

Full Length Research Article
Prospects for the Therapeutic Potential of Inhibitors Targeting JNK and p53 in Alzheimer's Disease
Gleb Zyuz`kov, Larisa Miroshnichenko, Alexander Chayikovskyi, Larisa Kotlovskaya
Adv. life sci., vol. 10, no. 2, pp. 298-303, June 2023
*– Corresponding Author: Gleb Zyuz`kov (zgn@pharmso.ru )
Authors' Affiliations
[Date Received: 14/03/2023; Date Revised: 02/05/2023; Date Published Online: 30/06/2023; Date Updated: 11/09/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: It is promising to search for fundamentally new approaches to the treatment of Alzheimer's disease (AD) within the framework of creating methods for stimulating neurogenesis and developing a strategy for targeted pharmacological regulation of intracellular signal transduction in regenerative-competent cells (RCCs). Of particular interest in this case are JNK and p53. The purpose of the work was to investigate the possibility of regulating the RCCs functions using JNK and p53 inhibitors in the modelling β-amyloid-induced neurodegeneration (βAIN) in vitro.
Methods: The studies were performed using C57BL/6 mice. To model βAIN in vitro, the 25-35 fragment Amyloid β(βA) was used. The effect of the JNK inhibitors (SP600125) and p53 (Pifithrin-a, Cyclic) on the functioning of different types of progenitors and glial cells of the subventricular zone of the cerebral hemispheres (SVZ)was studied. NPCs, astrocytes, oligodendrocytes, and microglial cells were isolated from SVZ cells using immunomagnetic separation.
Results: We found the ability of the JNK inhibitor to stimulate the proliferation of NSCs and NPCs in βAIN simulation. While the p53 inhibitor is characterized by stimulatory activity in relation to the proliferation of only committed neuronal precursors under conditions of neurotoxic exposure to βA. In addition, inhibition of JNK and p53 resulted in stimulation of the secretion of neurotrophins by oligodendrocytes and microglial cells in the in vitro βAIN model.
Conclusion: The results indicate the potential ability of the JNK inhibitor to most consistently stimulate the implementation of the proregenerative properties of progenitor and neuroglial cells in AD.
Keywords: Alzheimer's disease; Neural stem cells; β-amyloid; JNK, p53
Introduction![]()
Alzheimer's disease (AD) is a common severe neurodegenerative disease with an unfavorable outcome [1, 2]. The currently used drug approaches to AD therapy based on the protection and/or modulation of the functions of mature nerve cells are unable not only to restore the morphofunctional state of the CNS, but also to stop the progression of the disease [1–4].
Despite the unknown etiology of this disease [1, 5], many aspects of its pathogenesis have been studied. For example, there is no doubt about the critically important role of the toxic effect of pathogenic fragments of β-amyloid (βA) on cells of the nervous tissue [3-6]. Moreover, it has been experimentally proven that the dysfunction of neurons and their death in conditions of amyloid aggregate formation occurs against a backdrop of loss of plasticity of the nervous tissue and the capability of neurogenesis [1, 3, 7].
At the same time, our earlier data showed that impairment of neurogenesis processes in AD develops as a result of uncoupling of mitoses and specialization of neural stem cells (NSCs) and desynchronization of their activity with neuronal-committed precursors (NCPs), as well as with the neurotrophin-producing function of glial cells [3, 7-9].
Based on this, it is promising to develop pharmacological methods of AD therapy principled of regulating the functions of regenerative-competent cells (RCCs) of the nervous tissue and coordinating the functions of their individual representatives (NSCs, NCPs, neuroglial cells) [3, 7-10]. The realization of this approach looks favorable within the context of the strategy for targeted regulation of intracellular signaling in RCCs [11-14]. The selectivity of action in this case depends on the characteristics of intracellular signal transduction in heterogeneous tissue-specific progenitors and/or particular expression of certain types and isoforms of signaling molecules in cells of different tissues [11, 15]. Therefore, one of the key stages in the development of AD treatments within the framework of this concept of pharmacotherapy is the identification of intracellular signaling molecules that can fundamentally change the functioning of certain RCCs.
JNK and p53 are known to be concerned with regulating proliferation and differentiation nervous tissue progenitors, together with the humoral function of neuroglial cells, including neurodegeneration caused by alcohol abuse [12, 16]. However, the function of these molecules in the NSCs and NCPs functioning, and neuroglial cells indirectly involved in neurogenesis in AD is still not known.
The purpose of the work was to investigate the possibility of regulating the RCCs functions (NSCs, NCPs, astrocytes, oligodendrocytes, microglial cells) using JNK and p53 inhibitors in the simulation βA-induced neurodegeneration (βAIN) in vitro.
Methods![]()
Chemicals and Drugs
Amyloid β 25-35 (Sigma-Aldrich, Germany); JNK inhibitor (SP600125, InvivoGen, USA); p53 inhibitor (Pifithrin-a, Cyclic, Santa Cruz Biotechnology, Inc. USA); basic culture medium for nerve cells MACS Neuro Medium; reagents for magnetic immunoseparation(anti-PSA-NCAM MicroBeads; anti-ACSA-2 MicroBead Kit; Anti-O4 MicroBeads; Anti-CD11b MicroBeads) (all produced by MiltenyiBiotec, Germany); hydroxycarbamidehydurea (Calbiochem, USA); DMSO (Sigma-Aldrich, USA); Primaria Cell Culture Plate (size 96 well) (Corning, USA); ethanol (Kemerovo Pharmaceutical Factory LLC, Russia).
Animals and experimental design
The research work has been carried out in respect of the principles of the human experimental technique (Directive 2010/63/EU of the European parliament and of the council on the protection of animals used for scientific purposes). The study was approved by the Institute`s local Ethics Committee (protocol GRIPh& RM-2022-01/12). The studies were performed on C57BL/6 male mice at the age of 2 months and were approved by the local ethical committee of the Institute (protocol GRIPh&RM-2022-01/12). The experiments took place in the autumn and winter. The mice were sacrificed by decapitation under deep anesthesia in a CO2 chamber (M1-TSFM-1, EZ Systems Inc., USA).
In vitro experiments, using cultural methods, studied the impact of the JNK (10 μM) and p53 (5 μM) inhibitors on the realization of the potential for growth of progenitors (NSCs, NCP) and the production of neurotrophins (growth factors) by astrocytes, oligodendrocytes, microglial cells in βAIN [3, 11, 14]. The corresponding cultures of cells with βA without inhibitors served as controls.
βAIN Modeling
To model neurodegeneration in vitro, the βA fragment 25-35 (Amyloid β-Protein Fragment 25-35, Sigma-Aldrich, USA) was used. The neurotoxic agent at a concentration of 1 mM was incubated for 7 days in the CO2 incubator (Thermo Scientific 8000 DH, Thermo Fisher Scientific Inc., USA) for aggregating proteins. Final concentration of aggregated 25-35 βA in the vitro culture medium was 20 µM [3, 11, 14].
Progenitor Cells Study
Nervous tissue to produce cellular material was isolated from the subventricular zone of the brain hemispheres (SVZ). NSCs were studied by culturing unfractionated cells from SVZ. To study NCP, CD56+(PSA-NCAM+) cells were used, which were isolated with the MiniMACS Cell Separator (MiltenyiBiotec, Germany) and the corresponding MicroBead (by positive selection method) [11, 13, 15]. Both types of cells were incubated in MACS Neuro Medium at a concentration of 105 / ml for 5 days in the incubator (Thermo Scientific 8000 DH, Thermo Fisher Scientific Inc., USA) under standard conditions (at 5% CO2 and 37°C). In cell cultures, the number of clonogenic cells, their mitotic activity, and specialization(differentiation/maturation) intensity were counted. The number of NSCs and NCPs was recorded by the number of colony forming units (CFU, neurospheres of 100 or more cells) in the corresponding cell cultures. The progenitor mitotic activity (mitotic index) was valued at using hydroxyurea, which inhibits DNA synthesis in the synthetic phase of mitosis (at a concentration of 1 μM).The intensity of cell specialization was determined based on the ratio of cluster-forming units (ClFU, neurospheres of 100-30 cells) to CFU (specialization index) [11, 13, 15].
Neuroglial Cells Study
The cells of astro-, oligodendro-, and microglia were isolated from SVZ cells using the MiniMACS Cell Separator and anti-ACSA-2, anti-O4, and anti-CD11b MicroBeads, respectively. The cells were incubated in basic culture medium in the Thermo Scientific 8000 DH (Thermo Fisher Scientific Inc., USA) (for 2 days at the concentration of 2×106 / ml) to obtain supernatants. The neurotrophin(a complex of all of active humoral factors against CFU)production was studied by the effect of supernatants on the neurospheres formation (neurosphere stimulating activity, NSA). The test system was a culture of intact of the SVZ cells (105 / ml) [11, 17].
Statistical analysis
The results were analyzed with Mann-Whitney test for independent samples. The data are given as arithmetical means, the significance level p < 0.05 [18].
Results![]()
Influence of βA on the Progenitor Functioning
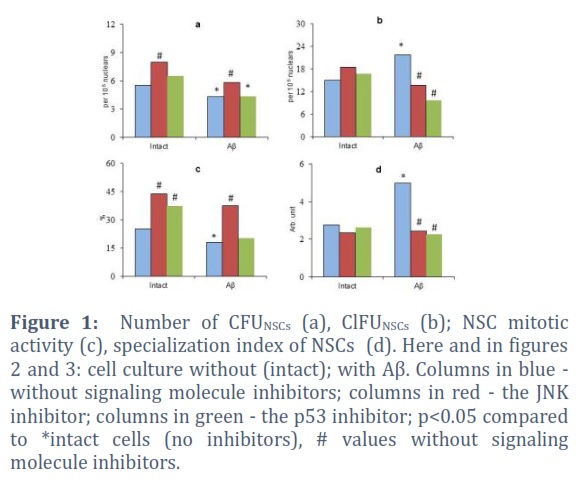
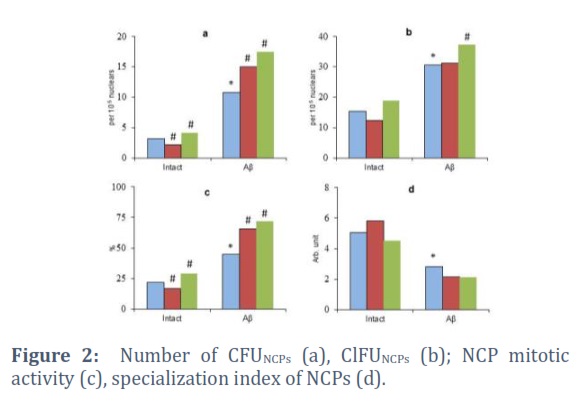
The fragment 25-35 βA, when introduced into the cell culture, caused a reduction in the performance of CFUNSСs (up to 77.3% of the initial level) and their proliferative activity (Figure 1a, c). Also, there was a significant growth in the number of ClFUNSСs in the culture medium with βA (up to 140.3% of the same parameter in the βA-free medium) and an acceleration of the NSC specialization (up to 180.2% of the same indicator in cell culture without βA) (Figure 1b, d). At the same time, Aβ stimulated the clonogenic activity of CD56+ cells. The number of CFUNСPs and their mitosis increased to 342.3% and 201.3% of the controls, respectively (Figure2a, c). The specialization index of committed precursors, on the contrary, significantly decreased (to 54.8% of the same indicator in the control) (Figure2b, d).
The results are consistent with data on the uncoupling of the NSC and NCP functioning under the influence of neurotoxic Aβ fragments [3, 7-9]. At the same time, an excessively high rate of the NSCs maturation, which can cause aberrant cell development, should be considered an important manifestation of inadaptation of the system of cell renewal of the nervous tissue (in addition to changes in the mitotic activity of progenitors and uncoupling of proliferation and differentiation of NCPs) [7, 9, 19].
Influence of βA on the secretory function of neuroglial cells
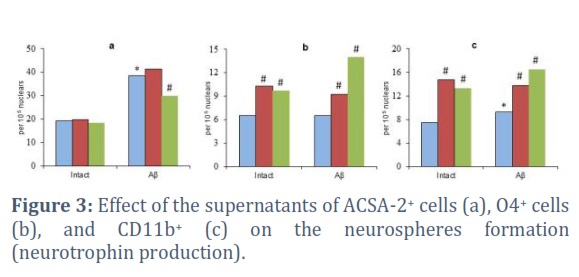
The βA effect on neuroglial cells led to an increase in the secretion of neurotrophins by ACSA-2+ and CD11b+ cells (up to 202.4% and 126.8% of the corresponding control levels) (Figure 3a, c). The NSA of supernatants from O4+ cells did not change under the influence of βA(Figure 3b).
When analyzing these data, it should be taken into account that changes in NSA were recorded by changes in colony formation under the influence of conditioned media of neuroglial cells. This is an integrative indicator reflecting the secretion of both cytokines that stimulate the functions of progenitors and the production of factors that inhibit their functions (primarily pro-inflammatory cytokines) [14, 20, 21]. Moreover, the contribution of the production of inhibitors of the functioning of progenitor cells to colony formation can be very large. It is known that aggregated amyloid initiates an excessive inflammatory reaction, which plays the role of a pathogenic factor in AD[4, 21].
Influence of the signaling molecules inhibitors on NSC functioning
When studying the effect of the JNK and p53 inhibitors on the achieving growth potential of multipotent NSCs, some interesting phenomena were revealed. The JNK blockade in unfractionated cells in a medium without βA has resulted in the increase of the CFUNSСs number in the unfractionated cells culture (up to 145.5% of the initial level) and their mitotic activity (up to 174.1% of the initial value). While the p53 inhibitor had no significant effect on these parameters. Changes in ClFUNSСs and the intensity of NSC specialization were not recorded in both cases (upon inactivation of both JNK and p53) (Figure 1).
In many respects, a different picture was observed when unfractionated cells were cultivated in the presence of βA. In this case, the stimulating effects on NSC proliferation remained only in the JNK inhibitor (up to 209.3% of the control, a similar parameter in the medium with βA without inhibitors of signaling molecules). This naturally led to an increase in the number of CFUNSСs (up to 135.6% of the control level). At the same time, both inhibitors significantly reduced the amount of ClFU in the culture of unfractionated cells (Figure 1b). In both cases, a decrease in the rate of NSC differentiation was also recorded (up to 48.4% and 45.4% of the control level with JNK and p53 blockade, respectively). Moreover, this should be considered as a potential factor for increasing the efficiency of neurogenesis in βAIN, which consists in preventing aberrant cell development due to the excessive intensity of progenitor differentiation [10, 14].
Thus, only the selective blockade of JNK stimulated the cell cycle progression of multipotent NSCs. At the same time, inactivation of both studied signaling molecules was accompanied by inhibition of excessive activation of the specialization of these cells [7, 9, 19].
Influence of the signaling molecules inhibitors on NCP functioning
The investigation of the JNK and p53 inhibitors effect on the functioning of neuronal precursors revealed other patterns. The JNK blockade in conditions of optimum vital activity of NPCs reduced the level of their proliferation (up to 80.4% of the control) and the clonogenic capacity of the CD56+ cells (up to 68.5% of the control of the initial level) (Figure 2a, c). While the p53 inactivation under these conditions was accompanied, on the contrary, by stimulation of the NCP mitosis and the release of CFUNCPs (up to 133.8% and 131.5% of the control levels, respectively).
When βAIN modeling, NPCs reacted differently to the targeted regulation of intracellular signaling. When cultivating CD56+ cells in the βA presence, both inhibitors increased the number of CFU (up to 138.5% and 161.6% of the control values in the JNK and p53 inhibition, respectively) and their mitotic activity (up to 145.6% and 160.4% from control values for blockade of JNK and p53, respectively). No significant changes were recorded in terms of the cluster formation and maturation intensity in both cases (as well as in the βA-free medium) (Figure 2b, d).
Thus, the response of committed neuronal precursors to the JNK inhibitor depended on the initial conditions of their vital activity. But in βAIN, this pharmacological agent significantly stimulated their proliferation. In contrast, the blockade of p53 increased the mitotic activity of NCPs regardless of external conditions.
Influence of the signaling molecules inhibitors on the functioning of neuroglial cells
During the experiments, an ambiguous effect of selective the JNK and p53 blockers on implementing of the secretory function of neuroglial cells was revealed. The inactivation of these signaling molecules in astrocytes had practically no effect on the production of neurotrophic growth factors. The only exception was the p53 inhibition in βAIN modeling. In this case, a decrease in the parameter was observed (up to 77.9% of the control) (Figure 3a).
The inactivation of JNK and p53 in O4+ and CD11b+ cells was together with a pronounced increase in the supernatant NSA in both cases (cell cultivation with and without βA). The increase in NSA in the test system when using conditioned media obtained by incubation of oligodendrocytes and microglial cells under intact conditions reached 157.7% and 196.7% with the JNK blockade and 150.0% and 176.7% with the p53 inhibition(from appropriate control levels) (Figure 3b, c).
Thus, targeted blockade of JNK and p53 stimulated the secretory function of oligodendrocytes and microglial cells, which naturally activated the activity of progenitors, including under βAIN conditions.
Figures & Tables
Discussion![]()
The results confirm the development of the phenomenon of discoordination in the activity of multipotent and committed progenitor neurons under the influence of βA [3, 7-10]. Inhibition of the mitotic activity of NSCs was revealed against the background of an increase in that of NSCs. The specialization intensity of multipotent NSCs increased, while in neuronal precursors, on the contrary, it decreased. Thus, desynchronization of the proliferative activity of NSCs and NPCs developed in the context of de-coupling proliferation and differentiation of both types of progenitors. Moreover, the revealed very rapid intensity of the differentiation/maturation of NSCs could be one of the factors of the aberrant pathway of development of nerve cells in AD [7, 9, 19]. Such dysfunctions in the functioning of the system of cellular renewal of the nervous tissue cannot but affect the efficiency of neurogenesis in AD [3, 9, 22]. Moreover, the influence of phosphorylated tau proteins, as well as cholinergic changes in the brain and other disturbances in tissue homeostasis in AD [23-25] will exacerbate and multiply progenitor dysfunction in AD in situ.Therefore, the most promising approaches to stimulate neurogenesis in AD should be the development of pharmacological methods for synchronous stimulation of the activity of RCCs of various types [7, 9].
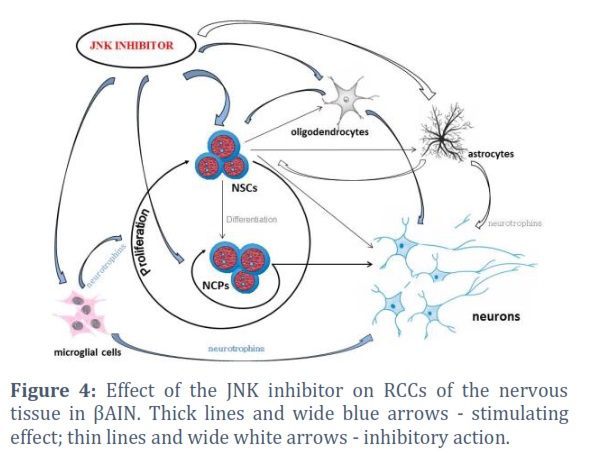
At the same time, it was found that under βAIN conditions, the most coordinated activity of RCCs of the nervous tissue is observed during JNK blockade (Figure 4). Such targeted regulation of intracellular signaling can increase the achievement of growth potential of both committed precursors and NSCs (in contrast to the use of the p53 inhibitor). It should be noted that inhibition of both JNK and p53 was accompanied by inhibition of excessive activation of NSC specialization processes [7, 9]. That is, the p53 inhibitor also (as well as the JNK blocker) affected the neurogenesis disadaptation factor in βAIN, which prevented the aberrant development of nerve cells [8, 9, 19]. In addition, experiments have shown that targeting JNK and p53 can significantly stimulate the neurotrophin-producing function of oligodendrocytes and microglial cells (and also, possibly, suppress the production of neuroinflammation-inducing factors secreted by the latter in AD) [20, 21]. The negative effect on the feeder activity of neuroglia was found only in the p53 inhibitor, and only in relation to astrocytes. Although how important this may be requires further study.
The findings indicate the prospects for the development of new approaches to stimulate neurogenesis in AD using JNK and p53 inhibitors. However, the most preferred target in Alzheimer's drug discovery is JNK targeting.
Conflict of Interest
The authors state that there is no conflict of interest here.
Acknowledgement
We thank the directorate of the Tomsk National Research Medical Center and the director of the Goldberg Research Institute of Pharmacology and Regenerative Medicine V.V. Zhdanov personally for providing the infrastructure for work.
Financial Support
The investigation was conducted with with a grant from Russian Science Foundation No. 22-25-00069, https://rscf.ru/project/22-25-00069/.
![]()
References
- Lei P, Ayton S, Bush AI. The essential elements of Alzheimer's disease. The Journal of biological chemistry, (2021); 296: 100105.
- Scheltens P, De Strooper B, Kivipelto M, Holstege H, Chételat G, et al. Lancet, (2021); 397(10284): 1577-1590.
- Zyuz’kov GN, Miroshnichenko LA, Chayikovskyi AV, Kotlovskaya LY. Functional State of Various Types of Regenerative Competent Neural Tissue Cells in β-Amyloid-Induced Neurodegeneration. Bulletin of experimental biology and medicine, (2022); 173(6): 709-713.
- Guzman-Martinez L, Calfío C, Farias GA, Vilches C, Prieto R, et al. New Frontiers in the Prevention, Diagnosis, and Treatment of Alzheimer's Disease. Journal of Alzheimer's disease, (2021); 82(s1): S51-S63.
- Ogbodo JO, Agbo CP, Njoku UO, Ogugofor MO, Egba SI, et al. Alzheimer's Disease: Pathogenesis and Therapeutic Interventions. Current aging science, (2022); 15(1): 2-25.
- Weaver DF. β-Amyloid is an Immunopeptide and Alzheimer's is an Autoimmune Disease. Current Alzheimer research, (2021); 18(11): 849-857.
- Zyuz’kov GN, Miroshnichenko LA, Chayikovskyi AV, Kotlovskaya LY. NF-кB: a target for synchronizing the functioning nervous tissue progenitors of different types in Alzheimer's disease. Current molecular pharmacology, (2023); 16(2): 234-241.
- Zyuz’kov GN, Miroshnichenko LA, , Kotlovskaya LY, Chayikovskyi AV. Prospect of Using ERK1/2 and р38 in Regeneration-Competent Cells of Nervous Tissue as a Drug Targets for Treating Alzheimer's Disease. Biointerface research in applied chemistry, (2023); 13(2): 1-9.
- Zyuz’kov GN, Miroshnichenko LA, , Kotlovskaya LY, Chayikovskyi AV. Inhibitors of Intracellular Signaling Molecules: New Horizons in Drug Discovery for the Treatment of Alzheimer's Disease. Biointerface research in applied chemistry, (2023); 13(5): 1–11.
- Zyuz’kov GN, Miroshnichenko LA, Chayikovskyi AV, Kotlovskaya LY. The Role of MARK ERK1/2 and p38 in Regulation of Functions of Neural Stem Cells and Neuroglia under Conditions of β-Amyloid-Induced Neurodegeneration. Bulletin of experimental biology and medicine, (2022); 173(4): 424-428.
- Zyuz’kov GN. Targeted Regulation of Intracellular Signal Transduction in Regeneration-Competent Cells: A new Direction for Therapy in Regenerative Medicine. Biointerface research in applied chemistry, (2021); 11(4): 12238-12251.
- Zyuz’kov GN, Miroshnichenko LA, Polyakova TYu, Stavrova LA, Simanina EV. JNK and p53 inhibition in regenerationcompetent cells of nerve tissue: a novel approach for treatment of ethanol-induced neurodegeneration. ACTA Pharmaceutica Sciencia, (2023); 61(1): 49-63.
- Zyuz’kov GN, Miroshnichenko LA, Polyakova TYu, Simanina EV. Neuroprotective and Neuroregenerative Effects of Shikonin-mediated Inhibition of NF-κB/Stat3 in Alcoholic Encephalopathy. Letters in drug design and discovery, (2023); 20: E-pub Ahead of Print (Published on: 07 December, 2022)
- Zyuz’kov GN, Miroshnichenko LA, Polyakova TYu, Simanina EV, Stavrova LA. Targeting cAMP-pathway in Regeneration-Competent Cells of Nervous Tissue: Potential to Create a Novel Drug for Treatment of Ethanol-Induced Neurodegeneration. Central nervous system agents in medicinal chemistry, (2021); 21(3): 172-180.
- Zyuz’kov GN, Miroshnichenko LA, Polyakova TYu, Zhdanov VV, Simanina EV, et al. Specific Features of Intracellular Signal Transduction in the Regulation of Functions of Neural Stem Cells and Committed Neuronal Progenitors. Bulletin of experimental biology and medicine, (2021); 170(4): 522-527.
- Zyuz’kov GN, Bryushinina OS, Zyuz'kova YuG, Lakeev AP, et al. Targeting JNK as a Novel Approach to Drug Metabolism Regulation. Biointerface research in applied chemistry, (2022); 12(6): 7596-7605.
- Zyuz’kov GN, Zhdanov VV, Miroshnichenko LA, Polyakova TYu, Simanina EV, et al. The Role of JAK and STAT3 in Regulation of Secretory Function of Neuroglial Cells of Different Types in Ethanol-Induced Neurodegeneration. Bulletin of experimental biology and medicine, (2022); 172(6): 686-690.
- Curtis MJ, Bond RA, Spina D, Ahluwalia A, Alexander SP, et al. Experimental design and analysis and theirreporting: New guidance for publication in BJP. British journal of pharmacology, (2015); 172(14): 3461-3471.
- Bryson BL, Tamagno I, Taylor SE, Parameswaran N, Chernosky NM, et al. Aberrant Induction of a Mesenchymal/Stem Cell Program Engages Senescence in Normal Mammary Epithelial Cells. Molecular cancer research: MCR, (2021); 19(4): 651-666.
- Siddiqui A, Akhtar S, Shah Z, Othman I, Kumari Y. Inflammation Drives Alzheimer's Disease: Emphasis on 5-lipoxygenase Pathways. Current neuropharmacology, (2021); 19(6): 885-895.
- Lamie PF, Abdel-Fattah MM, Philoppes JN. Design and synthesis of new indole drug candidates to treat Alzheimer's disease and targeting neuro-inflammation using a multi-target-directed ligand (MTDL) strategy. Journal of enzyme inhibition and medicinal chemistry, (2022); 37(1): 2660-2678.
- Babcock KR, Page JS, Fallon JR, Webb AE. Adult Hippocampal Neurogenesis in Aging and Alzheimer's Disease. Stem Cell Reports, (2021); 16(4): 681-693.
- Wegmann S, Biernat J, Mandelkow E. A current view on Tau protein phosphorylation in Alzheimer's disease. Current opinion in neurobiology, (2021); 69: 131-138.
- Downey J, Lam JCK, Li VOK, Gozes I. Somatic Mutations and Alzheimer's Disease. Journal of Alzheimer's disease, (2022); 90(2): 475-493.
- Giacobini E, Cuello AC, Fisher A. Reimagining cholinergic therapy for Alzheimer's disease. Brain, (2022); 145(7): 2250-2275.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0
![]()