenized at a ratio of 1:100 (flies(mg)/volume µL PBS (pH 7.4) + protease inhibitor cocktail) and then centrifuged at 5000 rpm for 5 min at 4°C. The resulting supernatant was then used as the source of the enzyme. The reaction mixture consisted of 80 µL of phosphate-buffered saline (PBS) (pH 7.4), 50 µL of 0.32 mM DTNB, 20 µL of the sample (supernatant), and 50 µL of 10 mM acetylthiocholine. The optical densities (ODs) were read for 10 minutes at a 405 nm wavelength using a Platos-R-496-AMP AMEDA microplate reader (Labordiagnostik GmbH, Graz, Austria). Acetylcholinesterase activity was calculated using the following formulas [39]:
% inhibition = (Control-Treatment/Control)*100 (Eq. 1)
Remaining activity = 100 – % inhibition. (Eq. 2)
Statistical analysis
We performed all statistical analyses using GraphPad Prism 9 for Windows (GraphPad Software, San Diego, CA, United States). To identify differences between the treatment groups and the control group, a two-tailed Student's t-test was employed. A one-way ANOVA was used to analyze the AChE % activity, followed by the Bonferroni post hoc test. Gene expression data was analyzed using a two-way ANOVA followed by the Bonferroni post hoc test. Any differences with a p-value of less than 0.05 were considered statistically significant. The data was presented as mean ± standard error of the mean (SEM).
Results![]()
Effect of DLM on fly survival
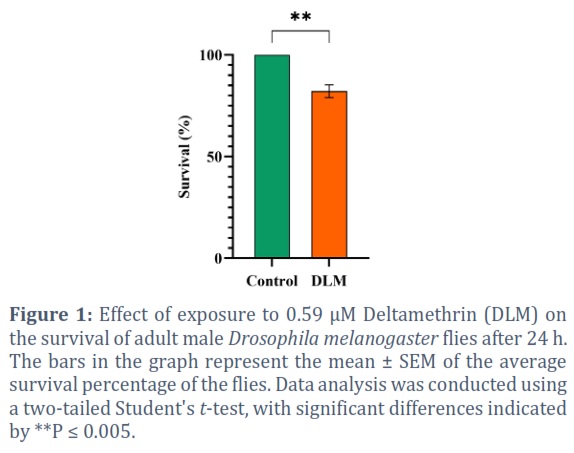
Male adult flies exposed to 0.59 µM of DLM for 24 h exhibited a significant decline in the percentage of surviving flies (82.2%) in comparison to the control group (100%) (P = 0.0047) (Figure 1).
Effect of DLM on locomotor performance
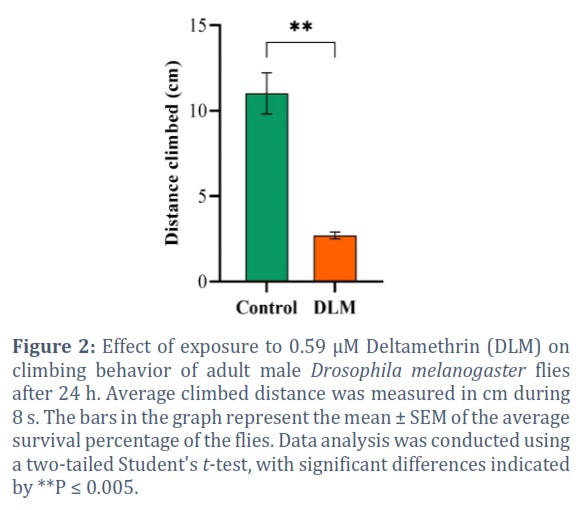
Wild-type flies exposed to DLM for 24 h displayed a significant defect in reaching the top of the climbing apparatus as compared to the control flies. The average distance climbed by the untreated flies was 11.01 cm compared to 2.70 cm in DLM-treated flies during 8 s (P = 0.0025) (Figure 2).
Effect of DLM on AChE activity
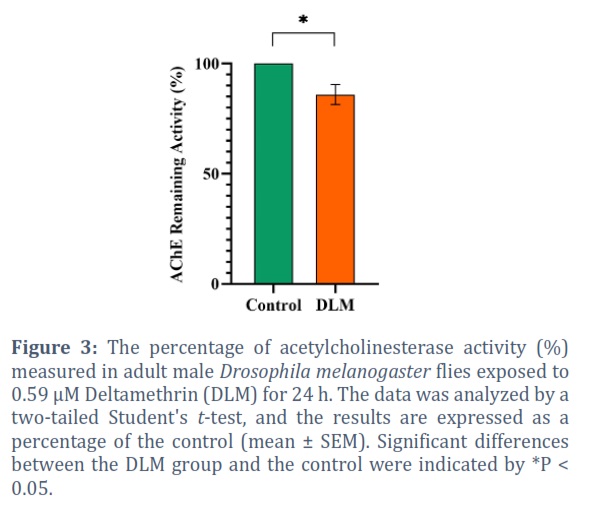
A significant difference was detected in AChE activity after 24 h of exposure to DLM, the enzyme activity exhibited a 9.56 % (P = 0.0476) decrease compared to the control (Figure 3).
Effect of DLM on gene expression profile of dopaminergic system
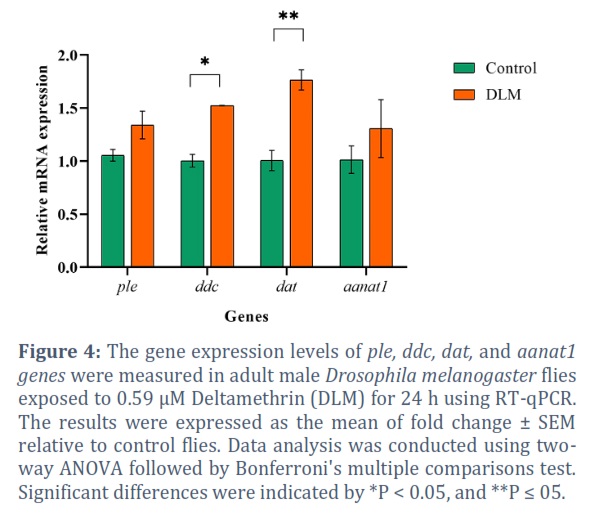
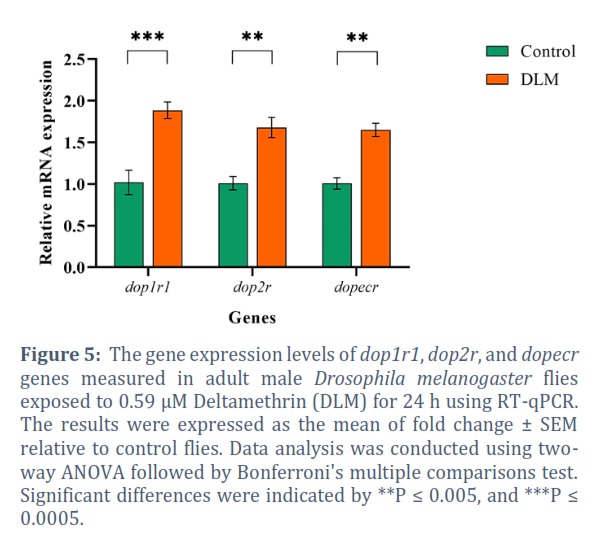
Exposure to DLM caused a significant upregulation effect on dop1r1 85.20% (P = 0.0002), dat 75.85% (P = 0.0082), ddc 51.66% (P = 0.0353) (Figure 4), dop2r 67.06% (P = 0.0039), and dopecr 64.16% (P = 0.0058) (Figure 5). Although no statistically significant differences were found among the other genes, DLM produced upregulation in all of them with 26.90% ple and 28.74% aanat1 (Figure 4).
Effect of neuroprotective agents on survival of DLM-treated flies
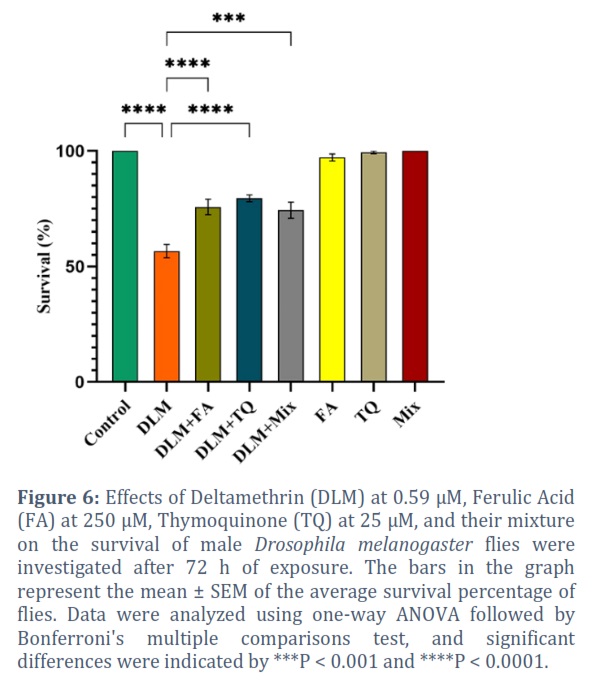
Flies exposed to 0.59 µM DLM for 72 h had a significant decrease in survival 56.67% compared to the control 100% (P < 0.0001) (Figure 6). Co-exposure of flies to 0.59 µM DLM, 250 µM FA, 25 µM TQ, and their combination (FA + TQ), significantly improved the survival percentage by 19.1% (P < 0.0001), 22.9 % (P < 0.0001), and 17.7% (P = 0.0002), respectively (Figure 6). Neither FA, TQ, nor their combined administration had a significant effect on the survival of the flies, with 97.19%, 99.44, and 100.00%, respectively (Figure 6).
Locomotor performance of flies exposed to DLM and neuroprotective agents
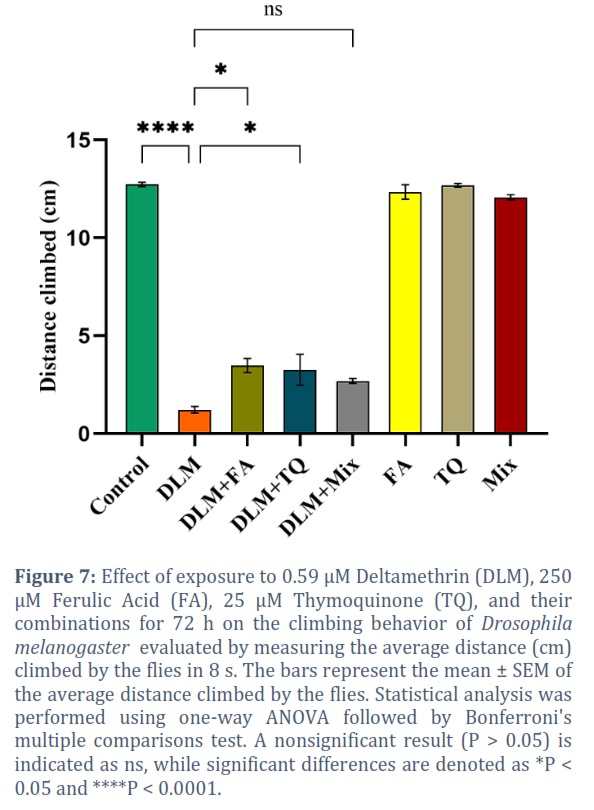
Exposing the flies to 0.59 µM DLM for 72 h displayed a big defect in reaching the top of the climbing apparatus as compared to control flies indicating locomotor dysfunction. The average distance climbed by the control group was 12.73 cm compared to 1.22 cm in DLM treated group during 8 s (P < 0.0001) (Figure 7). Co-exposure of flies to 250 µM FA, 25 µM TQ, and their combination prevented locomotor impairment induced by the DLM (Figure 7). Flies exposed to DLM+ FA, DLM+ TQ, and DLM+ Mix climbed higher distances 3.48 (P = 0.0343), 3.27 (P = 0.0304), and 2.70 cm (P = 0.2977), respectively than flies exposed to DLM alone (Figure 7). The FA, TQ, and their combined administration did not have a significant effect on the locomotor performance of flies with 12.35, 12.69, and 12.07 cm climbing distances, respectively (Figure 7).
Effect of neuroprotective agents on the AChE activity in flies exposed to DLM
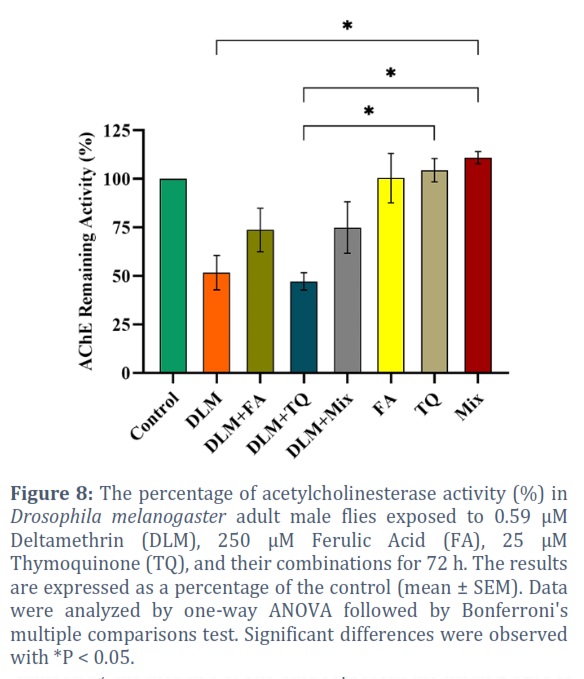
The AChE activity was inhibited with a 48.30% reduction in the flies exposed to DLM for 72 h (P = 0.032) (Figure 8). Co-exposure to the neuroprotective agents did not completely restore the enzyme activity compared to the control. However, FA alleviated the inhibition to 26.22%. Whereas, TQ did not show any improvement in the AChE activity, yet, the combined effect of FA and TQ alleviated the inhibition to 25.04%. The neuroprotective agents did not show any adverse effect on the AChE enzyme when administered alone.
Discussion![]()
The harmful effects of insecticides on organisms, particularly commonly used ones, are a pressing concern, as exposure to these chemicals in the environment can damage biological systems. The present study evaluated the effects of DLM, a major pyrethroid insecticide, on the expression of dopaminergic genes, AChE activity, and impairment of behavior in D. melanogaster. Additionally, it investigated the possible ameliorative mechanism of FA and TQ as natural compounds. Our data revealed that DLM caused damage to the exposed flies, and FA and TQ mitigated some of the harmful effects.
Deltamethrin is the most powerful synthetic pyrethroid that is easily available to the general consumer market. Exposure to DLM and other pyrethroids has become widespread mainly because they were used as substitutes for organophosphorus insecticides [8 , 40]. Pyrethroids have quick access to tissues, including the central nervous system (CNS), due to their lipophilic nature. Thus, it is possible that even small doses can produce significant biological and pathophysiological effects. In recent years, pyrethroid exposure has been linked to neurophysiological and neurobehavioral disorders in humans [41]. The generation of free radicals by DLM leads to behavioral changes mediated by the altered action of neurotransmitters [42]. In recent years, plant-based foods and their active components have attracted scientific attention for their health-promoting properties. The free radical scavenging activity of natural products appears to play an important role in their antioxidant activity; thus, the role of natural compounds had recently received considerable attention as a dietary antioxidant. In different experimental studies, FA and TQ were reported to have neuroprotective effects [16 , 22]. Hence, the findings of the current study shed light on some aspects of the neurotoxicity of DLM.
We initially studied the effects of DLM on the locomotor, dopaminergic, and cholinergic systems of D. melanogaster. A 24-hour exposure to DLM significantly reduced fly survival rate and climbing abilities of surviving flies. Similar observations were reported in rats exposed to DLM that exhibited decreased locomotor activity [43 , 44]. Additionally, a significant reduction in AChE activity was observed. Deltamethrin caused alteration in the expression of dopamine (DA) -related genes. We observed significant upregulation of DA receptor genes with the highest upregulation observed in dop1r1 followed by dop2r, dopecr, dat and the ddc. Even though the other genes did not exhibit significant changes, there was a slight upregulation following the same trend of the affected genes. It is likely that DLM induced oxidative stress, which in turn, induced rapid upregulation of all genes. Another explanation is that DLM depleted the DA content in D. melanogaster and activated the negative feedback mechanism in which genes responsible for DA biosynthesis (ple, and ddc) were upregulated to compensate for the loss of DA and DA- receptors (dop1r1, dop2r, and dopecr) were upregulated as a response of increased dat expression. In fact, dat upregulation is documented as a sign of pyrethroid exposure and toxicity. Additionally, the gene responsible for DA metabolism (aanat1) was upregulated consolidating the DA loss hypothesis. One experiment showed that in dat overexpressing mice, the levels of D1 and D2 receptors were significantly increased, suggesting that changes in DA-receptor found in DLM-exposed mice are the result of upregulation in response to an increase in the number of DAT and, subsequently, a decrease in extracellular levels of DA [45]. Consistent with our findings, a study reported that developmental DLM exposure increased striatal D1 levels in male mice [45]. On the contrary, larval fish exposed to DLM developmentally showed decreased expression of dr1d1, increased levels of tyrosine hydroxylase (TH), and decreased drd2a transcripts. A significant increase in larval swim activity was observed after DLM application to zebrafish [46]. Several studies reported a reduction in mRNA of TH in cell lines [47] and other vertebrate model organisms [46 , 47 48]. Our results differ from those of the previously mentioned studies and the differences might be attributed to the differential responses of developing and adult organisms to toxins, as well as the differences among various animal species and the duration of exposure. In both studies, DLM modulates TH expression, thereby altering DA biosynthesis. It was shown that exposure to the lowest concentration of DLM resulted in increased TH expression immediately after exposure. On the other hand, repeated exposure to DLM has been shown to decrease TH mRNA and protein expression in adult male rats [47]. Although the D1 and D2 auto-receptors have opposite effects on cAMP signal transduction, they act synergistically to modulate locomotor activity [49]. A change in D1 receptor signal transduction often leads to corresponding changes in D2 receptor-mediated responses; D1 regulates the sensitivity of D2 [50]. Therefore, it is plausible that a sustained increase in dop1r1 transcripts could promote a subsequent upregulation of dop2r at the transcriptional level, which we observed in our results. A 72 h exposure to the same DLM concertation led to twice as much mortality as a 24 h exposure. Additionally, loss of coordinated movements and severe climbing defects were observed.
Exposing the D. melanogaster to DLM caused about a 50% reduction in AChE activity after 72 h compared to the control group, but it was not statistically significant using the Bonferroni method. This finding was not surprising because the Bonferroni method considers all the comparisons (28 comparisons in this study) and this may result, in some cases, in having insufficient statistical power. Nonetheless, it should be mentioned that when we used the Dunnett method, which compared all the treatments to the control only, we detected significant differences between the DLM and the control. Long-term exposure resulted in higher mortality and more damage to the nervous system as evidenced by severely impaired locomotor performance and AChE activity. According to previous studies, DLM-exposed rats had significantly lower AChE activity than the control group [8, 51]. The inhibitory effect of DLM on AChE activity in rats was previously explained by its ability to reduce the ACh binding space on AChE’s aromatic, hydrophobic surface, probably due to its lipophilicity [9]. The excessive inhibition of AChE can cause nervous system malfunction, leading to neurodegenerative disorders [8]. Researchers have suggested that voltage-gated calcium channels, chloride channels, and GABA receptors might modulate neurotransmitter release, particularly ACh, DA, and serotonin, in the acute and chronic manifestations of pyrethroid-induced neurotoxicity [52]. In rats, DLM exposure significantly decreased the content of DA in the striatum [48].
In this study, co-exposure to DLM and FA led to normalization of AChE levels, although the change was not significant possibly due to the concentration used. Future studies should optimize concentrations to maximize benefits and minimize side effects. On the other hand, TQ at 25 µM did not improve the enzyme activity. Despite this, it protected against DLM-induced mortality and climbing deficits in our study. Yet, its protective effect might be due to another mechanism of action that affects the AChE activity. Combining FA and TQ did not further improve the enzyme activity indicating that the neuroprotective effect was due to the individual effect of FA alone. There were no significant differences between the control and DLM co-administered with FA and the combined FA+TQ. This means that FA was able to restore some of the enzyme activity. Since TQ did not interfere with FA neuroprotective effects on AChE, it is clear that it is not likely to interfere with FA mechanisms of protection. There was no change in any of the parameters evaluated when flies were exposed to pure neuroprotective agents without DLM, indicating that the concentrations selected in the current study were safe. The results of some studies indicate that FA treatment protects DA neurons from rotenone-induced toxicity [16]. One more study showed that FA was able to reverse the toxic effect of cadmium-induced brain damage on cholinesterase activities in rats [53]. An earlier study found that TQ was able to rescue levels of AChE inhibited by malathion, which is an organophosphate insecticide (OP) [54]. Our study; however, did not demonstrate a reverse effect of TQ on the inhibitory effect of DLM on AChE. Probably we used a dose that was insufficient to reverse the inhibition. Another reason could be related to the different mechanisms of action between DLM and OPs.
Our study found that DLM damages the nervous system of D. melanogaster, leading to locomotor dysfunction and increased mortality rates. Expression of DA-related genes was also affected by DLM exposure. However, co-administration of neuroprotective agents FA and TQ reversed the effects of DLM. FA and TQ improved survival and negative geotaxis performance, likely due to their antioxidant and neuroprotective properties. No additional benefits were observed from combining FA and TQ. Further studies are recommended to better understand the link between DA content and behavioral deficits, as well as the neuroprotective effect of FA and TQ on dopaminergic pathways.
In conclusion, the findings of the present study have shown that DLM at 0.59 µM was able to cause severe damage in adult D. melanogaster, including increased mortality, locomotor deficits, inhibition of AChE, and disturbance of dopaminergic gene expression. A disturbance was observed in two main systems involved in movement coordination. This study illustrates some of the mechanisms through which DLM can cause neurotoxicity. Here we reported for the first time the neuroprotective action of FA and TQ against the DLM-induced neurotoxicity in D. melanogaster. The FA alleviated the inhibition of AChE that was caused by DLM, which attributes a neuroprotective potential to this natural compound. Consequently, FA and TQ may offer a promising source of potential therapeutic agents for the treatment of DLM intoxications.
Conflict of Interest
The authors declare that there is no conflict of interest.
Acknowledgement
This study was funded by the Master’s Program of the Biology Department at the UAE University, and we would like to express our thanks for their support.
![]()
References
- El Golli-Bennour E, Timoumi R, Annaibi E, Mokni M, Omezzine A, Bacha H, Abid-Essefi S. Protective effects of kefir against deltamethrin-induced hepatotoxicity in rats. Environmental Science and Pollution Research, (2019); 26(18): 18856–18865.
- Han B, Lv Z, Zhang X, Lv Y, Li S, Wu P, Yang Q, Li J, Qu B, Zhang Z. Deltamethrin induces liver fibrosis in quails via activation of the TGF-β1/Smad signaling pathway. Environmental Pollution, (2020); 259: 113870.
- Moser VC. Animal models of chronic pesticide neurotoxicity. Human & Experimental Toxicology, (2007); 26(4): 321–332.
- Tewari A, Banga HS, Gill JPS. Sublethal chronic effects of oral dietary exposure to deltamethrin in Swiss albino mice. Toxicology and Industrial Health, (2018); 34(6): 423–432.
- Abdel-Daim MM, Abuzead SMM, Halawa SM. Protective role of Spirulina platensis against acute deltamethrin-induced toxicity in rats. PloS one, (2013); 8(9): e72991–e72991.
- Romero A, Ares I, Ramos E, Castellano V, Martínez M, Martínez-Larrañaga MR, Anadón A, Martínez MA. Evidence for dose-additive effects of a type II pyrethroid mixture. In vitro assessment. Environmental Research, (2015); 138: 58–66.
- Li HY, Wu SY, Ma Q, Shi N. The pesticide deltamethrin increases free radical production and promotes nuclear translocation of the stress response transcription factor Nrf2 in rat brain. Toxicology and industrial health, (2011); 27(7): 579–590.
- Mani VM, Sadiq AMM. Naringin modulates the impairment of memory, anxiety, locomotor, and emotionality behaviors in rats exposed to deltamethrin; a possible mechanism association with oxidative stress, acetylcholinesterase and ATPase. Biomedicine & Preventive Nutrition, (2014); 4(4): 527–533.
- Mohammadi H, Ghassemi-Barghi N, Malakshah O, Ashari S. Pyrethroid exposure and neurotoxicity: a mechanistic approach. Archives of Industrial Hygiene and Toxicology, (2019); 70(2): 74–89.
- Park JH, Jung JW, Ahn Y-J, Kwon HW, Neuroprotective properties of phytochemicals against paraquat-induced oxidative stress and neurotoxicity in Drosophila melanogaster. Pesticide Biochemistry and Physiology, (2012); 104(2): 118–125.
- Farombi EO, Abolaji AO, Farombi TH, Oropo AS, Owoje OA, Awunah MT. Garcinia kola seed biflavonoid fraction (Kolaviron), increases longevity and attenuates rotenone-induced toxicity in Drosophila melanogaster. Pesticide Biochemistry and Physiology (2018); 145: 39–45.
- Seidl SE, Potashkin JA. The promise of neuroprotective agents in Parkinson’s disease. Frontiers in neurology, (2011); 2: 68.
- de Andrade Teles RB, Diniz TC, Costa Pinto TC, de Oliveira Júnior RG, Gama E Silva M, de Lavor ÉM, Fernandes AWC, de Oliveira AP, de Almeida Ribeiro FPR, da Silva AAM, et al. Flavonoids as Therapeutic Agents in Alzheimer’s and Parkinson’s Diseases: A Systematic Review of Preclinical Evidences. Oxidative medicine and cellular longevity, (2018); 2018: 7043213.
- Pandey KB, Rizvi SI. Plant polyphenols as dietary antioxidants in human health and disease. Oxidative medicine and cellular longevity, (2009); 2(5): 270–278.
- Li X, Zhang J, Rong H, Zhang X, Dong M. Ferulic Acid Ameliorates MPP+/MPTP-Induced Oxidative Stress via ERK1/2-Dependent Nrf2 Activation: Translational Implications for Parkinson Disease Treatment. Molecular Neurobiology, (2020); 57(7): 2981–2995.
- Ojha S, Javed H, Azimullah S, Abul Khair SB, Haque ME. Neuroprotective potential of ferulic acid in the rotenone model of Parkinson’s disease. Drug design, development and therapy, (2015); 9: 5499–5510.
- Thapliyal S, Singh T, Handu S, Bisht M, Kumari P, Arya P, Srivastava P, Gandham R. A Review on Potential Footprints of Ferulic Acid for Treatment of Neurological Disorders. Neurochemical Research, (2021); 46(5): 1043–1057.
- Ramadan MF. Nutritional value, functional properties and nutraceutical applications of black cumin (Nigella sativa L.): an overview. International Journal of Food Science & Technology, (2007); 42(10): 1208–1218.
- Akram Khan M, Afzal M. Chemical composition of Nigella sativa Linn: Part 2 Recent advances. Inflammopharmacology, (2016); 24(2–3): 67–79.
- Isaev NK, Chetverikov NS, Stelmashook E V, Genrikhs EE, Khaspekov LG, Illarioshkin SN. Thymoquinone as a Potential Neuroprotector in Acute and Chronic Forms of Cerebral Pathology. Biochemistry (Moscow), (2020); 85(2): 167–176.
- Sedaghat R, Roghani M, Khalili M. Neuroprotective effect of thymoquinone, the nigella sativa bioactive compound, in 6-hydroxydopamine-induced hemi-parkinsonian rat model. Iranian journal of pharmaceutical research: IJPR (2014); 13(1): 227.
- Farkhondeh T, Samarghandian S, Shahri AMP, Samini F. The Neuroprotective Effects of Thymoquinone: A Review. Dose-response : a publication of International Hormesis Society, (2018); 16(2): 1559325818761455–1559325818761455.
- Radad K, Moldzio R, Taha M, Rausch W-D. Thymoquinone protects dopaminergic neurons against MPP+and rotenone. Phytotherapy Research, (2009); 23(5): 696–700.
- Ardah MT, Merghani MM, Haque ME. Thymoquinone prevents neurodegeneration against MPTP in vivo and modulates α-synuclein aggregation in vitro. Neurochemistry International, (2019); 128: 115–126.
- Caesar LK, Cech NB. Synergy and antagonism in natural product extracts: when 1 + 1 does not equal 2. Natural product reports, (2019); 36(6): 869–888.
- Jana SC, Bettencourt-Dias M, Durand B, Megraw TL. Drosophila melanogaster as a model for basal body research. Cilia, (2016); 5: 22.
- Ugur B, Chen K, Bellen HJ. Drosophila tools and assays for the study of human diseases. Disease models & mechanisms, (2016); 9(3): 235–244.
- Pandey UB, Nichols CD. Human disease models in Drosophila melanogaster and the role of the fly in therapeutic drug discovery. Pharmacological reviews, (2011); 63(2): 411–436.
- Sudati JH, Vieira FA, Pavin SS, Dias GRM, Seeger RL, Golombieski R, Athayde ML, Soares FA, Rocha JBT, Barbosa NV. Valeriana officinalis attenuates the rotenone-induced toxicity in Drosophila melanogaster. NeuroToxicology, (2013); 37: 118–126.
- Hossain MS, Sapon MA. Drosophila as a Model for Analyzing of Human Genetic and Pathogenic Related Diseases. International Journal of Innovation and Scientific Research, (2014); 12: 126-134.
- Panchal K, Tiwari AK. Drosophila melanogaster “a potential model organism” for identification of pharmacological properties of plants/plant-derived components. Biomedicine & Pharmacotherapy, (2017); 89: 1331–1345.
- Cheng L, Baonza A, Grifoni D. Drosophila Models of Human Disease. BioMed research international, (2018); 2018: 7214974.
- Abdulbaki H, Al-Deeb M. Chlorpyrifos-induced dopaminergic damage in Drosophila melanogaster assessed by gene expression , AChE assay , and negative geotaxis. Genetics and Molecular Reserch, (2022); 21(3).
- Livak KJ, Schmittgen TD. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods, (2001); 25(4): 402–408.
- Norry FM, Larsen PF, Liu Y, Loeschcke V. Combined expression patterns of QTL-linked candidate genes best predict thermotolerance in Drosophila melanogaster. Journal of Insect Physiology, (2009); 55(11): 1050–1057.
- Figueira FH, de Quadros Oliveira N, de Aguiar LM, Escarrone AL, Primel EG, Barros DM, da Rosa CE. Exposure to atrazine alters behaviour and disrupts the dopaminergic system in Drosophila melanogaster. Comparative Biochemistry and Physiology Part C: Toxicology & Pharmacology, (2017); 202: 94–102.
- Inagaki HK, Ben-Tabou de-Leon S, Wong AM, Jagadish S, Ishimoto H, Barnea G, Kitamoto T, Axel R, Anderson DJ. Visualizing neuromodulation in vivo: TANGO-mapping of dopamine signaling reveals appetite control of sugar sensing. Cell, (2012); 148(3): 583–595.
- Ellman GL, Courtney KD, Andres V, Feather-Stone RM. A new and rapid colorimetric determination of acetylcholinesterase activity. Biochemical pharmacology, (1961); 7: 88–95.
- Anjum SI, Yousf MJ, Ayaz S, Siddiqui BS. Toxicological evaluation of chlorpyrifos and Neem extract (Biosal B) against 3RD instars larvae of Drosophila melanogaster. Journal of Animal & Plant Sciences, (2010); 20(1): 9–12.
- Pérez JJ, Williams MK, Weerasekera G, Smith K, Whyatt RM, Needham LL, Barr DB. Measurement of pyrethroid, organophosphorus, and carbamate insecticides in human plasma using isotope dilution gas chromatography-high resolution mass spectrometry. Journal of chromatography. B, Analytical technologies in the biomedical and life sciences, (2010); 878(27): 2554–2562.
- Mani VM, Gokulakrishnan A, Sadiq AM. Molecular Mechanism of Neu-rodevelopmental Toxicity Risks of Occupational Exposure of Pyrethroid Pesticide with Reference to Deltamethrin-A Critical Review. BAOJ Pathology, (2017); 1(008).
- Wolansky MJ, Harrill JA. Neurobehavioral toxicology of pyrethroid insecticides in adult animals: A critical review. Neurotoxicology and Teratology, (2008); 30(2): 55–78.
- Lazarini CA, Florio JC, Lemonica IP, Bernardi MM. Effects of prenatal exposure to deltamethrin on forced swimming behavior, motor activity, and striatal dopamine levels in male and female rats. Neurotoxicology and Teratology, (2001); 23(6): 665–673.
- Johri A, Yadav S, Singh RL, Dhawan A, Ali M, Parmar D. Long lasting effects of prenatal exposure to deltamethrin on cerebral and hepatic cytochrome P450s and behavioral activity in rat offspring. European Journal of Pharmacology, (2006); 544(1–3): 58–68.
- Richardson JR, Taylor MM, Shalat SL, Guillot 3rd TS, Caudle WM, Hossain MM, Mathews TA, Jones SR, Cory-Slechta DA, Miller GW. Developmental pesticide exposure reproduces features of attention deficit hyperactivity disorder. FASEB journal : official publication of the Federation of American Societies for Experimental Biology, (2015); 29(5): 1960–1972.
- Kung TS, Richardson JR, Cooper KR, White LA. Developmental Deltamethrin Exposure Causes Persistent Changes in Dopaminergic Gene Expression, Neurochemistry, and Locomotor Activity in Zebrafish. Toxicological sciences : an official journal of the Society of Toxicology, (2015); 146(2): 235–243.
- Liu G, Ma Q, Shi N. Tyrosine hydroxylase as a target for deltamethrin in the nigrostriatal dopaminergic pathway. Biomedical and Environmental Sciences, (2006); 19(1): 27.
- Tayebati SK, Di Tullio MA, Ricci A, Amenta F. Influence of dermal exposure to the pyrethroid insecticide deltamethrin on rat brain microanatomy and cholinergic/dopaminergic neurochemistry. Brain Research, (2009); 1301: 180–188.
- Robertson HA. Synergistic Interactions of D1- and D2-Selective Dopamine Agonists in Animal Models for Parkinson’s Disease: Sites of Action and Implications for the Pathogenesis of Dyskinesias. Canadian Journal of Neurological Sciences / Journal Canadien des Sciences Neurologiques, (1992); 19(S1): 147–152.
- Hasbi A, O’Dowd BF, George SR. Dopamine D1-D2 receptor heteromer signaling pathway in the brain: emerging physiological relevance. Molecular brain, (2011); 4: 26.
- Khan AM, Raina R, Dubey N, Verma PK. Effect of deltamethrin and fluoride co-exposure on the brain antioxidant status and cholinesterase activity in Wistar rats. Drug and Chemical Toxicology, (2017); 41(2): 123–127.
- Soderlund DM, Clark JM, Sheets LP, Mullin LS, Piccirillo VJ, Sargent D, Stevens JT, Weiner ML. Mechanisms of pyrethroid neurotoxicity: implications for cumulative risk assessment. Toxicology, (2002); 171(1): 3–59.
- Adefegha SA, Omojokun OS, Oboh G, Fasakin O, Ogunsuyi O. Modulatory Effects of Ferulic Acid on Cadmium-Induced Brain Damage. Journal of Evidence-Based Complementary & Alternative Medicine, (2016); 21(4): NP56–NP61.
- Abdel-Daim MM, Abushouk AI, Bungău SG, Bin-Jumah M, El-kott AF, Shati AA, Aleya L, Alkahtani S. Protective effects of thymoquinone and diallyl sulphide against malathion-induced toxicity in rats. Environmental Science and Pollution Research, (2020); 27(10): 10228–10235.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0
![]()