

Full Length Research Article
Isolation of agar-degrading Bacillus sp. AT6 and preliminary application for seaweed saccharification
Dang Thi Thanh Ha1, Trinh Thi Phuong Thao2, Le Thi Kim Thoa2, Tran Trung Dung1, Tran Thi Minh Ha1, Phan Thi Thuc Quyen1, Bui Thi Khuyen3, Nguyen Duc Huy3*
Adv. life sci., vol. 10, no. 3, pp. 390-397, September 2023
*– Corresponding Author: Nguyen DucHuy (ndhuy@hueuni.edu.vn)
Authors' Affiliations
2. Jeonbuk National University, Jeonju-si, Jeollabuk-do, 54896 – Republic of Korea
3. Institute of Biotechnology, Hue University, Hue – Vietnam
[Date Received: 05/03/2023; Date Revised: 15/06/2023; Date Published Online: 30/09/2023]
Editorial Note on Version of Record
26 June 2025: This article has been corrected. See https://doi.org/10.62940/als.v13i0.4221 for more information.
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Agar is a common polysaccharide found in nature. However, agar is strongly resistant to the degradation processing, leading to limitation of its application in various areas. Thus, finding an effective solution for agar saccharification significantly improves the economically effects of agar based substrates.
Methods: Soil samples were collected from TienPhong Forestry Ltd. Company, ThuyXuan District, ThuaThien Hue province, Vietnam. Potential agar degrading bacteria were screened on a mineral salt agar medium. The isolate was identified based on 16S rRNA nucleotide sequence, morphological, physiological, and biochemical characteristics. Agarase production was evaluated by modification culture conditions including incubation time, shaking speed, and initial inoculum size. Molecular mass of extracellular agarase was determined by zymogram analysis following SDS-PAGE. The effect of pH, temperature, metal ions, and organic solvents were performed to characterize the enzyme. Application of enzyme was investigated on seaweed saccharification.
Results: An agar degrading bacterial strain was isolated from soils and identified as Bacillus sp. AT6. Maximal agarase accumulation was obtained in the culture containing an inoculum size of 10% (v/v), shaking speed of 210 rpm, and 96 hours incubation. The agarase revealed a single band on zymogram analysis with an apparent molecular weight of 180 kDa. The optimal temperature and pH were 40°Cand pH 8.0, respectively. All tested metal ions and organic solvents partially decreased enzyme activity. Treating seaweed with agarase resulted in reducing sugars release present in the reaction, indicating the saccharification of seaweed was successful.
Conclusion: Bacillus sp. AT6 is a new report of agarolytic bacteria that produces extracellular agarase enzymes. The present results promise strain AT6 is a great candidate for agar saccharification for industrial application.
Keywords: Agar degrading bacterium; Agarase; Hydrolysis, hydrolyzed products; Soil bacteria
Introduction![]()
Agars are the most important component contributing to cell walls structure of genera Gelidium and Gracilaria red algae. The Gracilaria genus plays economic importance as the most abundant and promising resource for agar production. This alga is commercially cultivated in various countries in subtropical regions such as Indonesia, Argentina, South Korea, Vietnam, Philippines, Chile, and Namibia [1]. Agars consist of the matrix between agaroses and agaropectin backbones, accounting for 70% and 30% dry weight, respectively [2,3]. Agarose is linked by backbones of (1→3)-β-D-galactose and (1→4)-3,6-anhydro-α-L-galactose units. Meanwhile, agaropectin polysaccharide chain consists of 3,6-anhydro-l-galactose units in integration with pyruvate and sulfoxy/methoxy residues [4]. Agars have been widely applied in food, pharmaceutical, cosmetics industries [5]. Basically, agar structure can be degraded by different methods including enzymatic saccharification and chemical hydrolysis. However, enzymatic saccharification is a preferable strategy for agar-oligosaccharides preparation due to high efficiency, less contamination and environmentally friendly [6].
Agarases (EC 3.2.1.81) belong to the glycoside hydrolase (GH) family which catalyze the hydrolysis of agar. According to the distinct cleavage pattern or production formation, agarases contain α-agarase (E.C. 3.2.1.158) and β-agarase (E.C. 3.2.1.81). α-agarase acts on α‐1,3‐linkages, whereas β-agarase catalyzes β‐1,4‐linkages, resulting in agaro-oligosaccharides and neoagaro-oligosaccharides products, respectively [7-9]. Agarases and their hydrolysis products have extensive applications in the cosmetic, pharmaceutical and food industries [10]. These agaro-oligosaccharides exhibit a great physiological activities including moisturizing effect [11], skin whitening effects, anti-inflammatory [12], macrophage-stimulating, antioxidant, and immunogenic activities [13], anti-diabetic and anti-obesity effects [14], and can be used as prebiotics [11].
Up to date, most agar degrading bacteria have been reported living in marine habitats such as marine sediments, marine algae, and seawater including Vibrio sp. [15], Ammoniibacillus sp. [16], Acinetobacter junii [17], and Flammeovirga sp. [18]. However, a few studies have reported several species of agarase enzyme-producing bacteria are deriving from freshwater and terrestrial environments [19]. In this study, we investigated the agarolytic bacteria isolated from soils in local forestry farm. The effect of culture conditions on agarases production and biochemical properties of extracellular agarases by isolate was also evaluated. Finally, the isolate was further applied for seaweed (Gracilaria verrucosa) hydrolysis.
Methods![]()
Sample collection
Soil samples were collected from TienPhong Forestry Ltd. Company, ThuyXuan District, ThuaThien Hue province, Vietnam and stored in plastic bags. Then, the samples were delivered and stored at 4°Cin laboratory for further analysis.
Screening and isolation agarase producing bacteria
One gram of soil was suspended in distilled water with ratio 1:10 and diluted by 10-fold serial dilution method to obtain 10−4, 10−5, and 10−6 diluted solutions. Samples (0.1 mL) of each dilution were then plated on Mineral Salt Agar (MSA) containing (g/L): K2HPO4 0.5; (NH4)2SO4 0.5; MgSO4.7H2O 0.5; CaCl2 0.1; NaCl 0.5; agar 15 and pH 7.0. Plates were inoculated at 37°C for 7 days and daily quantified for agarolytic activity. The depression or liquefaction around colonies was assessed as a signal of agarase activity. After 7 days, plates were stained using iodine solution. The appearance of pale-yellow halo zones was referred to as agar-degrading activity [20]. All colonies showing liquefaction, depressions and clearance zone in the agar were picked up, transferred to new LB medium plates and purified by repeating sub-culture on new LB medium. The colony with the highest potential agarolytic activity was selected for further investigation.
Identification of the bacterial isolates
The isolate was identified morphological characteristics including shape, colour, transparency, margin by Gram staining techniques and observed using a microscope [21]. Catalase assay was conducted by standard protocol [22].
The total DNA of isolate was extracted based on description of Sambrook et al. (2001) [23]. The total DNA was diluted in 30 µL sterile water. The 16S rRNA nucleotide sequence was amplified using a primer pair consisting of 27-F (5’-AGAGTTTGATCCTGGCTCAG–3’) and 1492-R (5′-TACCTTGTTACGACTT-3′). The amplification was carried out in a Thermal Cycler (Eppendorf, Germany). The amplification protocol consisted of an initial denaturation step for 5 min at 95°C, 30 cycles of denaturation step for 60 s at 95°C, annealing step for 30 s at 55°C, extension step for 90 s at 72°C. The reaction was extended for 10 min at 72°C. Amplicon products were analyzed by electrophoresis on 1 % (w/v) agarose gel. The 16S rRNA nucleotide sequence was obtained by a DNA sequencing (Firstbase, Malaysia). The nucleotide sequence was analyzed to remove errors and aligned with nucleotide sequences on GenBank database using BLAST tool. Sequence similarities between the isolates and closest relatives were determined and recorded. Isolate was classified by conducting a phylogenetic analysis by Maximum Likelihood methodusing MEGAsoftware (Ver. 11) [24].
Agarase production
The single colony of isolate was inoculated in 5 mL LB medium for overnight (approximately 16 h). The overnight culture (1%, v/v) was transferred into 50 mL MSA medium containing 0.2% (w/v) agar and continuously cultured at 37°C for 120 h, shaking speed of 180 rpm. The supernatant cultures were harvested every 24 h by removing cell pellets via centrifugation at 10,000 rpm, 4°C, 15 min. Agarase activity was quantified according to the enzymatic assay [17].
Enzymatic assay
Agarase activity was determined by 3,5-dinitrosalicylic acid (DNSA) reagent procedure with some modifications [25]. One hundred microliters of the enzyme solution was mixed a volume of 400 µL containing 50 mM phosphate buffer (pH 7.0) and 0.2% (w/v) agarose [26]. The reaction was carried out at 40°C for 15 min [27]. Then, a volume of 500 μL DNSA solution was added to the reaction mixture. The reaction was terminated by boiling for 10 minutes. The reducing sugars release was measured at the absorbance of 540 nm. Finally, the reduced sugars were calculated based on D-galactose concentration. Enzyme activity was expressed as unit and was assessed as an enzyme amount required for producing 1 μmol of D-galactose per minute through agarose hydrolysis.
Effect of culture conditions on enzyme production
The culture conditions affecting on agarase production were conducted by culture the isolate on 50 mL MSA containing 0.2% (w/v) agar, pH 7.0. The culture was carried out at 37°C for 5 days. Inoculum sizes of 1, 5, and 10% were selected to evaluate agarase accumulation in medium culture. Meanwhile, shaking speeds were performed at values of 150, 180 and 210 rpm. One milliliter of culture was harvested daily and removed the cells pellet by centrifuging at 10.000 rpm, 4°C for 15 min. The supernatants were used for enzymatic quantification assay as described above.
Zymogram analysis
Zymogram analysis was carried out to estimate the molecular mass of agarase using a 5% polyacrylamide stacking gel and a 12% polyacrylamide separating gel containing 2% agar. After electrophoresis, the gel was sunk in 2% (v/v) Triton X-100 for 30 min at room temperature and washed three times using distilled water. Gel was incubated in 50 mMTris-HCl buffer (pH 8) for 4 hat 37ºC. To visualize agarase activity, the gel was stained using Lugol’s iodine solution for 10 minutes. The appearance of pale-yellow zones against a brown-violet background was assessed as agarase activity [28].
Agarase characterization
The effect of pH on enzyme activity was evaluated using different pH buffer solution. Glycine-HCl was used to generate buffers with pH 2-3, sodium acetate was prepared for pH 4-5 buffers, sodium phosphate was carried out for pH 6-8 buffers and glycine -NaOH was conducted for pH 9-12 buffers. Enzymatic assay was conducted as mentioned above. The effect of temperature was performed in a temperature ranging from 20 to 70°C. The residual activity was accessed as the activity percentage in compared to the maximum enzyme activity.
The effects of metal ions and organic solvents on enzyme activity were carried out by incubation enzyme solution in the presence of 1 mM Ca2+, Co2+, Cu2+, Mn2+, Mg2+, Zn2+ and Fe2+ ions. The enzyme solution was incubated for 1 h at 40°C, pH 8.0. Then the retained agarase activities were recorded. To evaluate the effect of organic solvents, the retained enzyme activities were determined after 1 h of incubation enzyme solution at 40°C, pH 8.0 in the presence of various chemical detergents (acetone, methanol, ethanol, hexan and isopropanol) at 30% concentration. Reactions without metal ions or organic solvents were used as control.
Evaluation of seaweed hydrolysis
The extracellular agarase was harvested after 96 h of culture in MSA and centrifuging for 15 min at 10.000 rpm, 4°C. The seaweed (G. verrucosa) was purchased in local market and grinded into powder. Seaweed powder was mixed with 50 mM of pH 8.0 phosphate buffer to final concentrations of 0.1, 0.3 and 0.5% (w/v) and incubated at 40°CThe enzyme was added and the reactions were conducted for 24 hours. The reducing sugars release was measured as described above.
Data analysis
The experiments were repeated with three replicates and the data were analyzed using MS excel software (Ver. 2016).
Results![]()
Screening and isolation agar degrading bacteria
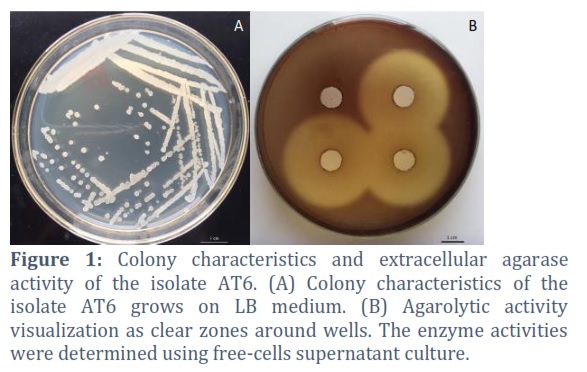
Potential agarolytic bacteria in the soils were preliminary screened MSA medium. After 24 hours inoculation, potential agarolytic bacteria grew on surface of agar plate and formed a clearing liquefaction and depression zone around colonies. Total of 7 isolates showing different morphology were transferred into new MSA agar plate. Figure 1 showed the agar hydrolytic by supernatant culture of the isolate AT6, indicating a clearance zone with diameter of 39 ± 1 mm after 24 h of incubation at 37°C. Thus, the isolate was selected for further investigation on agarolytic activity.
Bacteria identification
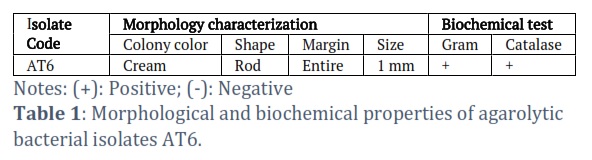
The identification of strain AT6 was based on morphological and physiological characteristics. The morphological characteristics are shown in Figure 1A and Table 1. AT6 isolate formed cream colonies and ranged in size 1.0 mm. The bacterium was a rod-shaped under the microscope observation. Gram staining indicated the isolate was Gram-positive. Meanwhile, the isolate tested catalase-positive.
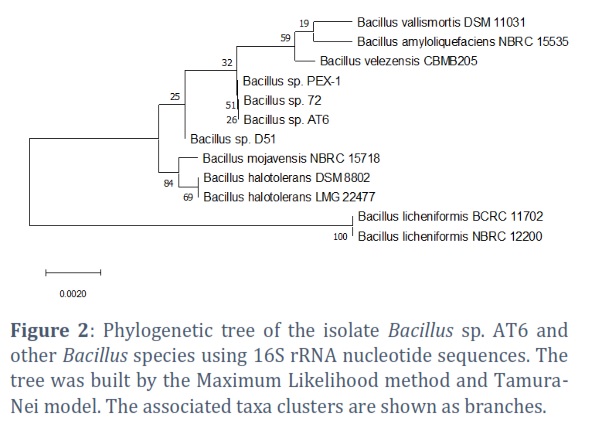
The 16S rRNA region was sequenced and the nucleotide sequence analysis clearly demonstrated that strain AT6 belongs to Bacillus genus and exhibited maximum similarity with Bacillus sp. 72 and Bacillus sp. PEX-1 (100% similarity). The phylogenetic tree between strain AT6 and other Bacillus species showing in Figure 2 confirmed the genetic relationship. Thus, the AT6 strain was identified as Bacillus sp. AT6. The nucleotide sequence has been deposited on GenBank databases under accession number of ON227004.
Agarase production
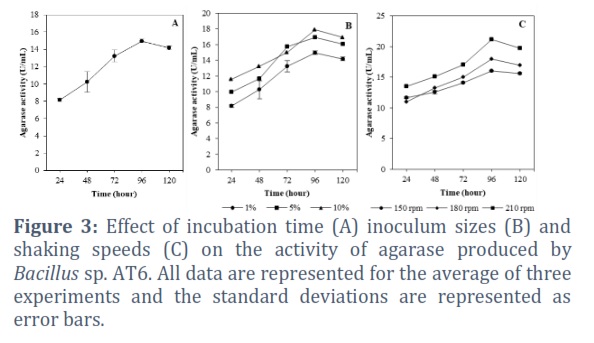
The effect of incubation time on agarase production is shown in Figure 3A. Particularly, the enzyme activity gradually increased after 24 hours of incubation (8.17 U/mL) and reached maximal activity (14.96 U/mL) at 96 hours of incubation. Extension culture after this optimum period did not increase the activity of the enzyme, whereas the activity dropped to 14.15 U/mL after 120 hours of incubation. Meanwhile, agarase enzyme activities of Bacillus sp. AT6 at different inoculum sizes are shown in Figure 3B. The results demonstrated that the size of the inoculum had a strong relation with enzyme activity. The lowest agarase activity was observed with a 1% (v/v) inoculum, while the highest activity (17.92 U/mL) was achieved with a 10% (v/v) inoculum (Figure 3B). The highest enzyme activity (17.92 U/mL) observed at 10% (v/v) inoculum size.
Oxygen is an essential factor for the growth of aerobic bacteria and typically depends on the shaking speed of the culture process. The effect of shaking speed on agarase accumulation was examined by varying the shaking speed from 150 to 210 rpm. Maximal agarase activity of 21.15 ± 0.23 U/mL obtained when the shaking speed was 210 rpm. The agarase activity reached 16.03 ± 0.13 and 17.93 ± 0.39 U/mL at 150 and 180 rpm, respectively (Figure 3C).
Zymogram analysis
Molecular mass and agarase activity were identified by SDS-PAGE using 12% polyacrylamide gels contained with 2% agar. Figure 4 showed an apparent clear band on a brown-violet background with molecular mass of approximately 180 kDa. This result indicated the Bacillus sp. AT6 secreted extracellular agarase at very high molecular mass.
Characterization of agarase
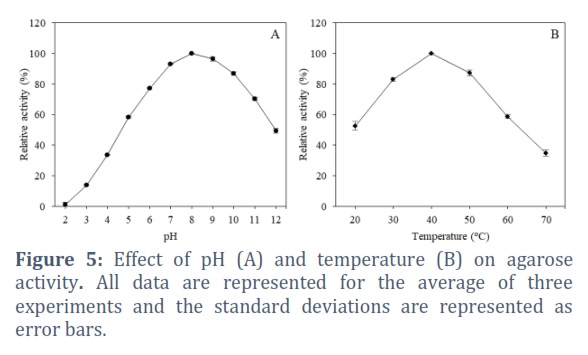
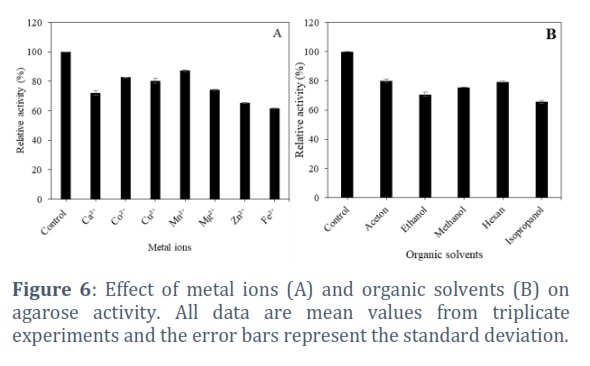
The effect of pH on agarase activity was investigated at different pH from 2.0 to 12.0. The obtained results in Figure 5A shows agarase from Bacillus sp. AT6 was active in wide pH ranging of 7-10 with peak activity at pH 8.0. Enzyme activity maintained 93.26%, 96.42%, and 87.05% at pH 7.0, 9.0, and 10.0, respectively. Meanwhile, the enzyme strongly inhibited in acidic conditions whereas the enzyme was strongly inhibited in acidic conditions, retaining less than 20% of its activity at pH 5.0. Thermal dependence of agarase was evaluated by quantifying the enzyme activity variation at temperatures from 20 to 70°C. The results in Figure 5B showed the influence of temperature on agarase activity secreted by Bacillus sp. AT6. The enzymatic activity gradually increased with increasing temperature up to the optimum temperature at 40⁰C and rapidly decreased at the temperature above 50°C. Effect of metal ions on agarase activity are shown in Figure 6A. Fe2+ and Zn2+ ions exhibited strong inhibitory effects on the agarase activity. In comparison, Ca2+, Co2+, Cu2+, Mn2+ and Mg2+ ions slightly exhibited a suppressive effect. Figure 6B showed the effects of different organic solvents on agarase activity, indicating the agarase activity significantly affected by organic solvents at final concentration of 30%. The enzyme activities were 80.09 ± 1.25%, 70.71 ±1.37%, 75.55 ± 0.08%, 79.35 ± 0.33%, and 65.64 ± 1.15 % in the presence of acetone, ethanol, methanol, hexan, and isopropanol, respectively compared to control.
Evaluation of seaweed hydrolysis
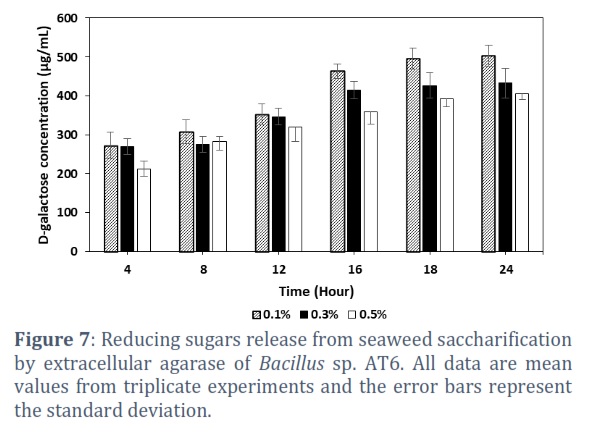
Agar is major component of seaweed. In this study, the ability of using extracellular agarase from Bacillus sp. AT6 to hydrolyze the seaweed was examined at various concentrations of 0.1, 0.3, and 0.5% (w/v). The results in Figure 7 indicated the highest amount of reducing sugars release was 503.02 ± 27.57 µg/mL after 24 hours of incubation at 40°C when incubated the enzyme with seaweed at concentration of 0.1%. Meanwhile, the agarase produced 432.86 ± 38.26 and 404.58 ± 13.97 µg reducing sugars at seaweed concentration of 0.3% and 0.5%, respectively.
Figures & Tables

To date, most agar- degrading bacteria have been detected in marine habitats, where red algae act as a substrate or only source of carbon. However, a few studies have focused on bacteria degrading agar from non-marine environments, such as soils. Previous studies isolated agar-degrading bacteria including Alteromonas sp. and Cytophaga sp. from freshwater; Spirochaetaalkalica from a soda lake; Paenibacillus sp. SSG-1 [29], Bacillus sp. [30], Cytophaga sp. [31]; Steroidobacter agariperforans from vegetable crop fields [32,33]; and Streptomyces coelicolor from soils [2]. In this study, a agar-degrading bacterium strain was successfully isolated and screened from soils. The molecular identification based on 16s rRNA nucleotide sequence, morphology, physiological and biochemical characteristic analysis indicated the isolate belongs to Bacillus genus.
Modification culture conditions affected on enzyme accumulation in which the optimal initial inoculum size, shaking speed, and incubation time were 10% (v/v), 210 rpm, and 96h, respectively. A smaller inoculum size necessitates more time for the cells to proliferate to a sufficient quantity to consume the substrate and produce the enzyme. A higher cells count in the optimum inoculum would assure fast division and biomass synthesis.Meanwhile, a low agitation intensity significantly impacts on enzyme accumulation by bacteria due to reducing oxygen supply and poor mixing of nutrients in medium fermentation culture [34].
Bacillus sp. AT6 produced a single extracellular agarase with molecular mass of approximately 180 kDa. The reported molecular mass of agarasesvarried from 20 to 360 kDa. Among them, the largest agarase is found in Alteromonas agarlyticus GJ1B [35], while the smallest agarase secreted by Vibrio sp. AP-2 [15]. Agarivorans sp. BK-1 produced three agar-degrading enzymes with molecular masses of 110, 90, and 55 kDa. The size of the agar-degrading enzyme with the highest activity was confirmed to be 110 kDa [36]. Meanwhile, soil bacteria including Acinetobacter sp. AGLSL-1, Bacillus sp. MK03, and Alteromonas sp. E-l produced agarase with molecular mass of 100 kDa, 113 kDa and 180 kDa, respectively [30,37,38].
The extracellular agarase exhibited maximal activity at pH 8.0 and temperate of 40°C. These findings are similar to agarase from Antarctic psychrophilic, Pseudomonas aeruginosa ZSL-2, and Thalassospira profundimonas with pH peaks at 8.0 [39,40, 41]. This optimum pH was higher compared to that of agarase from Alteromonas sp. C-1 of pH 6.5 [42], Vibrio sp. AP-2 of pH 5.5 [15]. The thermal dependent profiles of agarase from Bacillus sp. AT6 are consistent with the previous reportssuch as Alteromonas sp. E1 [38], Acinetobacter sp. AGLSL-1 [13], Microbulbifer sp. Q7 [6]. Meanwhile, agarase produced by Aquimarina agarilytica ZC1 exhibited lower optimal temperature of 25oC optimum pH at 7.0 [26]. Other studies reported higher optimal temperature for agarase including agarase from Alterococcus agarolyticusa [43], Halococcus sp. 197A [44], and Acinetobacter sp. [13]. In the present study, agarase produced by Bacillus sp. AT6 had moderate temperature optimum that suggests some advantages such as degrading the agar process at room temperature, reducing the cost of establishing conditions for enzyme activity. All metal ions and organic solvents partially inhibited enzyme activity. These effects were similar to previous report, in which Ca2+, Mg2+, Na+, Cu2+ and K+ ions slightlyinhibited the activity of β-agarase [41].
Agar products delivered from red algae have numerous applications for human. The hydrolysate products contain high bioactive activities including anti-inflammatory, antioxidant, and immunogenic activities [5]. Thus, agar hydrolysates are importance sources for various industries including cosmetic, pharmaceutical and food [45]. Moreover, the applied of microbial enzymes for agar hydrolysis process positively contributes to enhance the economic value chain of red algae. The present data demonstrated that extracellular agarase from Bacillus sp. AT6 efficiently hydrolyzed seaweed powder in which released reducing sugar reached 503.02 ± 27.57 µg/mL. However, the hydrolysis efficiency reduces when increasing the seaweed powder concentration. Thus, the present study suggests that enzymes and substrates only work optimally at an appropriate ratio; the saccharification efficiency decreased at higher substrate concentrations, suggesting the enzyme-to-substrate ratio is a critical factor. This study is enabling the application capacity of using extracellular agarase for treatment of seaweed or enriched agar substrates to higher value products.
Acknowledgment
This study was supported by the Tay Nguyen University under grant number T2022-57CBTĐ. The authors also acknowledge the partial support of Hue University under the Core Research Program, Grant No. NCM.DHH.2020.11.
Competing Interest
The authors declare that there is no conflict of interest.
Dang Thi Thanh Ha: Methodology, Investigation, Data analysis, Writing – original draft, Writing – review & editing, Visualization, Project administration. Trinh Thi Phuong Thao: Investigation, Data analysis, Writing – original draft, Writing – review & editing, Visualization. Le Thi Kim Thoa: Validation, Data analysis, Writing – review & editing. Tran Trung Dung: Writing – review & editing, Project administration. Tran Thi Minh Ha: Validation, Writing – review & editing. Phan Thi Thuc Quyen: Writing – review & editing, Funding acquisition. Bui Thi Khuyen: Investigation, Writing – review & editing. Nguyen Duc Huy: Conceptualization, Methodology, Writing – review & editing, Supervision.
![]()
References
- Radiah A, Misni S, Nazaruddin R, Norain Y, Adibi Rahiman M, et al. A preliminary agar study on the agar content and agar gel strength of Gracilaria manilaensis using different extraction methods. World Applied Sciences Journal, (2011); 15: 184-188.
- Temuujin U, Chi WJ, Chang YK, Hong SK. Identification and biochemical characterization of Sco3487 from Streptomyces coelicolor A3(2), an exo- and endo-type beta-agarase-producing neoagarobiose. Journal of Bacteriology, (2012); 194(1): 142-149.
- Veerakumar S, Manian R. Recombinant β-agarases: insights into molecular, biochemical, and physiochemical characteristics. 3 Biotech, (2018); 8(10): 445.
- Zhu Y, Zhao R, Xiao A, Li L, Jiang Z, et al. Characterization of an alkaline β-agarase from Stenotrophomonas sp. NTa and the enzymatic hydrolysates. International Journal of Biological Macromolecules, (2016); 86: 525-534.
- Cheong KL, Qiu HM, Du H, Liu Y, Khan BM. Oligosaccharides derived from red seaweed: production, properties, and potential health and cosmetic applications. Molecules, (2018); 23(10): 2451.
- Su Q, Jin T, Yu Y, Yang M, Mou H, et al. Extracellular expression of a novel beta-agarase from Microbulbifer sp. Q7, isolated from the gut of sea cucumber. AMB Express, (2017); 7(1): 220.
- Fu XT, Kim SM. Agarase: review of major sources, categories, purification method, enzyme characteristics and applications. Marine Drugs, (2010); 8(1): 200-218.
- Ramos KRM, Valdehuesa KNG, Banares AB, Nisola GM, Lee WK, et al. Overexpression and characterization of a novel GH16 beta-agarase (Aga1) from Cellulophaga omnivescoria W5C. Biotechnology Letters, (2020); 42(11): 2231-2238.
- Hu Z, Lin BK, Xu Y, Zhong MQ, Liu GM. Production and purification of agarase from a marine agarolytic bacterium Agarivorans sp. HZ105. Journal of Applied Microbiology, (2009); 106(1): 181-190.
- Lu X, Chu Y, Wu Q, Gu Y, Han F, et al. Cloning, expression and characterization of a new agarase-encoding gene from marine Pseudoalteromonas sp. Biotechnology Letters, (2009); 31(10): 1565-1570.
- Yun E, Yu S, Kim K. Current knowledge on agarolytic enzymes and the industrial potential of agar-derived sugars. Applied Microbiology and Biotechnology, (2017); 101(14): 5581-5589.
- Jang M, Lee D, Kim N, Yu K, Jang H, et al. Purification and characterization of neoagarotetraose from hydrolyzed agar. Journal of Microbiology and Biotechnology, (2009); 19(10): 1197-1200.
- Lakshmikanth M, Manohar S, Souche Y, Lalitha J. Extracellular β-agarase LSL-1 producing neoagarobiose from a newly isolated agar-liquefying soil bacterium, Acinetobacter sp., AG LSL-1. World Journal of Microbiology and Biotechnology (2006); 22: 1087-1094.
- Hong S, Lee J, Kim E, Yang H, Park J, et al. Anti-obesity and anti-diabetic effect of neoagarooligosaccharides on high-fat diet-induced obesity in mice. Marine Drugs, (2017); 15(4): 90.
- Aoki T, Araki T, Kitamikado M. Purification and characterization of a novel beta-agarase from Vibrio sp. AP-2. European Journal of Biochemistry, (1990); 187(2): 461-465.
- Sakai M, Deguchi D, Hosoda A, Kawauchi T, Ikenaga M. Ammoniibacillus agariperforans gen. nov., sp. nov., a thermophilic, agar-degrading bacterium isolated from compost. International Journal of Systematic and Evolutionary Microbiology, (2015); 65(Pt 2): 570-577.
- Roseline TL, Sachindra NM. Characterization of extracellular agarase production by Acinetobacter junii PS12B, isolated from marine sediments. Biocatalysis and Agricultural Biotechnology, (2016); 6219-226.
- Liu Y, Tian X, Peng C, Du Z. Isolation and characterization of an Eosinophilic GH 16 beta-agarase (AgaDL6) from an agar-degrading marine bacterium Flammeovirga sp. HQM9. Journal of Microbiology and Biotechnology, (2019); 29(2): 235-243.
- Parashar S, Kumar N. Studies on agarolytic bacterial isolates from agricultural and industrial soil. Iranian Journal of Microbiology, (2018); 10(5): 324-333.
- Faturrahman, Meryandini A, Junior M, Rusmana I. Isolation and identification of an agar-liquefying marine bacterium and some properties of its extracellular agarases. Biodiversitas Journal of Biological Diversity, (2011); 12(4): 192-197.
- George MG. Bergey’s manual of systematic bacteriology. 2005; Springer New York, NY.
- Buchanon RE, Gibbons NE. Bergey’s manual of determinative bacteriology. 1989; The Williams and Wilkins company, Baltimore.
- Sambrook J, Maccallum P, Russell D. Molecular cloning: a laboratory manual. 2001; Cold Spring Harbor Press: NY.
- Tamura K, Stecher G, Kumar S. MEGA11: Molecular evolutionary genetics analysis version 11. Molecular Biology and Evolution, (2021); 38: 3022-3027.
- Miller GL. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Analytical Chemistry, (1959); 31(3): 426–428.
- Lin B, Liu Y, Lu G, Zhao M, Hu Z. An agarase of glycoside hydrolase family 16 from marine bacterium Aquimarina agarilytica ZC1. FEMS Microbiology Letters, (2017); 364(4): fnx012.
- Dong Q, Ruan L, Shi H. A β-agarase with high pH stability from Flammeovirga sp. SJP92. Carbohydrate Research, (2016); 432: 1-8.
- Fu XT, Lin H, Kim SM. Purification and characterization of a novel beta-agarase, AgaA34, from Agarivorans albus YKW-34. Applied Microbiology and Biotechnology (2008); 78(2): 265-273.
- Hosoda A, Sakai M, Kanazawa S. Isolation and characterization of agar-degrading Paenibacillus spp. associated with the rhizosphere of spinach. Bioscience, Biotechnology, and Biochemistry, (2003); 67(5): 1048-1055.
- Suzuki H, Sawai Y, Suzuki T, Kawai K. Purification and characterization of an extracellular beta-agarase from Bacillus sp. MK03. Journal of Bioscience and Bioengineering, (2003); 95(4): 328-334.
- Duckworth M, Turvey JR. The action of a bacterial agarase on agarose, porphyran and alkali-treated porphyran. Biochemical Journal, (1969); 113(4): 687-692.
- Sakai M, Hosoda A, Ogura K, Ikenaga M. The growth of Steroidobacter agariperforans sp. nov., a novel agar-degrading bacterium isolated from soil, is enhanced by the diffusible metabolites produced by bacteria belonging to Rhizobiales. Microbes and Environments, (2014); 29(1): 89-95.
- Ikenaga M, Kataoka M, Yin X, Murouchi A, Sakai M. Characterization and distribution of agar-degrading Steroidobacter agaridevorans sp. nov., isolated from rhizosphere soils. Microbes and Environments, (2021); 36(1): ME20136.
- Zafar A, Aftab M, Iqbal I, Dind Z, Saleema M. Pilot-scale production of a highly thermostable α-amylase enzyme from Thermotoga petrophila cloned into E. coli and its application as a desizer in textile industry. RSC Advances, (2019); 9984-992.
- Potin P, Richard C, Rochas C, Kloareg B. Purification and characterization of the alpha-agarase from Alteromonas agarlyticus (Cataldi) comb. nov., strain GJ1B. European Journal of Biochemistry, (1993); 214(2): 599-607.
- Ahn BK, Min KC, Lee DG, Kim A, Lee SH. Characterization of agarase from a marine bacterium Agarivorans sp. BK-1. Journal of Life Science, (2019); 29(11): 1173-1178.
- Lakshmikanth M, Manohar S, Lalitha J. Purification and characterization of β-agarase from agar-liquefying soi bacterium Acinetobacter sp., AG LSL-1. Process Biochemistry, (2009); 44: 999–1003.
- Kirimura K, Masuda N, Iwasaki Y, Nakagawa H, Kobayashi R, et al. Purification and characterization of a novel β-agarase from an alkalophilic bacterium, Alteromonas sp. E-1. Journal of Bioscience and Bioengineering, (1999); 87: 436–441.
- Li J, Hu Q, Li Y, Xu Y. Purification and characterization of cold-adapted beta-agarase from an Antarctic psychrophilic strain. Brazilian Journal of Microbiology, (2015); 46(3): 683-690.
- Ziayoddin M, Lalitha, Junna, Shinde, Manohar. Optimization of agrase production by alkaline Pseudomonas aeruginosa ZSL-2 using Taguchi experimental design. International Letters of Natural Sciences, (2014); 17: 180-193.
- Zeng C, Zhang L, Miao S, Zhang Y, Zeng S, et al. Preliminary characterization of a novel beta-agarase from Thalassospira profundimonas. Springerplus, (2016); 5(1): 1086.
- Leon O, Quintana L, Peruzzo G, Slebe JC. Purification and properties of an extracellular agarase from Alteromonas sp. strain C-1. Applied and Environmental Microbiology, (1998); 58(12): 4060-4063.
- Shieh WY, Jean WD. Alterococcus agarolyticus, gen.nov., sp.nov., a halophilic thermophilic bacterium capable of agar degradation. Canadian Journal of Microbiology, (1998); 44(7): 637-645.
- Minegishi H, Shimane Y, Echigo A, Ohta Y, Hatada Y, et al. Thermophilic and halophilic β-agarase from a halophilic archaeon Halococcus sp. 197A. Extremophiles, (2013); 17(6): 931-939.
- Long J, Ye Z, Li X, Tian Y, Bai Y, et al. Enzymatic preparation and potential applications of agar oligosaccharides: a review. Critical Reviews in Food Science and Nutrition, (2022); 1-17.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0