

Full Length Research Article
Naringenin as a potent inhibitor molecule for targeting microtubule affinity-regulating kinase 4 (mark4): a molecular docking and in vitro study for therapeutics of Alzheimer’s disease
Mohammad Zubair Alam1,2*, Hala Abubaker Bagabir3, Mohammad Alameen Faisal Zaher4, Thamer M. A. Alqurashi5, Badrah S. Alghamdi1, 6, Ghulam Md Ashraf1, 2, Mohsin Kazi7
Adv. life sci., vol. 11, no. 1, pp. 136-143, February 2024
*– Corresponding Author: Mohammad Zubair Alam (mohdzubairalam@yahoo.com)
Authors' Affiliations
2. Department of Medical Laboratory Technology, Faculty of Applied Medical Sciences, King Abdulaziz University, Jeddah – Saudi Arabia
3. Department of Physiology, Faculty of Medicine, King Abdulaziz University, Rabigh Campus – Saudi Arabia
4. Department of Physiology, Faculty of Medicine, King Abdulaziz University, Jeddah – Saudi Arabia
5. Department of Pharmacology, Faculty of Medicine, Rabigh Campus, King Abdulaziz University – Saudi Arabia
6. Department of Physiology, Neuroscience Unit, Faculty of Medicine, King Abdulaziz University, Jeddah 22252 – Saudi Arabia
7. Department of Pharmaceutics, College of Pharmacy, P.O. BOX-2457, King Saud University, Riyadh 11451 – Saudi Arabia
[Date Received: 01/08/2023; Date Revised: 09/10/2023; Date Published: 25/02/2024]
Editorial Note:
28 May 2025: While reconciling the record of Turnitin originality analysis, 49% content of this article was found generated using AI tool/s without clear disclosure along with similarity on more than 4% with at least one source published prior to this article.
Editorial board of Advancements in Life Sciences has started the process of retracting this article due to the above post-publication findings. The process shall be concluded after registering responses from the authors. Meanwhile, full text of the article shall remain unavailable for citations (this notice has been updated following insights derived from relevant COPE cases and the industry standards).
Rescinded: Editorial Expression of Concern
23 June 2025: Editorial expression of concern issued on 18 May 2025 is hereby rescinded on account of author’s explanation / justification of the AI generated / paraphrased content. Author’s justification reads “We used AI for Language support only because as non-native English speakers, we have always faced challenges in achieving native-level fluency and clarity in our academic writing. We acknowledge that we used AI-based tools to polish and correct English grammar in a few portions of the manuscript, as highlighted by the AI detection report“. Readers may access rebuttal letter here. However, the board has decided to keep the contents of this article unchanged.
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: The targeted inhibition of Microtubule Affinity-Regulating Kinase 4 (MARK4) using small molecule inhibitors has emerged as a promising therapeutic approach for diverse diseases, including neurological disorders such as Alzheimer’s disease. Ongoing research endeavors aim to develop novel and more effective MARK4 inhibitors with enhanced target specificity and reduced off-target effects. In the present study, we sought to investigate the binding affinity and impact of Naringenin on the activity of MARK4.
Methods: We employed a combination of molecular docking and other bioinformatics methods, a fluorescence-based inhibition assay, as well as a kinase activity assay to assess the binding affinity and inhibition potential of Naringenin against MARK4. Additionally, we utilized the MTT assay to examine the effect of Naringenin on the viability of two cell lines: the normal human cell line HEK-293 and the neuronal cell line SH-SY5Y. The IC50 dose of Naringenin, determined from the MTT assay, provided a valuable reference point for subsequent neuronal cell line experiments.
Results: The results of the molecular docking demonstrated a robust binding affinity of Naringenin (-7.8 kcal/mol) to MARK4, affirming its potency as an inhibitor. Moreover, the fluorescence inhibition and kinase activity assays confirmed the inhibitory effect of Naringenin on MARK4. Interestingly, the MTT assay outcomes indicated that increasing concentrations of Naringenin did not significantly impact the viability of HEK 293 cells, while exhibiting a pronounced effect on SH-SY5Y neuronal cells. The IC50 concentration of Naringenin was determined to be 10.0 ± 1.33 μM for SH-SY5Y neuronal cells.
Conclusion: In conclusion, this study reported Naringenin as a potential inhibitor molecule for MARK4, offering promising prospects for future therapeutic interventions in neuronal disorders specifically for Alzheimer’s disease.
Keywords: Inhibition; Microtubule Affinity-Regulating Kinase 4 (MARK4); Molecular docking; Naringenin; Neuronal disorders
Introduction![]()
Microtubule Affinity Regulating Kinase 4 (MARK4) is a serine/threonine kinase that plays a crucial role in regulating cell polarity and microtubule dynamics [1]. MARK4 is a member of the AMP-activated protein kinase (AMPK) family and is highly conserved across species from worms to humans. Its pivotal role in cellular processes, such as cell division, migration, and differentiation, makes it an essential protein in maintaining cellular homeostasis and tissue integrity [2]. MARK4 consists of several functional domains, including a kinase domain, and ubiquitin-associated (UBA) domain, and a regulatory domain [3]. The kinase domain is responsible for the enzymatic activity of MARK4, while the UBA domain is involved in binding to ubiquitin and mediating interactions with other proteins. The regulatory domain serves as a platform for MARK4’s interaction with other molecules and is crucial for its subcellular localization and activity.
Activation of MARK4 is tightly regulated, and its activity is influenced by various factors, including phosphorylation, protein-protein interactions, and subcellular localization [4]. Phosphorylation of specific residues within the regulatory domain modulates the enzyme’s activity, and MARK4 is typically kept in an inactive state until it is activated by upstream kinases [5]. One of the prominent functions of MARK4 is its involvement in establishing and maintaining cell polarity [6]. MARK4 regulates cell polarity by phosphorylating various target proteins, such as microtubule-associated proteins (MAPs) and polarity regulators like Par-3 and Par-6 [7]. These phosphorylation events lead to the reorganization of the cytoskeleton and the establishment of polarized cell structures. In neurons, MARK4 is especially important for axon formation and dendritic branching, ensuring proper synaptic connectivity and neuronal function. MARK4 plays a central role in regulating microtubule dynamics by phosphorylating microtubule-associated proteins (MAPs) including tau protein and destabilizing microtubules in specific cellular regions [8]. Phosphorylation of MAPs by MARK4 promotes microtubule disassembly, leading to the generation of short, dynamic microtubule fragments [9]. These fragments can then be utilized for cellular processes requiring rapid remodeling of the cytoskeleton, such as cell migration and intracellular transport.
Due to its critical role in cell polarity and microtubule dynamics, dysregulation of MARK4 has been associated with various human diseases [10, 11]. MARK4’s involvement in tau phosphorylation has drawn significant attention due to its connection to neuronal diseases [11]. Neurodegenerative diseases, such as Alzheimer’s disease, are characterized by the abnormal accumulation of proteins, including tau, leading to neuronal dysfunction and cell death [12]. MARK4 is known to phosphorylate tau, contributing to the formation of neurofibrillary tangles, a hallmark of Alzheimer’s disease [13]. By targeting MARK4, researchers aim to reduce tau hyperphosphorylation, thereby potentially slowing down or preventing the progression of neurodegeneration. Therapeutic approaches that inhibit MARK4 could help restore microtubule stability and reduce neurotoxic tau aggregates, providing a potential disease-modifying treatment for Alzheimer’s disease and related disorders [14]. Apart from its role in neurodegenerative diseases, MARK4 has also been implicated in other cellular processes (Basheer et al., 2023). It is involved in the regulation of cell cycle progression, apoptosis (programmed cell death), and cellular response to stress [15]. Dysregulation of MARK4 activity has been associated with certain cancers, highlighting its importance in tumorigenesis and tumor progression [16].
Although much remains to be explored regarding the exact mechanisms and regulatory pathways involving MARK4, research efforts are ongoing to gain a comprehensive understanding of its functions and potential therapeutic implications. As with many other protein kinases, the development of specific inhibitors or modulators of MARK4 activity could hold promise for addressing various diseases such as Alzheimer’s disease where its upregulation plays a significant role.
Methods![]()
Target protein and ligand preparation
Initially, we obtained the 3D crystal structure of Microtubule Affinity-Regulating Kinase 4 (MARK4) from the Protein Data Bank using PDB ID 5ES1 as reference [17]. This structure was resolved at 2.80 Å and was in a complex with a pyrazolopyrimidine inhibitor molecule. Further, we retrieved the 2D structure of the ligand molecule, Naringenin, from DrugBank with Accession Number DB03467 (https://go.drugbank.com/drugs/DB03467). To ensure the target protein’s suitability for docking studies, a standard receptor preparation protocol was applied [18, 19]. We utilized the Swiss-PDB Viewer tool to perform energy minimization, which involved adjusting the atomic positions to release local constraints and remove the inhibitor molecule from the protein-inhibitor complex. During this process, we detected and rectified various issues, including missing side chains, atoms, or bonds, molecule-chain breaks, added water molecules, alternate locations, and others. For the subsequent docking using AutoDock Vina, we further prepared the target protein by adding appropriate polar hydrogens and assigning Kollman United Atom Charges. Additionally, both the target protein and ligand molecule were converted to the .pdbqt format, which is required for conducting molecular docking in AutoDock Vina.
Molecular docking
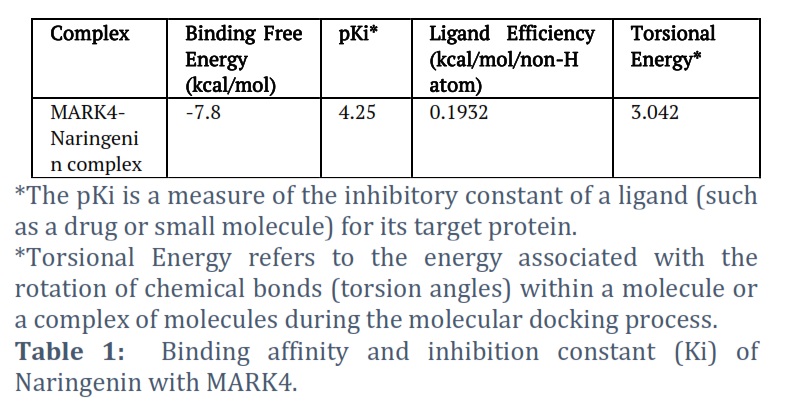
The molecular docking investigations were conducted utilizing AutoDock Vina and MGL tools, following the methodology outlined by Sultan et al. [18, 19]. The objective was to dock Naringenin scaffolds into the active sites of MARK4. For this purpose, a grid size with X, Y, and Z coordinates of 57, 84, and 73 Å, respectively, was established, centered at coordinates of −31.21, 14.93, and 71.51, respectively. The grid spacing was set to 1.00 Å, and the exhaustiveness parameter was configured to 8. Furthermore, the pKi, ligand efficiency (kcal/mol/non-H atom), and torsional energy for the MARK4-Naringenin complex were computed to gain additional insights into the binding interaction.
ADMET properties and biological activity prediction
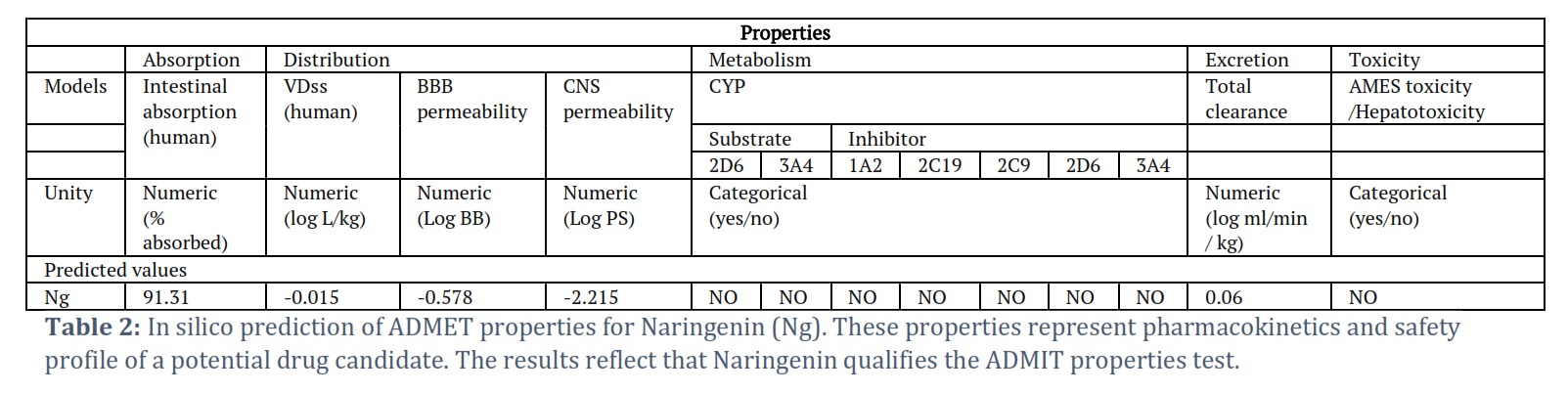
ADMET properties, sometimes referred to as ADME-Tox properties, are essential considerations in drug discovery and development. ADMET is an acronym that stands for Absorption, Distribution, Metabolism, Excretion, and Toxicity. These properties collectively help assess the pharmacokinetics and safety profile of a potential drug candidate. The pkCSM tool was utilized to evaluate the ADMET (Absorption, Distribution, Metabolism, Excretion, and Toxicity) properties of the chosen Naringenin molecule [20]. The SMILE ID of Naringenin was retrieved from the DrugBank database (https://go.drugbank.com/drugs/DB03467) and then uploaded to the online pkCSM server to determine its ADMET properties.
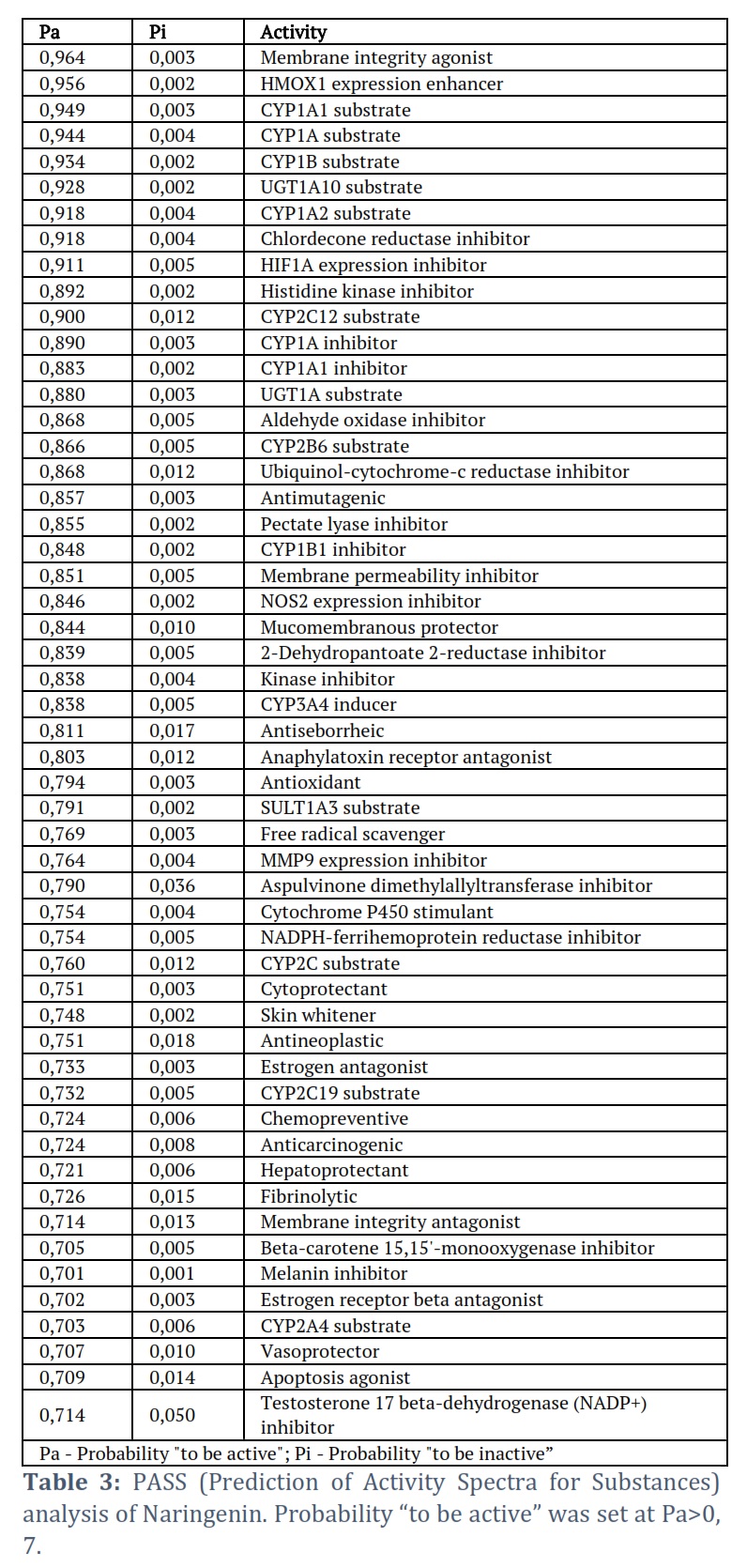
PASS (Prediction of Activity Spectra for Substances) is a computational tool used in cheminformatics and drug discovery to predict the biological activities of chemical compounds. It’s designed to estimate the potential pharmacological activities of compounds based on their chemical structure. PASS analysis relies on a large database of chemical structures and their known biological activities to make predictions for new or untested compounds. To conduct the PASS analysis, the chemical structure of Naringenin, presented in the standard “SMILES” format, was uploaded to the online server (http://www.pharmaexpert.ru/passonline/) for predicting the probability of biological activities associated with the compound.
MTT assay for cell viability
For this experiment, two cell lines, namely HEK-293 (normal cell line) and SH-SY5Y (neuronal cell line), were cultured in a 96-well microplate at a density of 5,000 cells per well in 100 µL of DMEM medium. The microplate was then placed in a CO2 incubator at 37°C with humidified conditions for 24 hours. After the 24-hour incubation period, the MTT reagent was prepared by dissolving 5 mg of MTT in 1 mL of sterile PBS. Next, the culture medium in the wells was replaced with 100 µL of fresh medium containing the MTT reagent at a final concentration of 0.5 mg/mL. The cells were further incubated for 4 hours at 37°C. Following the 4-hour incubation, the medium with the MTT reagent was removed from the wells, and 100 µL of DMSO was added to each well to dissolve the purple formazan crystals produced by the MTT reagent. To ensure complete dissolution, the microplate was gently shaken for 10 minutes. The absorbance of each well was then measured at 570 nm using a plate reader. To calculate the net absorbance, the absorbance of blank wells (containing only culture medium and MTT reagent) was subtracted from the absorbance of the test wells. The percentage of cell viability was determined by dividing the net absorbance of the test wells by the net absorbance of the control wells (cells without any treatment) and multiplying the result by 100.
In vitro inhibition assay
The recombinant human MARK4 protein, affinity purified to a level exceeding 90% on SDS-PAGE, was acquired from Abcam (ab105211). Naringenin was purchased from Sigma, USA (N5893; CAS No.: 67604-48-2). Fluorescence binding studies involving MARK4 and Naringenin were conducted using a JASCO 6300 spectrofluorometer with a 1 cm quartz cell. To generate the fluorescence spectrum of MARK4 and the MARK4-Naringenin complex, the following steps were executed: (1) Excitation of MARK4 was performed at a wavelength of 280 nm. (2) Emission from MARK4 was recorded in the range of 300-500 nm. (3) The excitation and emission slit widths were set to 10 mm, and the response was set to medium. Subsequently, the obtained fluorescence data from MARK4 and the MARK4-Naringenin complex were subjected to analysis. This analysis aimed to derive various binding parameters for the MARK4-Naringenin complex, shedding light on the inhibition of MARK4 by Naringenin.
Kinase assay
To investigate the impact of Naringenin on the activity of MARK4, an ATPase assay was conducted following the methodology described by Rule et al. [21]. This assay served to measure the activity of MARK4 by monitoring the release of inorganic phosphate (Pi) during the hydrolysis of ATP. In the experiment, different concentrations of Naringenin (ranging from 0 to 10 µM) were added to MARK4 (at a concentration of 2 µM), and the mixture was incubated for 1 hour at 25°C. Subsequently, freshly prepared ATP (200 µM) and MgCl2 (10 mM) were introduced into the reaction mixture, which was then incubated for an additional 30 minutes at 25°C. To halt the reaction, BIOMOL® reagent was added, leading to the formation of a green-colored complex. The absorbance of this complex was measured at 620 nm using a microplate reader. The ATPase assay employed Malachite green reagent (Biomol, Enzo Life Sciences, New York, NY, USA) as a crucial component.
Results![]()
Molecular docking revealed Naringenin as potential inhibitor of MARK4
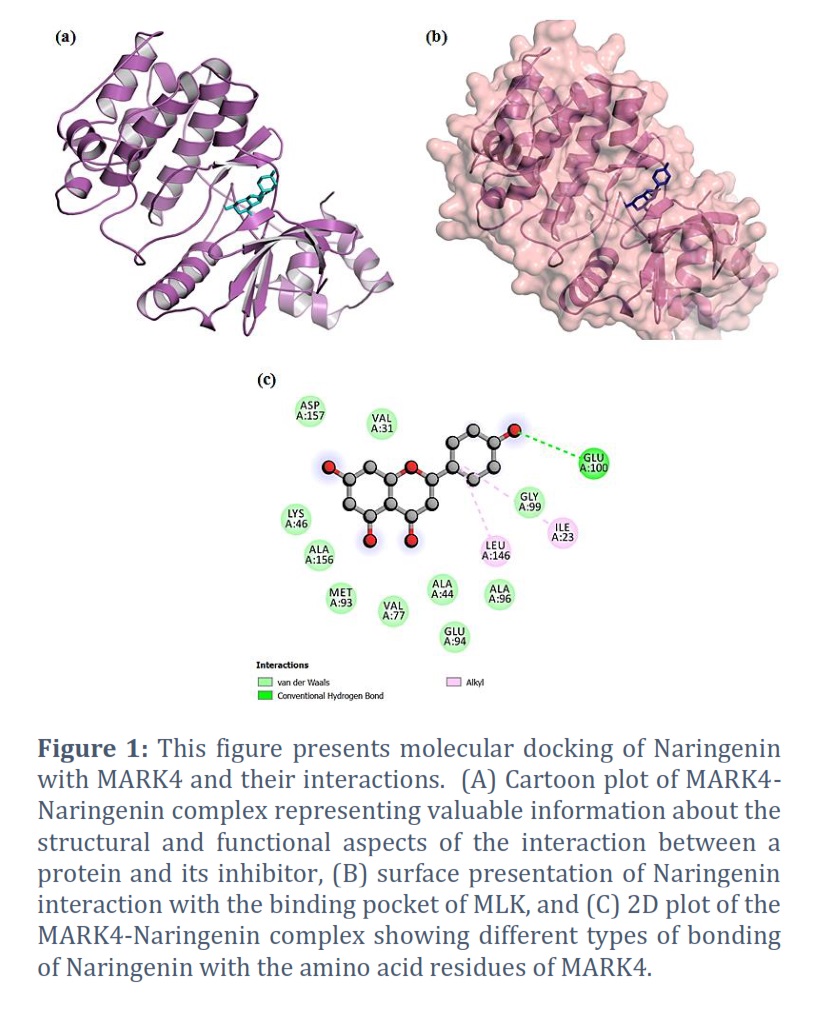
The molecular docking results of the MARK4-Naringenin complex are presented in Table 1 and Figure 1. The docking analysis yielded affinity scores and docked poses, revealing that Naringenin exhibited a notable binding affinity score of −7.8 kcal/mol with MARK4 (Table 1). The docking pose exhibited the successful docking of Naringenin into the deep binding cavity of MARK4 (Figure 1B). Furthermore, the docking results demonstrated a preferential hydrogen bond interaction of Naringenin with the GLU100 amino acid residue of the kinase domain of MARK4 (Figure 1C). Additionally, Naringenin formed several hydrophobic interactions with MARK4 (Figure 1C).
ADMET predictions and PASS analysis showed that Naringenin has drug like properties
Results of the ADMIT predictions are represented in Table 2. ADMET predictions indicated that Naringenin satisfies all requisite pharmacokinetic criteria. Therefore, Naringenin may be validated and developed as a therapeutic molecule to inhibit the enzymatic activity of the MARk4 associated with neuronal diseases. In addition, PASS Analysis revealed numerous favorable biological properties associated with Naringenin (Table 3). Results showed that Naringenin is associated with several biological properties including neuronal activities.
MTT assay showed a significant decrease in the viability of SH-SY5Y neuronal cells.
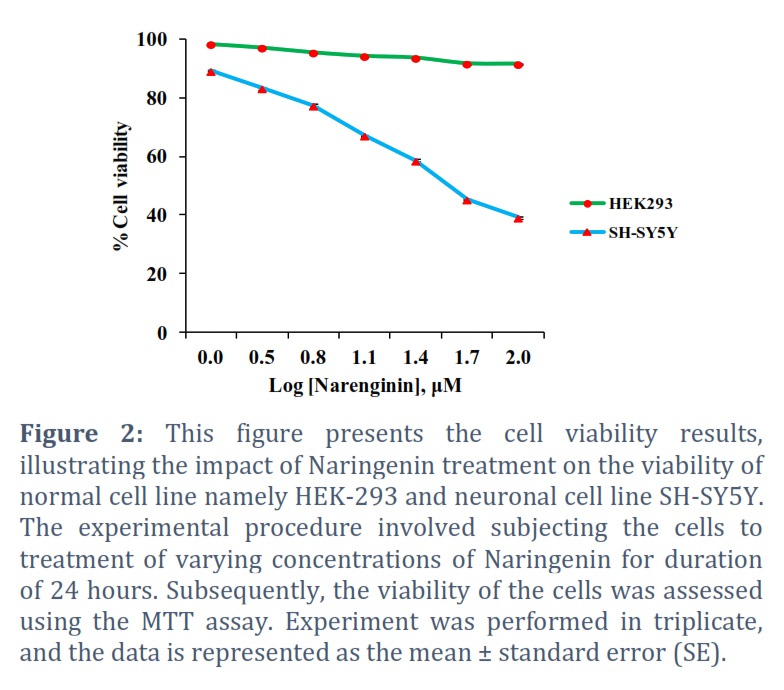
The outcomes of MTT assay analysis are presented in Figure 2. The MTT assay results revealed that the increasing concentrations of Naringenin did not exert a significant effect on the viability of HEK-293 cells. However, the viability of SH-SY5Y neuronal cells exhibited a noticeable and significant response to the treatment. There was a significant decrease in the viability of SH-SY5Y cells. Notably, the IC50 concentration of Naringenin was determined to be 10.0 ± 1.33 μM for SH-SY5Y cells.
Fluorescence-based binding assay and kinase assay revealed Naringenin as potential inhibitor of MARK4
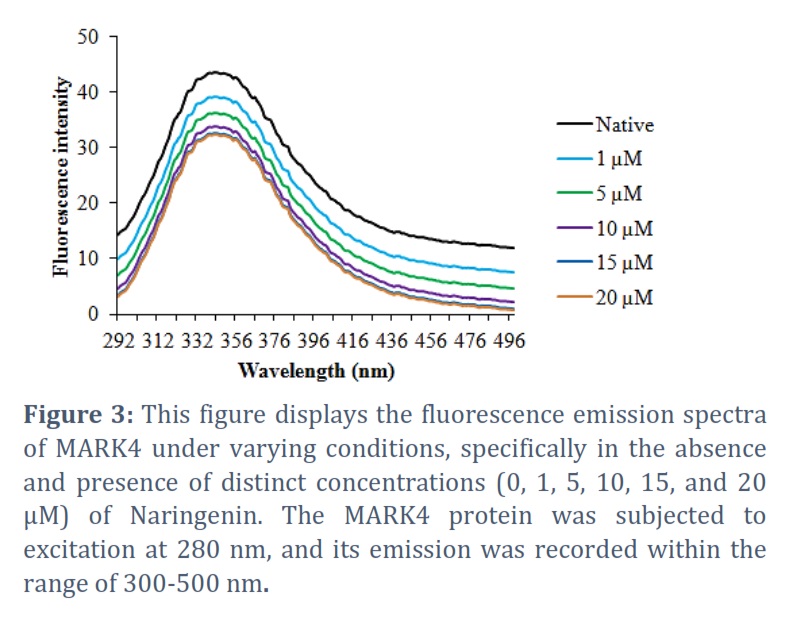
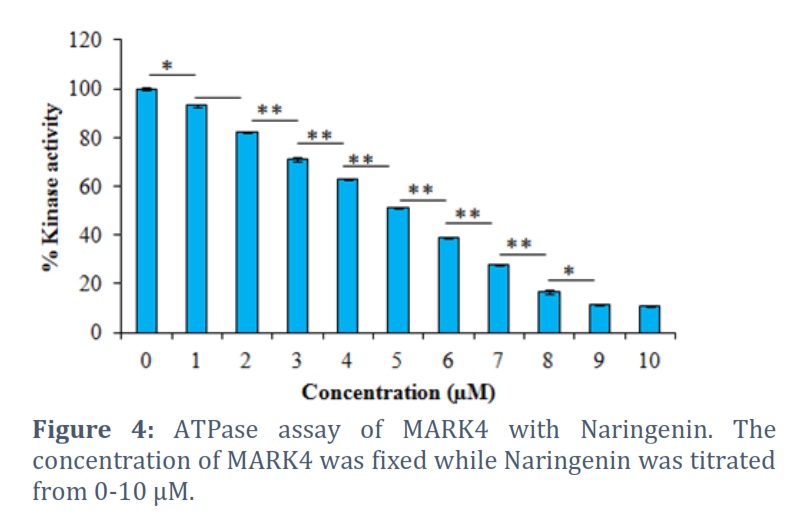
The obtained fluorescence emission spectra of MARK4 at various concentrations of Naringenin (ranging from 0 to 10 µM) are presented in Figure 3. Our analysis revealed that as the concentration of Naringenin increased, the fluorescence of MARK4 exhibited a corresponding decrease. This observation strongly indicates that Naringenin acts as an inhibitor of MARK4 by hindering its activity. The outcomes of kinase assay are graphically presented in Figure 4. Notably, the results demonstrate a pronounced reduction in the activity of MARK4 with the increasing concentration of Naringenin (Figure 4). These findings clearly indicated a noticeable reduction in the kinase activity of MARK4 with increasing concentrations of Naringenin, signifying its inhibitory effect on the enzyme.
Figures & Tables
Alzheimer’s disease is a progressive neurodegenerative disorder that primarily affects the elderly population. It is the most common cause of dementia and is characterized by cognitive decline, memory loss, impaired thinking, and behavioral changes [22]. Therapeutic strategies for Alzheimer’s disease are focused on slowing down disease progression, managing symptoms, and improving the quality of life for affected individuals. However, it is important to note that there is currently no cure for Alzheimer’s disease, and available treatments can only help manage symptoms to some extent. There is ongoing research to identify disease-modifying treatments for Alzheimer’s disease. These treatments aim to target the underlying pathology of the disease, such as the accumulation of amyloid plaques and tau tangles in the brain. MARK4 has emerged as a promising therapeutic target in the context of Alzheimer’s disease. This enzyme plays a crucial role in regulating microtubule dynamics, which are essential for maintaining neuronal structure and function [13]. Dysregulation of microtubule-associated processes has been implicated in Alzheimer’s disease pathogenesis, contributing to the formation of neurofibrillary tangles and neuronal degeneration [12]. By targeting MARK4, there is potential to modulate microtubule stability and potentially mitigate disease-related pathology. Thus, investigations into the inhibition or modulation of MARK4 present a compelling possibility for the development of novel therapeutic interventions in Alzheimer’s disease.
The current investigation focused on assessing the potential of Naringenin as an inhibitor of MARK4, employing a combination of in silico and in vitro approaches [18, 19]. Initially, molecular docking was utilized to predict the binding affinity between Naringenin and MARK4. The molecular docking results of the MARK4-Naringenin complex are presented in Table 1 and Figure 1. Our investigation involved molecular docking to elucidate the potential binding interaction between Naringenin and MARK4, and to explore their bonding characteristics, as well as determine their binding affinity [19]. The docking analysis yielded affinity scores and docked poses, revealing that Naringenin exhibited a notable binding affinity score of −7.8 kcal/mol with MARK4 (Table 1). Furthermore, the docking results demonstrated a preferential interaction of Naringenin with the binding pocket of MARK4. In-depth examination of the binding mode and interaction pattern of Naringenin with MARK4 was carried out using PyMOL. This analysis unveiled the formation of hydrogen bonds with the GLU100 residue of the kinase domain of MARK4 (Figure 1B). Additionally, Naringenin engaged in several hydrophobic interactions with MARK4. The docking pose exhibited the successful docking of Naringenin into the deep binding cavity of MARK4 (Figure 1C). Based on the comprehensive docking results, we posit that Naringenin holds promise as a potential binding molecule and inhibitor of MARK4. Furthermore, ADMET predictions indicated that Naringenin satisfies all requisite pharmacokinetic criteria (Table 2). Therefore, Naringenin may be validated and developed as a therapeutic molecule to inhibit the enzymatic activity of the MARk4 associated with neuronal diseases. In addition, PASS Analysis revealed numerous favorable biological properties associated with Naringenin (Table 3). Results showed that Naringenin is associated with several biological properties including neuronal activities. These findings further prioritize the Naringenin for further testing and optimization based on the PASS prediction results if the predicted biological activity is desirable. Collectively, these findings suggest the potential of Naringenin as a promising therapeutic agent for inhibiting the enzymatic activity of MARK4, particularly in the context of neuronal diseases like Alzheimer’s disease.
Next, we investigated the impact of Naringenin on the viability of HEK-293 and SH-SY5Y cells. We used MTT assay (3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide assay), a commonly used colorimetric assay to measure cell viability and assess cell proliferation or cytotoxicity [23]. It is widely used in biological and biomedical research to evaluate the effects of various compounds, drugs, or treatments on cell cultures. The cells were exposed to various concentrations of Naringenin ranging from 0 to 100 μM. The outcomes of this analysis are presented in Figure 2. The MTT assay results revealed that the increasing concentrations of Naringenin did not exert a significant effect on the viability of HEK-293 cells. However, the viability of SH-SY5Y neuronal cells exhibited a noticeable and significant response to the treatment. There was a significant decrease in the viability of SH-SY5Y cells. Notably, the IC50 concentration of Naringenin was determined to be 10.0 ± 1.33 μM for SH-SY5Y cells.
Following the confirmation of Naringenin’s binding to MARK4 through computer simulations, we employed a fluorescence-based binding assay to quantify the strength of the interaction between the two entities. It is a fluorescence-based binding assay that is a valuable technique for assessing ligand binding affinity [24]. It serves as a useful tool for screening ligands that may exhibit inhibitory effects on target proteins [25]. This assay relies on measuring the changes in protein fluorescence upon binding of the ligand (Naringenin) to MARK4. The obtained fluorescence emission spectra of MARK4 at various concentrations of Naringenin (ranging from 0 to 10 µM) are presented in Figure 3. Our analysis revealed that as the concentration of Naringenin increased, the fluorescence of MARK4 exhibited a corresponding decrease. This observation strongly indicates that Naringenin acts as an inhibitor of MARK4 by hindering its activity. These findings suggest the potential potency of Naringenin as a robust inhibitor of MARK4, leading us to further investigate its inhibitory effects using the ATPase assay.
For further investigation into the functional behavior of MARK4, we employed a kinase assay. Kinases are enzymes that facilitate the transfer of phosphate groups from ATP (adenosine triphosphate) to specific substrates, such as proteins or other small molecules [26]. The study of kinase activity is crucial for comprehending various cellular processes and signaling pathways, as kinases play a pivotal role in regulating numerous cellular functions, including cell growth, differentiation, and metabolism. A kinase assay was performed, assessing the impact of various concentrations of Naringenin on MARK4 activity. The outcomes of this assay are graphically presented in Figure 4. The baseline activity of MARK4 without Naringenin was set at 100%. Notably, the results demonstrate a pronounced reduction in the activity of MARK4 with the increasing concentration of Naringenin (Figure 4). These findings clearly indicated a noticeable reduction in the kinase activity of MARK4 with increasing concentrations of Naringenin, signifying its inhibitory effect on the enzyme.
In conclusion, the present investigation provides evidence for the potential of Naringenin as a promising MARK4 inhibitor. The combination of in silico and in vitro methodologies demonstrated a high binding affinity between Naringenin and MARK4, with favorable pharmacokinetic properties and biological attributes, thereby implying its plausible therapeutic utility in diseases, notably Alzheimer’s disease, where MARK4 dysregulation is implicated. Nevertheless, it is imperative that additional investigations be undertaken to assess the efficacy and safety of Naringenin as a MARK4 inhibitor, as well as to refine its dosage and delivery methods to align with clinical requirements.
Acknowledgment
The authors extend their appreciation to the Deputyship for Research & Innovation, Ministry of Education in Saudi Arabia for funding this research work through project number IFPRC-168-141-2020 and King Abdulaziz University, DSR, Jeddah, Saudi Arabia.
Conflict of Interest
The authors declare that there is no conflict of interest.
MZA and BSA conceived the idea, HAB and MAFZ collected and analyzed computational data, TMAA, MZA, and GMA conducted in vitro experiments, MZA, MK, and GMA wrote the manuscript.
![]() References
References
- Drewes G, Ebneth A, & Mandelkow E. MMAPs, MARKs and microtubule dynamics. Trends in biochemical sciences, (1998); 23(8): 307-311.
- Chudobová J, & Zempel H. Microtubule affinity regulating kinase (MARK/Par1) isoforms differentially regulate Alzheimer-like TAU missorting and Aβ-mediated synapse pathology. Neural regeneration research, (2023); 18(2): 335-336.
- Timm T, Marx A, Panneerselvam S, Mandelkow E, & Mandelkow EM. Structure and regulation of MARK, a kinase involved in abnormal phosphorylation of Tau protein. BMC neuroscience, (2008); 9 Suppl 2(Suppl 2): 1471-2202.
- Trinczek B, Brajenovic M, Ebneth A, & Drewes G. MARK4 is a novel microtubule-associated proteins/microtubule affinity-regulating kinase that binds to the cellular microtubule network and to centrosomes. The Journal of biological chemistry, (2004); 279 (7):5915-5923.
- Rovina D, Fontana L, Monti L, Novielli C, Panini N, Sirchia SM, Erba E, Magnani I, & Larizza L. Microtubule-associated protein/microtubule affinity-regulating kinase 4 (MARK4) plays a role in cell cycle progression and cytoskeletal dynamics. European journal of cell biology, (2014); 93(8-9): 355-365.
- Timm T, Von Kries JP, Li X, Zempel H, Mandelkow E, & Mandelkow EM. Microtubule affinity regulating kinase activity in living neurons was examined by a genetically encoded fluorescence resonance energy transfer/fluorescence lifetime imaging-based biosensor: inhibitors with therapeutic potential. The Journal of biological chemistry, (2011); 286(48): 41711-41722.
- Tang EI, Mruk DD, & Cheng CY. MAP/microtubule affinity-regulating kinases, microtubule dynamics, and spermatogenesis. The Journal of endocrinology, (2013); 217(2): R13-R23.
- Barbier P, Zejneli O, Martinho M, Lasorsa A, Belle V, Smet-Nocca C, Tsvetkov PO, Devred F, & Landrieu I. Role of Tau as a Microtubule-Associated Protein: Structural and Functional Aspects. Frontiers in aging neuroscience, (2019); 11: 204.
- Ebneth A, Drewes G, Mandelkow EM, & Mandelkow E. Phosphorylation of MAP2c and MAP4 by MARK kinases leads to the destabilization of microtubules in cells. Cell motility and the cytoskeleton, (1999); 44 (3): 209-224.
- Liu Z, Gan L, Chen Y, Luo D, Zhang Z, Cao W, Zhou Z, Lin X, & Sun C. Mark4 promotes oxidative stress and inflammation via binding to PPARγ and activating NF-κB pathway in mice adipocytes. Scientific reports, (2016); 6: 21382.
- González A, Singh SK, Churruca M, & Maccioni RB. Alzheimer’s Disease and Tau Self-Assembly: In the Search of the Missing Link. International journal of molecular sciences, (2022); 23(8): 4192.
- Oba T, Saito T, Asada A, Shimizu S, Iijima KM, & Ando K. Microtubule affinity-regulating kinase 4 with an Alzheimer’s disease-related mutation promotes tau accumulation and exacerbates neurodegeneration. The Journal of biological chemistry, (2020); 295(50): 17138-17147.
- Basheer N, Smolek T, Hassan I, Liu F, Iqbal K, Zilka N, & Novak P. Does modulation of tau hyperphosphorylation represent a reasonable therapeutic strategy for Alzheimer’s disease? From preclinical studies to the clinical trials. Molecular psychiatry, (2023); 28(6):2197-2214.
- Ballatore C, Lee VMY, and Trojanowski JQ, Tau-mediated neurodegeneration in Alzheimer’s disease and related disorders. Nature Reviews Neuroscience, (2007); 8(9):663-72.
- Wang L, Pu W, Wang C, Lei L, & Li H. Microtubule affinity regulating kinase 4 promoted activation of the NLRP3 inflammasome-mediated pyroptosis in periodontitis. Journal of oral microbiology, (2021); 14(1): 2015130.
- Shen X, Liu X, Wan S, Fan X, He H, Wei R, Pu W, Peng Y, & Wang C. Discovery of Coumarin as Microtubule Affinity-Regulating Kinase 4 Inhibitor That Sensitize Hepatocellular Carcinoma to Paclitaxel. Frontiers in chemistry, (2019); 7: 366.
- Sack JS, Gao M, Kiefer SE, Myers JE, Jr, Newitt JA, Wu S, & Yan C. Crystal structure of microtubule affinity-regulating kinase 4 catalytic domain in complex with a pyrazolopyrimidine inhibitor. Acta crystallographica. Section F, Structural biology communications, (2016); 72(Pt 2): 129-134.
- Sultan A, Ali R, Sultan T, Ali S, Khan NJ, & Parganiha A. Circadian clock modulating small molecules repurposing as inhibitors of SARS-CoV-2 Mpro for pharmacological interventions in COVID-19 pandemic. Chronobiology International, (2021); 38(7): 971-985.
- Sultan A, Ali R, Ishrat R, & Ali S. Anti-HIV and anti-HCV small molecule protease inhibitors in-silico repurposing against SARS-CoV-2 Mpro for the treatment of COVID-19. Journal of Biomolecular Structure and Dynamics, (2022); 40(23): 12848-12862.
- Ouassaf M, Belaidi S, Mogren Al Mogren M, Chtita S, Ullah Khan S, & Thet Htar T. Combined docking methods and molecular dynamics to identify effective antiviral 2, 5-diaminobenzophenonederivatives against SARS-CoV-2. Journal of King Saud University. Science, (2021); 33(2): 101352.
- Rule CS, Patrick M, & Sandkvist M. Measuring In Vitro ATPase Activity for Enzymatic Characterization. J Vis Exp, (2016); (114): 54305.
- DeTure MA, & Dickson DW. The neuropathological diagnosis of Alzheimer’s disease. Molecular neurodegeneration, (2019); 14(1): 32.
- Grela E, Kozłowska J, Grabowiecka. Current methodology of MTT assay in bacteria – A review. Acta Histochemica, (2018); 120(4): 303-311.
- Gochin M, Savage R, Hinckley S, & Cai L. A fluorescence assay for rapid detection of ligand binding affinity to HIV-1 gp41. Biological chemistry, (2006); 387(4): 477-483.
- Bragina ME, Stergiopulos N, & Fraga-Silva RA. Fluorescence-Based Binding Assay for Screening Ligands of Angiotensin Receptors. Methods in molecular biology (Clifton, N.J.), (2017); 1614: 165-174.
- Haubrich BA, & Swinney DC. Enzyme Activity Assays for Protein Kinases: Strategies to Identify Active Substrates. Current drug discovery technologies, (2016); 13(1): 2-15.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0