

Full Length Research Article
Biolistic Inoculation of Selected Host Plants with different Begomoviruses and their associated Betasatellites: using partially digested rolling circle amplified Viral/Satellite DNA
Mehwish Rauf1́*, Muhammad Zia-Ur-Rehman2, Muhammad Saleem Haider1
Adv. life sci., vol. 10, no. 3, pp. 472-478, September 2023
*– Corresponding Author: Mehwish Rauf (mehwishrauf@gmail.com )
Authors' Affiliations
2. Govt. Graduate College of Science, Wahdat Road Lahore – Pakistan
[Date Received: 24/04/2023; Date Revised: 30/05/2023; Date Published: 30/09/2023]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: The foundation of modern diagnostics of plant pathogens are based on the confirmation of Koch's postulates. However, following the rules of Koch's postulates is not always straightforward for the viruses because of their non-culturability and difficulties associated with the inoculation approaches.
Methods: Biolistic inoculation of plants with partially digested rolling circle amplified (RCA) viral components was demonstrated in this study. The verified DNA bands were excised from the gel and self-circularized using the rapid DNA ligation kit. The self-ligated viral DNA components were enriched by RCA and partially digested with the respective fast digest restriction enzymes. The RCA prep (partially digested 3 µg of virus and 3 µg of beta-satellite) was biolistically inoculated into healthy plants of Solanum lycopersicum, Nicotiana tabacum and Capsicum annum at the two-true leaf stage.
Results: By using this method, extracted begomoviruses from cotton [ (Cotton leaf curl Kokhran virus-Burewala strain (CLCuKοV-Bu), Tomato leaf curl New Delhi virus (ToLCNDV) and Chili leaf curl virus (ChiLCV) and their associated Cotton leaf curl Multan betasatellite (CLCuMB)] were successfully inoculated and propagated from the inoculation site. Furthermore, it has been noted that these viruses exhibit diverse behaviors in various host plants. The capacity of these viruses to infect systemically in C. annum without causing the typical disease symptoms is of interest.
Conclusion: Biolistic inoculation using partially digested RCA of viral or satellite DNA enables viruses to infect plants with either monopartite or bipartite genomes. This method is easy and quick, and allows begomovirus cultures to be created without the use of Agrobacterium tumefaciens or whiteflies.
Keywords: Begomovirus; Rolling circle amplification; Koch's postulate; Inoculation
Introduction![]()
Geminiviruses are composed of 14 genera named Becurtovirus, Begomovirus, Curtovirus, Capulavirus, Citlodavirus, Eragrovirus, Grablovirus, Maldovirus, Mastrevirus, Mulcrilevirus, Opunvirus, Topcuvirus, Topilevirus, and Turncurtovirus. These genera differ in their host range, insect vectors, and genomic organization (ictv.global/report/geminiviridae). The genus Begomovirus is the largest and most economically important group within the family Geminiviridae [1,2] with at least 409 virus species (ICTV, 2020). The genomes of begomoviruses are either monopartite or bipartite. Bipartite begomoviruses comprise two genomic components known as DNA-A and DNA-B, while monopartite begomoviruses consist of just a single component, analogous to the DNA-A component of bipartite begomoviruses. Both DNA-A and DNA-B are about 2.6 and 2.8 kb in size and are transcribed bi-directionally [3-5]. The DNA-A component is in charge of replication and transcription, whereas DNA-B is necessary for inter- and intracellular virus movement. For a systemic infection, both components must be present [6]. The majority of the monopartite begomoviruses are frequently coupled with one or two satellite DNAs. Satellite DNAs are classified as alpha-satellites or beta-satellites based on how their DNA is organized and how they affect the symptoms caused by the helper begomovirus. They rely on their helper viruses for their replication and also help mitigate the symptoms they cause [7].
Experimental transmission of begomoviruses is difficult and requires extensive research efforts. Mechanical inoculation from plant to plant is possible for some begomoviruses, but only at low rates, and for others, it is not possible at all. Graft transmission necessitates the presence of a live, diseased plant, and not all plants are easily grafted. The whitefly vector can easily transmit most begomoviruses from plant to plant, but this requires a live, diseased plant and considerable facilities for its maintenance. Furthermore, none of these methods allow for long-term virus culture storage or the manipulation of viral DNA prior to inoculations. Inoculation techniques such as agro-inoculation and particle bombardment of DNA as an inoculum were developed for these reasons [8].
In the agro-inoculation method, A. tumefaciens is used to deliver binary vectors harboring a tandem repeat (dimer) of a cloned viral DNA into host cells [9]. As a result, genome-sized viral DNA is produced, which spreads systemically throughout the plant and causes disease symptoms. Agro inoculation, on the other hand, requires time-consuming subcloning processes to insert the viral DNA into the binary vector. This method also introduces A. tumefaciens, another pathogen, into the host plant. Agro-inoculation of cloned begomovirus DNA has been demonstrated to not always mimic whitefly transmission, which is likely due to the challenges encountered with A. tumefaciens in infecting particular hosts [10].
Particle bombardment for gene transfer gained significant attention from genetic engineers for a particular reason. The particle gun works where other procedures fail on occasion. In the 1960s, plant virologists developed the first generation of micro-projectile techniques by using high-velocity micro-projectiles (DNA-coated tungsten or gold particles) to injure plant cells and facilitate the entry of virus particles or DNA. This micro-projectile approach had the advantage of being technically simple and was applicable to all plant species [11]. By particle bombardment, plants have also been inoculated with cloned begomovirus DNA (using either monomer or dimer), which resulted in a high inoculation efficiency, thus limiting the need for complex DNA modifications and facilitating genetic analysis of begomoviruses [12,13].
This manuscript describes the development of a novel approach to inoculate test plants with begomoviruses. The partially digested rolling circular amplified viral DNA was used to develop a rapid and simple method. The versatility of this approach with regard to different begomoviruses and inoculated plants is demonstrated.
Methods![]()
Test Plants
The following plants were selected as host plants in this study: S. lycopersicum L. (cultivar Nagina), C. annuum L. (cultivar F1-Henna), and N. tabacum L. (cultivar Samsun). Certified seeds of S. lycopersicum, N. tabacum, and C. annuum were kindly provided by Dr. Idrees Ahmad Nasir (Centre of Excellence in Molecular Biology, University of the Punjab, Lahore, Pakistan). The seeds were sown in Pro-Mix and transferred to small, ideally 4-inch diameter plastic pots containing sterilized sandy loam soil when the seedlings were 3 weeks old. Plants were grown in an insect-free growth room under controlled conditions at 25-28 °C with a photoperiod of 16 h light/8 h dark, respectively.
Gene Gun Method
The RCA product was prepared and micro-projectiled by a hand-held gene gun (CEMB Laboratory).
Preparation of RCA Product for Bombardment
Procurement of Viral Clone
The different monomers of begomoviruses, CLCuKοV-Bu (Accession no. HF567942), ToLCNDV (Accession no. HM134234), and ChiLCV, and their associated betasatellite (CLCuMB) (Accession no. HF567946), were obtained from the Virology lab of the Faculty of Agricultural Sciences, PU, Lahore, Pakistan. These clones have been preserved and archived.
Restriction Digestion of Recombinant Plasmid and Self-circulization
Digestion of recombinant plasmids carrying the full-length genome of a virus/satellite were performed with the respective fast digest restriction enzymes (Pst I, Hind III and Sal I) (Thermo Scientific). Following restriction, the digested DNA was run on a 1% agarose gel stained with ethidium bromide at 80 V for 2 h in order to separate the 2.8 kb virus, 1.4 kb betasatellite, and 3 kb vector bands. The confirmed DNA fragments were gel purified by using the GeneJET gel extraction kit (Thermo Scientific) according to the protocol provided by the manufacturer. Each purified DNA was self-circularized using the rapid DNA ligation kit (Thermo Scientific) and used as a template for RCA.
Rolling Circle Amplification
Self-ligated viral DNA was enriched by RCA using the TempliPhi DNA amplification kit (GE Healthcare). For RCA, 3 µL of the template was added to 15 µL of sample buffer in a sterilized autoclaved tube. The sample was heated to 95 °C for 3 min, to convert double-stranded DNA into single-stranded DNA. After cooling, the sample was mixed with 15 µL of reaction buffer and 0.6 µL of enzyme mix and incubated at 30 °C for 18 h. At the end of this incubation, the ɸ 29 DNA polymerase was deactivated by heating at 65 °C for 10 min. The resultant RCA products were confirmed by restriction digestion with the respective restriction enzymes. After confirmation, the RCA product was partially digested and used for the coating of tungsten particles.
Optimization of Biolistic Inoculation of Plants
Preparation and Coating of Tungsten Particles
Tungsten particles were used as microprojectiles for the delivery of DNA into plant cells. 5.0 mg of tungsten particles (1.1 microns [um] =M 17A) were taken in an eppendorf tube. Particles were washed with 100% ethanol, allowed to dry in laminar air flow, and re-suspended in 100 µL of sterile distilled water (SDW). Tungsten particles (micro-projectiles) were coated with partially digested RCA. For this, 100% ethanol, 2.5 M calcium chloride, and 100 mM spermidine were filter sterilized using a 0.45 µm filter assembly. Tungsten particles in SDW were homogenized by vortexing at high speed, and 15 µL of RCA prep (3 µg of virus and 3 µg of betasatellite) was added. A total of 110 µL of calcium chloride and spermidine were added to each tube while vortexing at low speed. After 10 min of incubation on ice, tubes were spun down for 5 s at 10,000 rpm. The supernatant was discarded, and the pellet was washed with 250 µL of absolute ethanol. The pellet was re-suspended in 120 µL of absolute ethanol and placed on ice until use.
Host Plant Preparation for Inoculation
Plants at the two-true leaf stage were used. Each true leaf was inoculated twice, preferably along the main or side vein. Prior to inoculation, plants were covered with 16 oz. plastic cup cages with ventilated tops. Two plants per pot were ideal because inoculation stress causes the plants to wilt.
Shooting of Projectiles
For the bombardment in each experiment, 20 µL aliquots per plant were employed. 20 µL of suspension (coated tungsten particles) was spread on each filter assembly and allowed to dry in laminar airflow. The filter assembly was fixed in the leur-lock and the plant in the pot to be bombarded was placed in the middle of the chamber. Once the needle indicated 28 inches of Hg negative pressure, the gun's chamber was carefully shut and evacuated. As soon as the pressure gauge read 60 Kg/cm2, the helium gas vent was opened. The pre-chamber was closed, and the fire switch was gently pressed three times. Turning the tap until the needle reaches zero marks allows the inside air to be released. The door was unlocked, and the plant was removed from the chamber. The inoculated plants were placed in growth chamber and covered with cups for 1-2 days so that high humidity was maintained. After 2 days, cups were removed, and plants were monitored on a regular basis for the appearance of symptoms.
Confirmation of Viral DNA by Polymerase Chain Reaction
The appearance of typical disease symptoms was evaluated up to 35 days post-inoculation (dpi). Total genomic DNA was isolated from all the biolistically inoculated plants using cetyltrimethylammonium bromide (CTAB) method [14] and served as a template for PCR amplification. In each PCR reaction, 1 µg of DNA was used. PCR primers (Core (AV)/Core (AC)) were used to amplify 650 bp of the CP gene of DNA-A of the ToLCNDV, ChiLCV, and CLCuKοV-Bu. Similarly, primers (Beta-F/Beta-R) were used to amplify 1400 bp of the CLCuMB. For CP, the thermal cycling was performed as initial denaturation at 95 ºC for 1 min, and thereafter, at 95 ºC for 1 min, 58 ºC for 1 min, 72 ºC for 1 min for 35 cycles, and at 72 ºC for 10 min. The cycling parameters for beta-satellite were: initial denaturation at 94 ºC for 2 min, 35 cycles of denaturation at 94 ºC for 45 s, annealing at 54.5 ºC for 45 s, and extension at 72 ºC for 1 min, with a final extension at 72 ºC for 10 min. After confirmation, positive plants were maintained under insect-proof conditions for transmission studies.
Results![]()
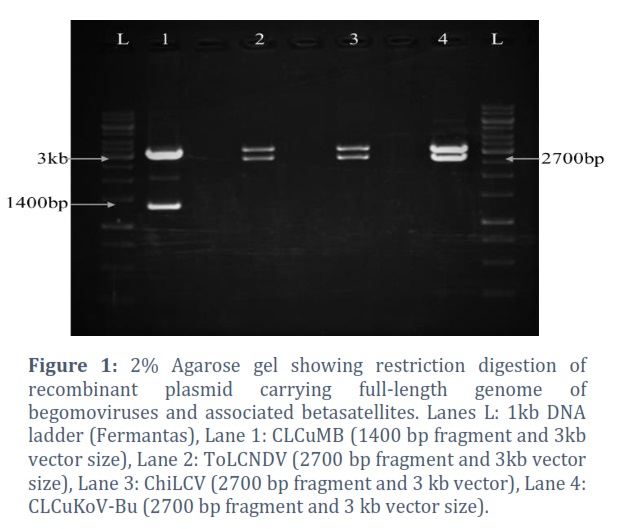
Restriction Digestion of Recombinant Plasmid
Cloned viral components of DNA-A (ToLCNDV, ChiLCV, and CLCuKοV-Bu) and CLCuMB were transformed into competent cells of E. coli, and plasmids were isolated. With the appropriate restriction enzymes, the isolated recombinant plasmids were digested and resolved on a 2% agarose gel (Figure 1).
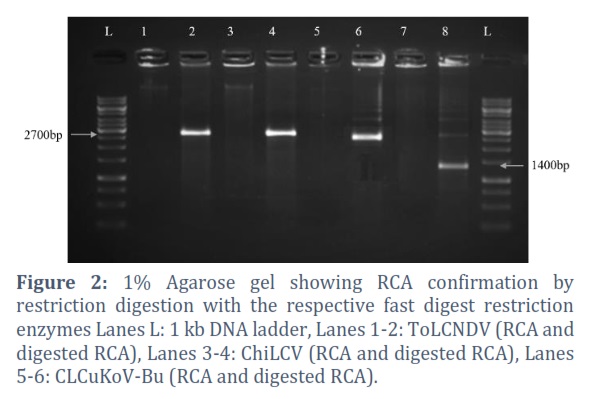
RCA of Self-Ligated Viral DNA and Confirmation by Restriction Enzymes
The confirmed DNA bands were purified from the gel and self-circularized using the Rapid DNA ligation kit. The self-ligated viral DNA components were enriched by RCA. For confirmation, RCA products were digested with the respective fast digest restriction enzymes (Figure 2).
Biolistic Inoculation of Plants and Symptoms Development
The confirmed RCA products were then partially digested with the respective fast digest restriction enzymes. The RCA prep (partially digested 3 µg of virus and 3 µg of beta-satellite) was biolistically inoculated into healthy plants of S. lycopersicum cv Nagina, N. tabacum cv Samsun, and C. annum cv F1-Henna at two-true leaf stage, to evaluate the viral component’s ability to infect systemically. Biolistic transformed plants were monitored periodically for the appearance of symptoms. At 35 dpi, all plants displayed mild to severe characteristic symptoms (Figure 3 and Table 1). Depending on the virus, infection rates ranged from very low to as high as 100%, similar to those obtained using the agro-inoculation technique, but with much less effort.
Due to inoculation stress (which causes the plants to wilt), 50% of the plant died, the number of inoculated plants was 13.
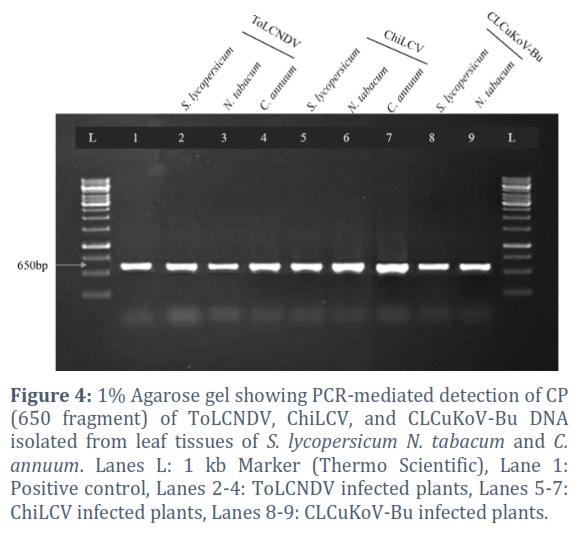
Confirmation of Viral DNA by PCR
The virus infection of biolistically inoculated plants was also confirmed by PCR (Figure 4). Total genomic DNA was isolated from the leaves of inoculated plants and subjected to PCR. A degenerate primer pair, Begomo Core (AV) and Core (AC), was used to amplify a 650 bp region of the begomovirus CP gene. Similarly, Beta-F and Beta-R primers were used to amplify 1400 bp of CLCuMB. The predicted size of the CP gene and beta-satellite could be amplified from symptomatic plants (mild to severe), whereas DNA isolated from mock-inoculated plants yielded no bands (Figure 3, panel 4). Overall, the results presented here demonstrated that ToLCNDV (DNA-A), ChiLCV (DNA-A), and CLCuKοV-Bu (DNA-A), with their associated CLCuMB can manage to replicate and spread from the site of inoculation.
It is also observed that these viruses behave distinctly in different host plants. Without causing characteristic disease symptoms in C. annum, the ability of these viruses to infect systemically is of interest. This finding further suggests that in C. annum, these viruses are unable to interact with factors involved in symptoms induction.
Figures & Tables

The present study reports a novel approach to inoculate test plants with selected begomoviruses and associated betasatellite. The partially digested rolling circular amplification of self-ligated viral DNA and betasatellite was used to develop a rapid and simple method for infectivity. The versatility of this approach with regard to different begomoviruses and inoculated plants is demonstrated. Results of the study presented here demonstrated that all the N. tabacum, and S. lycopersicum host plants were susceptible and displayed severe to mild symptoms when inoculated with ToLCNDV, ChiLCV, and CLCuKοV-Bu along with their cognate CLCuMB. Shahid et al. [15] reported that
the betasatellite, commonly associated with monopartite begomoviruses, can also enhance bipartite begomovirus virulence. Another study also revealed that coinfection of ToLCND with CLCuMB in S. lycopersicum and Nicotiana benthamiana enhanced viral DNA accumulation by suppressing the host defense and resulted in systemic infection. However, the movement functions encoded by the DNA-B component of ToLCNDV may be responsible for the rise in betasatellite and perhaps viral levels in co-infected cotton, allowing the infection to propagate to tissues that it would not typically reach [16].
The findings of our study are also in line with these studies and highlight the potential of betasatellite in symptom determination. The complementary sense strand of betasatellite has βC1 ORF, which is conserved among different betasatellites in terms of size and position. Constitutive expression and mutational analyses have shown that the βC1 protein is a strong symptom or pathogenicity determinant and plays a vital role in begomovirus infection [17]. This suggests that even relatively benign viruses or virus isolates can become aggressive when a beta-satellite is present. But these findings are not consistent with our observations in the case of C. annuum plants. Capsicum annuum host plants, when inoculated with ToLCNDV (DNA-A) and CLCuMB, were shown to be susceptible to the virus and the systemic spread of virus was confirmed by PCR. However, none of the plants displayed any symptoms. Numerous factors can contribute to the development of symptoms, but they all depend on the accumulation of viral proteins or nucleic acids that alter the plant's normal function and/or trigger a symptomatic defence response [18,19]. Previous studies have reported that the Rep of begomoviruses and curtoviruses interacts specifically with RBR, a key regulator of the plant cell cycle, via a unique motif. Any mutation of these motifs in Rep A and Rep leads to fewer symptoms and less virus accumulation in plants [20, 21].
This asymptomatic infectivity of ToLCNDV in C. annuum suggests that the virus might be unable to interact with factors causing symptoms. These factors are sometimes thought to be a part of the miRNA pathway, which is influenced by variables that determine the pathogenicity of viruses [22]. The C4 protein, which binds to siRNAs, suppresses PTGS and determines the pathogenicity of particular geminiviruses [23,24]. It has been shown that many host proteins, including SK4-1/SKK, and shaggy-like protein kinases, interact with the C4 protein in geminiviruses. This connection is crucial for C4 protein function to inhibit gene silencing and induce disease symptoms [25]. These findings imply that virus and host factors are important in the development of symptoms, which are probably connected to viral sequences and their matches to certain plant cellular mRNAs.
Biolistic inoculation with partially digested RCA of viral components proved successful for selected begomoviruses. This method allows viruses to infect plants regardless of whether they have monopartite or bipartite genomes. While we used this method with begomoviruses, this same method will most likely work with other viruses in the Geminiviridae family and their associated satellites.
Acknowledgment
The first author acknowledges Prof. Dr. Ahmad Ali Shahid, and Prof. Dr. Bushra Rashid (CEMB, PU, Lahore, Pakistan) for providing access to their laboratory facilities and for their all-time availability and sincere cooperation.
Conflict of Interest
The authors declare no conflict of interest.
M Rauf: Conceptualization, Methodology, Writing – Review & Editing English
M Zia-Ur-Rehman: Conceptualization, Investigation,
Muhammad Saleem Haider: Supervision, Validation
All authors have read and agreed to the published version of the manuscript.
![]()
References
- Zerbini FM, Briddon RW, Idris A, Martin DP, Moriones E, Navas-Castillo J, Rivera-Bustamante R, Roumagnac P, Varsani A, Consortium IR. ICTV virus taxonomy profile: Geminiviridae. The Journal of general virology, (2017);98(2):131.
- Walker PJ, Siddell SG, Lefkowitz EJ, Mushegian AR, Dempsey DM, Dutilh BE, Harrach B, Harrison RL, Hendrickson RC, Junglen S, Knowles NJ. Changes to virus taxonomy and the International Code of Virus Classification and Nomenclature ratified by the International Committee on Taxonomy of Viruses (2019). Archives of virology, (2019);164(9):2417-29.
- Varsani A, Navas-Castillo J, Moriones E, Hernández-Zepeda C, Idris A, Brown JK, Murilo Zerbini F, Martin DP. Establishment of three new genera in the family Geminiviridae: Becurtovirus, Eragrovirus and Turncurtovirus. Archives of virology, (2014);159:2193-2203.
- Anwar I, Bukhari HA, Nahid N, Rashid K, Amin I, Shaheen S, Hussain K, Mansoor S. Association of cotton leaf curl Multan betasatellite and Ageratum conyzoides symptomless alphasatellite with tomato leaf curl New Delhi virus in Luffa cylindrica in Pakistan. Australasian Plant Pathology, (2020);49:25-29.
- Fiallo-Olivé E, Navas-Castillo J. Molecular and biological characterization of a New World mono-/bipartite begomovirus/deltasatellite complex infecting Corchorus siliquosus. Frontiers in microbiology, (2020);11:1755.
- Hanley-Bowdoin L, Bejarano ER, Robertson D, Mansoor S. Geminiviruses: masters at redirecting and reprogramming plant processes. Nature Reviews Microbiology, (2013);11(11):777-788.
- Idris AM, Shahid MS, Briddon RW, Khan AJ, Zhu JK, Brown JK. An unusual alphasatellite associated with monopartite begomoviruses attenuates symptoms and reduces betasatellite accumulation. Journal of General Virology, (2011);92(3):706-717.
- Guenoune-Gelbart D, Sufrin-Ringwald T, Capobianco H, Gaba V, Polston JE, Lapidot M. Inoculation of plants with begomoviruses by particle bombardment without cloning: Using rolling circle amplification of total DNA from infected plants and whiteflies. Journal of virological methods, (2010);168(1-2):87-93.
- Grimsley N, Hohn T, Davies JW, Hohn B. Agrobacterium-mediated delivery of infectious maize streak virus into maize plants. Nature, (1987);325(6100):177-179.
- Buragohain AK, Sung YK, Coffin RS, Coutts RH. The infectivity of dimeric potato yellow mosaic geminivirus clones in different hosts. Journal of general virology, (1994);75(10):2857-2861.
- MacKenzie DR, Anderson PM, Wernham CC. A mobile air blast inoculator for plot experiments with maize dwarf mosaic virus. Plant Dis. Rep, (1966);50:363-367.
- Garzón-Tiznado JA, Torres-Pacheco I, Ascencio-Iba ez JT, Herrera-Estrella L. Inoculation of peppers with infectious clones of a new geminivirus by a biolistic procedure. Phytopathology-New York And Baltimore Then St Paul, (1993);83:514-514.
- Bonilla-Ramírez GM, Guevara-González RG, Garzon-Tiznado JA, Ascencio-Ibanez JT, Torres-Pacheco I, Rivera-Bustamante RF. Analysis of the infectivity of monomeric clones of pepper huasteco virus. Journal of general virology, (1997);78(4):947-951
- Doyle JJ. Isolation of plant DNA from faesh tissue. Focus, (1990);12:13-15.
- Shahid MS, Mansoor S, Brown JK, Briddon RW. Interaction of a tomato leaf curl New Delhi virus with a betasatellite enhances symptom severity in field-infected tomato plants. Tropical Plant Pathology, (2021);46:169-174.
- Jyothsna P, Haq QM, Singh P, Sumiya KV, Praveen S, Rawat R, Briddon RW, Malathi VG. Infection of tomato leaf curl New Delhi virus (ToLCNDV), a bipartite begomovirus with betasatellites, results in enhanced level of helper virus components and antagonistic interaction between DNA B and betasatellites. Applied Microbiology and Biotechnology, (2013);97:5457-5471.
- Saeed M, Behjatnia SA, Mansoor S, Zafar Y, Hasnain S, Rezaian MA. A single complementary-sense transcript of a geminiviral DNA β satellite is determinant of pathogenicity. Molecular Plant-Microbe Interactions, (2005);18(1):7-14.
- Culver JN, Padmanabhan MS. Virus-induced disease: altering host physiology one interaction at a time. Annu. Rev. Phytopathol, (2007);45:221-243.
- Pallas V, García JA. How do plant viruses induce disease? Interactions and interference with host components. Journal of General Virology, (2011);92(12):2691-2705.
- Kong LJ, Orozco BM, Roe JL, Nagar S, Ou S, Feiler HS, Durfee T, Miller AB, Gruissem W, Robertson D, Hanley-Bowdoin L. A geminivirus replication protein interacts with the retinoblastoma protein through a novel domain to determine symptoms and tissue specificity of infection in plants. The EMBO journal, (2000);19(13):3485-3495.
- McGivern DR, Findlay KC, Montague NP, Boulton MI. An intact RBR-binding motif is not required for infectivity of Maize streak virus in cereals, but is required for invasion of mesophyll cells. Journal of general virology, (2005);86(3):797-801.
- Amin I, Patil BL, Briddon RW, Mansoor S, Fauquet CM. Comparison of phenotypes produced in response to transient expression of genes encoded by four distinct begomoviruses in Nicotiana benthamianaand their correlation with the levels of developmental miRNAs. Virology Journal, (2011);8(1):1-24.
- Vanitharani R, Chellappan P, Pita JS, Fauquet CM. Differential roles of AC2 and AC4 of cassava geminiviruses in mediating synergism and suppression of posttranscriptional gene silencing. Journal of virology, (2004);78(17):9487-9498.
- Pandey P, Choudhury NR, Mukherjee SK. A geminiviral amplicon (VA) derived from Tomato leaf curl virus (ToLCV) can replicate in a wide variety of plant species and also acts as a VIGS vector. Virology Journal, (2009);6(1):1-3.
- Dogra SC, Eini O, Rezaian MA, Randles JW. A novel shaggy-like kinase interacts with the Tomato leaf curl virus pathogenicity determinant C4 protein. Plant molecular biology, (2009);71:25-38.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0