

Review Article
Overdominance in livestock breeding: examples and current status
Anh Phu Nam Bui1*, Trinh Lam Hoang Tam1, Pham Thi Phuong1, Nguyen Thuy Linh2
Adv. life sci., vol. 10, no. 4, pp. 525-529, December 2023
*– Corresponding Author: Anh Phu Nam Bui (anh.bpn@ou.edu.vn)
Authors' Affiliations
2. School of Agriculture and Aquaculture, Tra Vinh University – Vietnam
[Date Received: 31/10/2021; Date Revised: 26/07/2023; Date Published Online: 31/12/2023]
Editorial Note on Version of Record
24 June 2025: This article has been corrected. See https://doi.org/10.62940/als.v13i0.4216 for more information.
Abstract![]()
Introduction
Methods
Discussion
Conclusion
References
Abstract
Recent data have revealed that genetic variation could be attributed to overdominance, or heterozygote advantage. However, the genomic survey showed that only a small number of genes with polymorphisms maintained by overdominance, which is consistent with many published papers. Google Web, Google Scholar, NCBI Databases, and OMIC Tools were used to obtain data for this review paper. Different keywords were used to retrieve the required research articles and bioinformatics-based information, such as “overdominance’’ and “overdominance in animals. Research papers used for this review were published over the last 10 to 15 years, and information regarding overdominance in livestock was considered for the current review. It is hoped that in the future, more loci with overdominance will be discovered. In this review, we will illustrate eight examples of overdominance in livestock. We also want to emphasize that given a low number of reported cases of overdominance, it does not reflect the unimportance of heterozygote advantage in adaptive functions.
Keywords: Genomic survey; Polymorphisms; Livestock breeding
Introduction![]()
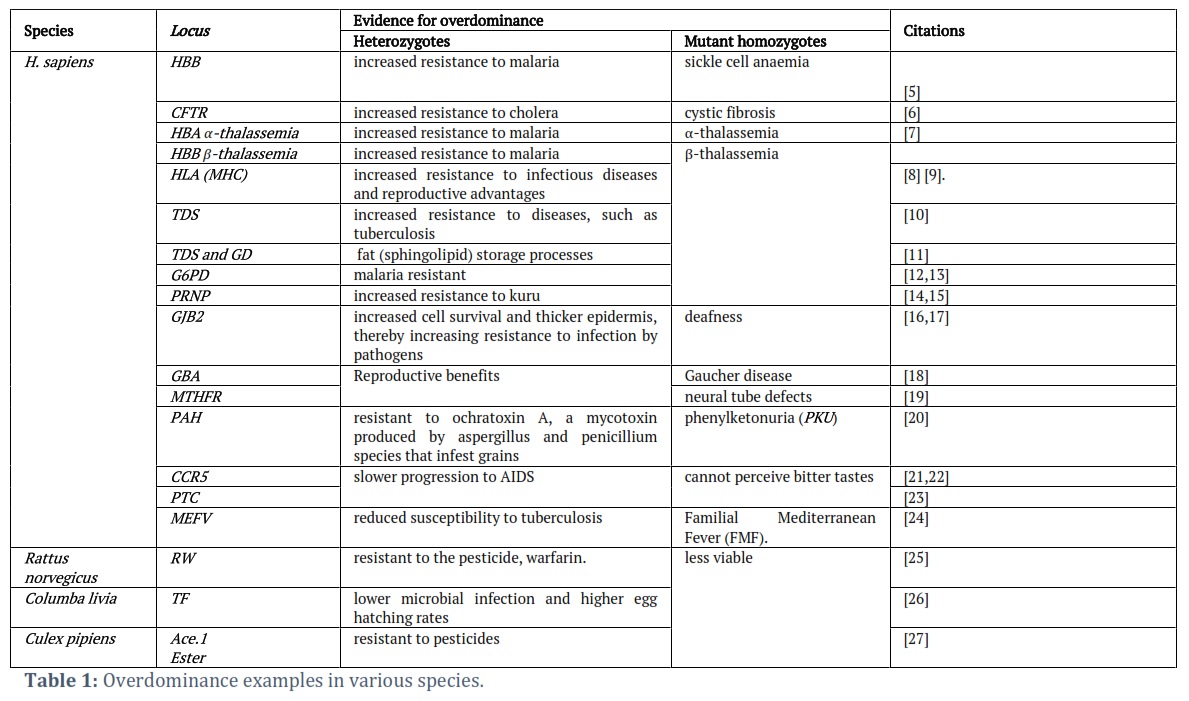
Overdominance is defined as heterozygous individuals having higher fitness over in both heterozygous and mutant homozygous forms. Overdominance, which is often used interchangeably with the term heterozygote advantage, has been used to provide a plausible explanation for genetic polymorphism in a particular population during natural or artificial selection [1]. Since its first documentation in 1922 [2], there have been only a small number of cases that can be classified as overdominance, which are often associated with disease resistance [3]. In Table 1, we presented a comprehensive, but not exhaustive, summary of overdominance examples collected from literature searches from human (Homo sapiens), brown rat (Rattus norvegicus), rock dove (Columba livia), and common house mosquito (Culex pipiens).
However, in these examples, the fitness of genotypes and their association with the infectious disease are not clearly elucidated. Moreover, some of the examples illustrated two directions of selection pressure. For instance, a classical textbook for overdominance is sickle-cell variations, including malaria, which favors heterozygotes and sickle-cell anaemia, which favors normal homozygotes. This polymorphism is not considered as overdominance since (i) heterozygotes should have the superior fitness over homozygotes, (ii) both malaria and sickle-cell anaemia affect survival, and (iii) the fitness of heterozygotes is environment-dependent and present only when malaria is present [3].
As a result, to fully portray overdominance in the current generation, the following indicators must be known (i) the DNA sequence of the gene and its mutant alleles under selection must be pre-determined; (ii) the relative fitness of each genotype must be measured (with heterozygotes exhibiting the highest relative fitness); (iii) the mechanism of selection must be understood i.e., the reason why heterozygotes are fitter than homozygotes [4].
When applying these criteria, it appears that there are not as many examples of overdominance as expected. Nevertheless, some persuasive examples have been collected from many studies on livestock. Here, we will illustrate and discuss these examples to portray genetic variations in the population by the maintenance of heterozygous individuals despite the lethality of the mutant alleles in homozygotes.
Methods
Literature search and selection criteria
Google Web, Google Scholar, NCBI Databases, and OMIC Tools were used to obtain data for this review paper. Different keywords were used to retrieve the required research articles and bioinformatics-based information, such as “overdominance’’ and “overdominance in animals. Research papers used for this review were those published over the last 10 to 15 years, and information regarding overdominance in animals was considered for the current review.
Methods![]()
Literature search and selection criteria
Google Web, Google scholar, NCBI Databases and OMIC Tools were used to obtain data for this review paper. Different key words were used to retrieve the required research articles and bioinformatics-based information, such as “overdominance’’ and “overdominance in animals’’. Research papers used for this review were those published over last 10 to 15 years and information regarding overdominance in animal was considered for current review.
Discussion![]()
Examples in livestock selection
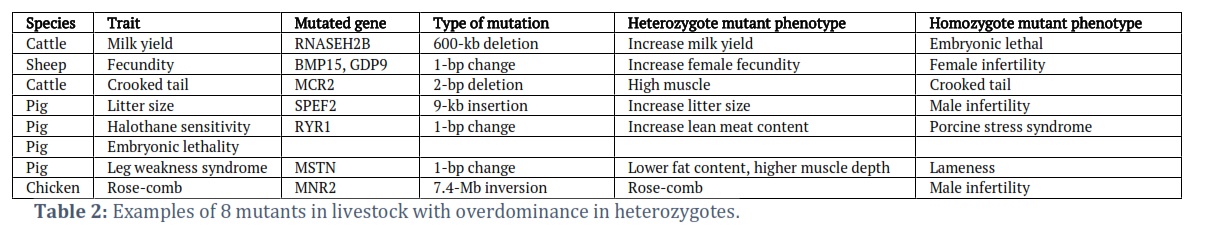
Numerous mutants in livestock recently identified by implementing molecular biology approaches appear to exhibit overdominance. Since the application of artificial insemination in breeding, the frequency of mutant alleles increases because of the superiority of the heterozygotes over mutant homozygotes and wild type homozygotes. Below, we will address eight examples of overdominance mutants in livestock, including 3 cases in cattle, 4 cases in pigs, and 1 case in poultry. The general information of these cases is presented in Table 2.
Milk yield
In cows, genetic correlation illustrates that milk yield and composition are negatively correlated with fertility. It is widely assumed that because of the negative energy balance during lactation of high-producing cows, their fertility is reduced [28]. In 2014, a fine mapping study conducted by Khadri et al. in Nordic Red cattle discovered a 660kb deletion that causes embryonically lethal mutation [29]. This deletion encompasses four genes, including RNASEH2B (ribonuclease H2, subunit B). The RNaseH2B gene encodes for the non-catalytic subunit of RNase H2, an endonuclease that specifically degrades the RNA of RNA: DNA hybrids and participates in DNA replication. RNASEH2B loss-of-function mutations are documented to cause embryonic death in mice [30,31] and Aicardi-Goutières syndrome type 2 in humans (AGS2, OMIM 610181). Nevertheless, it is also revealed that the deletion had positive effects on milk yield and composition in the heterozygous form. Upon the genetic survey, the presence of the heterozygote for this mutant in the Danish, Swedish, and Finnish Red Cattle was 13%, 23% and 32%, respectively. Due to the high prevalence of highly lethal alleles in the sampled population, it is hypothesized that conflicting characteristics of the mutant allele in fertility and milk yield and composition have led to the reduction of fertility in dairy cattle in recent years.
Fecundity
Mutations on the BMP15 or GDF9 genes have been revealed to affect female fecundity in domesticated sheep that display overdominance [32]. Both genes encode proteins that belong to the transforming growth factor ß superfamily. BMP15 and GDF9 proteins regulate ovarian function. Reduced expression of one of these genes may be associated with polycystic ovary syndrome. Heterozygous mutants on one of these genes increase ovulation rates and fecundity, while homozygotes reduce oocyte development and maturation. The observed frequencies of mutant alleles appear high in some breeds [33]. In 2014, a survey showed that there were more than 33% mutant alleles in the Belclare breed for the GDF9 gene.
Crooked tail
Loss-of-function mutation of the mannose receptor MCR2 has been reported to cause crooked tail phenotype in Belgian Blue beef cattle. MCR2 is a constitutively recycling endocytic receptor belonging to the mannose receptor family. It is found to bind and internalize both intact and degraded collagens and, in turn, take part in the turnover of collagens in both cytomembrane and extracellular matrix. In heterozygous individuals, this mutant phenotype is characterized by an increase in muscle mass, leading to skeletal and muscular malformations [34]. Although this phenotype is not lethal, some cases have been documented in which the heterozygote has retarded growth and poor meat quality. Thanks to the molecular biology approach, the mutant allele has been gradually eliminated from the population.
Embryonic lethality
The pig fetus is implanted by day 35 and born at day 114 of gestation. It may die at any time between those days for several reasons. Piglets that die and remain in the uterus undergo a series of standard changes. The skin loses its color, the eyes sink, and the placenta becomes darker. As water is removed, the fetus becomes drier and finally becomes dark brown, enveloped in its placenta. There have been several reasons attributed to mummified piglets, one of which was recently discovered in 2018 by Derk et al. BBS9 protein’s function has not been identified in pigs, although BBS9 mutants in mice showed embryonic lethality [35]. However, a 212-kb deletion has caused a truncated BBS9 protein, leading to a lower expression of the BMPER gene in swine. The homozygotic mutants cause fetal death, while heterozygotes increase growth rates [36].
Litter size
Immotile, short-tail sperm phenotype defect (ISTS) is a reproductive problem detected in Finnish Yorkshire population boars in the 1990s. This phenotype is characterized by lowered sperm counts, short sperm tails, and sperm structure abnormalities. In 2012, Sironen et al. (2012) successfully identified an insertion in an intron region of SPEF2 (Sperm flagellar protein 2) to be the causal mutation for ISTS [37]. SPEF2 is required for correct axoneme development in spermatozoa. Surprisingly, the frequency of SPEF2 mutant carriers in the pig population increased to 36% in 2001, which has led to the speculation of overdominance. Observation subsequently showed that SPEF2 mutant carriers have significantly higher litter size in first parity (0.51 piglets higher) than females not having the insertion.
Porcine stress syndrome
Pigs with porcine stress syndrome can be fatal without intervention. Other symptoms include behavioral disturbances, discoloration of the skin, and muscle rigidity. A homozygote for a single-nucleotide mutation in the RYR1 (ryanodine receptor 1) gene is responsible for this syndrome [38]. RYR1 protein is part of a group of related proteins called ryanodine receptors, which form channels that, when turned on, release positively charged calcium atoms from storage within cells. RYR1 channels play a critical role in muscles used for movement. Heterozygotes for this mutant exhibit higher lean meat content and larger musculature due to the increase in muscle contraction, resulting in more burning fat and muscle growth [39]. A survey conducted by O’Brien et al. (1993) depicted the presence of the mutant allele with the highest frequency in Pietrain (51.7% heterozygotes and 44.8% homozygotes) and in Landrace (33.2% heterozygotes, 2.1% homozygotes). After the introduction of DNA testing, the mutants can be molecularly identified, and the mutant allele can be eliminated from these breeds.
Leg weakness syndrome
Leg weakness, or lameness, is a serious problem in pig breeding due to its adverse impacts on animal welfare and productivity. Several studies conducted in various breeds reported there is a significant association between lameness and heritability, especially in Landrace breeds. By using whole-genome sequencing approaches, Matika et al. (2019) identified a mutation causing a premature stop codon in exon 3 of the MSTN gene on chromosome 15. MSTN is a member of the transforming growth factor beta (TGF-β) superfamily, which is highly conserved across species, and is typically expressed in developing and mature skeletal muscle as a key regulator of muscle growth [40]. In the homozygotic form, piglets suffer from lameness syndrome and do not survive post-40 kg live weight. However, heterozygotes have higher muscle depth and lower fat depth compared to wild type, suggesting that the deleterious allele was maintained at moderate frequency due to overdominance.
Rose comb
Rose-comb is a widely described monogenic trait that was first reported by William Bateson [41]. This trait has been documented in many breeds and is found in both heterozygotic and mutant homozygotic forms. The wild-type chicken exhibits a single comb phenotype, while the mutant chicken shows extensive phenotypic variability. As a result, the rose-comb phenotypes are influenced by several genes and represent an excellent model for gene interactions. The mutant phenotype is due to the inversion of 7.4Mb that relocates the gene homeodomain protein MNR2. Consequently, this leads to misexpression of MNR2 and disruption of the gene coiled-coil domain-containing protein 108 (CCDC108), causing poor sperm mobility in homozygotes and male fertility. However, the female chicken shows a normal phenotype in a mutant homozygote [42].
Figures & Tables
Overdominance remains a popular and persuasive explanation for genetic variation in a specific herd. Since its definition first proposed almost 100 years ago, the number of mutant alleles maintained by overdominance is low. To date, genomic data have only suggested a small number of genes that can be categorized as overdominant. However, it does not reflect that overdominance is not essential. In this review, we discuss eight examples that can be readily available for adaptations to new environmental challenges. Perhaps further analysis is needed to identify more genes regulated by overdominance.
Conflict of Interest
The authors declare that there is no conflict of interest.
ABPN and TVN conceived the project. TVN conducted literature searching. All the authors read and provided the edits. ABPN approved the manuscript.
![]() References
References
- Allison AC. Protection afforded by sickle-cell trait against subtertian malareal infection. British medical journal, (1954); 1(4857): 290-294.
- Fisher RA. XXI.—On the Dominance Ratio. Proceedings of the Royal Society of Edinburgh, (2014); 42321-341.
- Hedrick PW. What is the evidence for heterozygote advantage selection? Trends in Ecology & Evolution, (2012); 27(12): 698-704.
- Gemmell NJ, Slate J. Heterozygote Advantage for Fecundity. PLOS ONE, (2006); 1(1): e125.
- Allison AC. Protection afforded by sickle-cell trait against subtertian malarial infection. British Medical Association, (1954); 1(4857): 290-294.
- Gabriel SE, Brigman KN, Koller BH, Boucher RC, Stutts MJ. Cystic-fibrosis heterozygote resistance to cholera-toxin in the cystic-fibrosis mouse model. Science, (1994); 266(5182): 107-109.
- Flint J, Hill AVS, Bowden DK, Oppenheimer SJ, Sill PR, et al. High-frequencies of alpha-thalassemia are the result of natural-selection by malaria. Nature, (1986); 321(6072): 744-750.
- Penn DJ, Damjanovich K, Potts WK. MHC heterozygosity confers a selective advantage against multiple-strain infections. Proceedings of the National Academy of Sciences USA (2002); 99(17): 11260-11264.
- Sauermann U, Nurnberg P, Bercovitch FB, Berard JD, Trefilov A, et al. Increased reproductive success of MHC class II heterozygous males among free-ranging rhesus macaques. Hum Genet, (2001); 108(3): 249-254.
- Yokoyama S. Role of genetic drift in the high frequency of Tay-Sachs disease among Ashkenazic Jews. Annals of Human Genetics, (1979); 43(2): 133-136.
- Peleg L, Frisch A, Goldman B, Karpaty M, Narinsky R, et al. Lower frequency of Gaucher disease carriers among Tay-Sachs disease carriers. European Journal of Human Genetics, (1998); 6(2): 185-186.
- Luzatto L, FA U, Reddy S. Glucose-6-phosphate dehydorgenase deficient red cells: resistant to infection by malarial parasites. Science, (1969); 164839-842.
- Ruwende C, Khoo SC, Snow RW, Yates SNR, Kwiatkowski D, et al. Natural selection of hemi- and heterozygotes for G6PD deficiency in Africa by resistance to severe malaria. Nature, (1995); 376(6537): 246-249.
- Hedrick PW. A heterozygote advantage. Science, (2003); 302(5642): 57-57.
- Mead S, Stumpf MPH, Whitfield J, Beck JA, Poulter M, et al. Balancing selection at the prion protein gene consistent with prehistoric kuru like epidemics. Science, (2003); 300(5619): 640-643.
- Common JEA, Di WL, Davies D, Kelsell DP. Further evidence for heterozygote advantage of GJB2 deafness mutations: a link with cell survival. Journal of Medical Genetics, (2004); 41(7): 573-575.
- Meyer CG, Amedofu GK, Brandner JM, Pohland D, Timmann C, et al. Selection for deafness? Nature Medicine, (2002); 8(12): 1332-1333.
- Boas FE. Linkage to Gaucher mutations in the Ashkenazi population: Effect of drift on decay of linkage disequilibrium and evidence for heterozygote selection. Blood Cells, Molecules and Diseases, (2000); 26(4): 348-359.
- Schwartz CE, Hunter AGW, Holmes LB, Guttormsen SA, Tackels DC, et al. The methylenetetrahydrofolate reductase (MTHFR) C677T polymorphism: An example of heterozygote advantage by natural selection. American Journal of Human Genetics, (1997); 61(4): A211-A211.
- Woolf LI. The heterozygote advantage in phenylketonuria. Am J Hum Genet, (1986); 38(5): 773-775.
- Dean M, Carrington M, Winkler C, Huttley GA, Smith MW, et al. Genetic restriction of HIV-1 infection and progression to AIDS by a deletion allele of the CKR5 structural gene. Science, (1996); 273(5283): 1856-1862.
- Liu R, Paxton WA, Choe S, Ceradini D, Martin SR, et al. Homozygous defect in HIV-1 coreceptor accounts for resistance of some multiply-exposed individuals to HIV-1 infection. Cell, (1996); 86(3): 367-377.
- Wooding S, Kim UK, Bamshad MJ, Larsen J, Jorde LB, et al. Natural selection and molecular evolution in PTC, a bitter-taste receptor gene. American Journal of Human Genetics, (2004); 74(4): 637-646.
- Torosyan Y, Aksentijevich I, Sarkisian T, Astvatsatryan V, Ayvazyan A, et al. A population-based survey reveals an extremely high FMF carrier frequency in Armenia, suggesting heterozygote advantage. American Journal of Human Genetics, (1999); 65(4): A400-A400.
- Greaves JH, Redfern R, Ayres PB, Gill JE. Warfarin resistance – balanced polymorphism in Norway rat. Genetics Research, (1977); 30(3): 257-263.
- Frelinger JA. Maintenance of transferrin polymorphism in pigeons. Proceedings of the National Academy of Sciences U S A, (1972); 69(2): 326-329.
- Lenormand T, Bourguet D, Guillemaud T, Raymond M. Tracking the evolution of insecticide resistance in the mosquito Culex pipiens. Nature, (1999); 400(6747): 861.
- Lucy MC. Reproductive loss in high-producing dairy cattle: where will it end? Journal of Dairy Science, (2001); 84(6): 1277-1293.
- Kadri NK, Sahana G, Charlier C, Iso-Touru T, Guldbrandtsen B, et al. A 660-Kb Deletion with Antagonistic Effects on Fertility and Milk Production Segregates at High Frequency in Nordic Red Cattle: Additional Evidence for the Common Occurrence of Balancing Selection in Livestock. PLOS Genetics, (2014); 10(1): e1004049.
- Reijns MA, Rabe B, Rigby RE, Mill P, Astell KR, et al. Enzymatic removal of ribonucleotides from DNA is essential for mammalian genome integrity and development. Cell, (2012); 149(5): 1008-1022.
- White JK, Gerdin AK, Karp NA, Ryder E, Buljan M, et al. Genome-wide generation and systematic phenotyping of knockout mice reveals new roles for many genes. Cell, (2013); 154(2): 452-464.
- Davis GH. Major genes affecting ovulation rate in sheep. Genetics Selection Evolution, (2005); 37(1): S11.
- Hanrahan JP, Gregan SM, Mulsant P, Mullen M, Davis GH, et al. Mutations in the genes for oocyte-derived growth factors GDF9 and BMP15 are associated with both increased ovulation rate and sterility in Cambridge and Belclare sheep (Ovis aries). Biol Reprod, (2004); 70(4): 900-909.
- Fasquelle C, Sartelet A, Li W, Dive M, Tamma N, et al. Balancing Selection of a Frame-Shift Mutation in the MRC2 Gene Accounts for the Outbreak of the Crooked Tail Syndrome in Belgian Blue Cattle. PLOS Genetics, (2009); 5(9): e1000666.
- Veleri S, Bishop K, Dalle Nogare DE, English MA, Foskett TJ, et al. Knockdown of Bardet-Biedl syndrome gene BBS9/PTHB1 leads to cilia defects. PLoS One, (2012); 7(3): e34389.
- Derks MFL, Lopes MS, Bosse M, Madsen O, Dibbits B, et al. Balancing selection on a recessive lethal deletion with pleiotropic effects on two neighboring genes in the porcine genome. PLOS Genetics, (2018); 14(9): e1007661.
- Sironen A, Uimari P, Iso-Touru T, Vilkki J. L1 insertion within SPEF2 gene is associated with increased litter size in the Finnish Yorkshire population. Journal of Animal Breeding and Genetics, (2012); 129(2): 92-97.
- Fujii J, Otsu K, Zorzato F, de Leon S, Khanna VK, et al. Identification of a mutation in porcine ryanodine receptor associated with malignant hyperthermia. Science, (1991); 253(5018): 448-451.
- Salmi B, Trefan L, Bloom-Hansen J, Bidanel JP, Doeschl-Wilson AB, et al. Meta-analysis of the effect of the halothane gene on 6 variables of pig meat quality and on carcass leanness. Journal of Animal Science, (2010); 88(9): 2841-2855.
- McPherron AC, Lee S-J. Double muscling in cattle due to mutations in the myostatin gene. Proceedings of the National Academy of Sciences, (1997); 94(23): 12457-12461.
- Bateson W. Experiments with poultry. Reports to the Evolution Committee of the Royal Society, (1902); 1(87-124).
- Imsland F, Feng C, Boije H, Bed’hom B, Fillon V, et al. The Rose-comb mutation in chickens constitutes a structural rearrangement causing both altered comb morphology and defective sperm motility. PLoS Genetics, (2012); 8(6): e1002775.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0