

Full Length Research Article
Exploring Genomic Patterns to Identify Drug-Resistant TB: A Comprehensive Study of Age, Gender, Lineage, and Outcome
Anam Tariq*, Sahar Fazal
Adv. life sci., vol. 11, no. 2, pp. 354-361, May 2024
*– Corresponding Author: Anam Tariq (dbi163002@cust.pk)
Authors' Affiliations
[Date Received: 25/06/2023; Date Revised: 13/09/2023; Date Available Online: 18/04/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Tuberculosis (TB) is a prevalent bacterial infection caused by Mycobacterium tuberculosis (Mtb), primarily affecting the lungs, with rising incidence rates. Drug-resistant TB strains, including multidrug-resistant (MDR) and extensively drug-resistant (XDR) types, pose challenges with poor treatment success rates and increased mortality. Demographic factors and genomic patterns contribute to drug resistance, emphasizing the need for new drugs targeting specific genomic patterns.
Methods: This study analyzes the impact of demographics, treatment outcomes, and genomic mutations on drug-resistant TB using pattern identification techniques and whole-genome sequencing to discover therapeutic targets. This study primarily focused on the identification and analysis of MDR and XDR strains, considering various demographic characteristics and treatment outcomes. A dataset comprising 2,602 observations was utilized, and pattern recognition techniques were employed to identify significant features. Additionally, the study employs the whole-genome sequencing (WGS) pipeline to identify single nucleotide polymorphisms (SNPs) that result in new mutations in drug-resistant Mtb strains.
Results: The findings of the study revealed that XDR and MDR non-XDR TB were the most prevalent types of drug resistance, and they were associated with unfavorable treatment outcomes, including death or treatment failure. Males exhibited a higher susceptibility to both XDR and MDR non-XDR TB compared to females. The age of onset for both types of resistance was approximately 40 years. Among the observed variants in 31 commonly occurring genes,
Conclusion: These findings introduce a novel set of therapeutic targets specific to MDR and XDR Mtb types, which warrant further investigation for potential therapeutic interventions.
Keywords: Tuberculosis; Drug Resistance; Multidrug resistance; Demographics; Dataset; Gene; Mutations
Introduction![]()
Tuberculosis (TB) is one of the oldest bacterial infections caused by Mycobacterium tuberculosis (Mtb), with high prevalence among underdeveloped populations of the world [1]. According to the WHO’s Global TB Report 2022, 10.6 million people were diagnosed with tuberculosis in 2021 with a rise in the incidence rate of TB as compared to the past years [2]. Bacterial antibiotic resistance in Mtb is the hardest hurdle in treatment of this deadly disease Rifampicin (RIF) and isoniazid (INH) are two first line drugs used against TB for centuries; however, RIF and INH resistance is a public health dilemma that occurs due to the mutated rpoB and katG genes respectively. This condition leads resistant to the two most potent drugs i.e. rifampin and isoniazid and is known as Multi drug resistance MDR. Second line inject able drugs known as fluoroquinolones are used to treat such cases by preventing the synthesis of bacterial DNA. However, resistance of TB isolate against rifampin, isoniazid, and fluoroquinolones corresponds to XDR which is the most dangerous version of MDR linked with an increased mortality rate [3].
To get a deeper insight into the underlying genomic patterns associated with drug-resistant TB, various demographic features majorly lineage, outcome, age, gender, and type of resistance serve as key determinants [4].
This can be facilitated through exploratory data analysis (EDA) which uses pattern recognition as a fundamental activity to search for the patterns and trends in the isolated samples, in order to shortlist them based on their clinical features followed by visual inspection of the selected features [5]. Moreover, it will provide an insight for determining the antimicrobial resistance (AMR) genes that are specific to each subtype of the isolated samples or strains of Mtb, in turn helping to investigate novel anti-tubercular therapeutic biomarkers that can be targeted to each specific subtype of resistant Mtb strain in TB patients [6]. The purpose of this study is to apply pattern identification techniques to a clinical dataset of Mtb in order to analyze the impact of gender, age of onset, lineage, type of resistance, and treatment outcomes on the identification of specific types of drug resistance associated with treatment failure. Additionally, the study employs the whole genome sequencing (WGS) pipeline to identify single nucleotide polymorphisms (SNPs) that result in new mutations in drug-resistant Mtb strains, specifically those leading to multidrug-resistant and extensively drug-resistant types. This analysis aims to discover novel therapeutic targets that can effectively combat these resistant and pathogenic strains of Mtb, particularly MDR and XDR types.
Methods![]()
In this study, dataset retrieved from TB portal was analyzed on the basis of multiple features to search for the existing patterns by utilizing pattern identification and datamining techniques. These patterns are further used to shortlist pathogenic isolates of XDR and MDR (non-XDR) types of TB by employing WGS pipeline in order to identify novel antimicrobial resistant (AMR) genes that are specific to MDR and XDR strains of TB from United States National Institute of Allergy and Infectious Diseases (NIAID) TB Portals database.
Data collection and per-processing
The genomic data (2012-2021) was taken from TB portals which is an open-access, web-based platform for global drug resistant tuberculosis data sharing and analysis. The dataset contained demographic data including age, gender, age of onset, type of resistance, outcome, and lineage and accession no of NCBI accession number of genomic sequence of the pathogen. The csv file contains a total number of 2,602 observations and 19 different features against each observation. Subsequently, 1,140 observations were initially subset by omitting those which showed sensitivity towards the drugs and the ones that were cured. Afterwards, the dataset was further subset according to the most common types of resistance (XDR, MDR non-XDR) found among all the samples. After this sub-setting, we obtained 1,050 observations that were next used for further analysis.
Pattern identification
Data analysis and visualization techniques were used for the identification of underlying patterns in the tuberculosis dataset containing several features including gender, age of onset, type of resistance, outcome, and lineage. This step was carried out using a customize python script which utilized matlib and seaborn packages which are the basic tool for explanatory data analysis. To understand the overall distribution of aforementioned clinical descriptors within the dataset, each descriptor was first visualized separately by implementing data visualization techniques. Subsequently, some patterns and trends were obtained by the set of combinations, involving any two distinct features. Depending on the observed patterns, age of onset was compared with outcome of the disease, type of resistance, and lineage by using box plots to determine which age group has majorly faced the worst outcome, which type of bacterial resistance was responsible, and how lineage was being influenced in various age groups respectively. Similarly, gender distribution was compared with the treatment outcome, type of resistance, and lineage by utilizing the parameter hue. This was done by plotting bar chart plots to identify how gender (male, female) was being influenced by the outcome of disease, how resistance of bacteria was affecting both genders, and how different lineages were affecting each gender. Moreover, through bar charts, the type of resistance was first compared with the disease outcome to understand how resistance of the bacteria influences the outcome of disease followed by its comparison with the lineage in order to analyze which lineage is significantly being affected by the bacterial resistance. Furthermore, most frequent types of resistance (XDR, MDR non-XDR) were then visualized in combination with gender, age, and outcome of the disease.
Identification of Novel AMR genes in Multi and Extensively Drug-Resistant Mtb
ARIBA [7] is a tool used to detect Antimicrobial Resistance (AMR) genes by analyzing paired read data. The AMR database creates a cluster of reference sequences by similarity using CD-HIT [8]. A set of reads for each cluster is generated by mapping the input reads to the reference sequences using the mini map. The reads are assembled within each cluster using Fermi-lite, and the resulting contigs are compared to the reference sequences using nucmer, which is part of the MUMmer tool package. Any variations or differences between the assembled contigs and the reference sequences are identified by nucmer. Bowtie2 [9] and SAM tools [10] are also used in ARIBA. Bowtie2 is responsible for mapping the reads for clusters to the assembly, and SAM tools are used to call the variants. Finally, a comprehensive report indicates the presence or absence of variants for all sequences in the analyzed samples [7]. For further analysis, our particular focus was on novel coding gene variants that may be associated with antimicrobial resistance in both types of Mtb mentioned previously.
Additionally, using custom Python scripts, comparative analyses were conducted on gene clusters identified by ARIBA within each sample of XDR and MDR strains. This process revealed common novel variants for both XDR and MDR strains. The identified common and unique novel variants were subsequently employed for further analysis.
Results![]()
Identification of multi drug resistant isolates w.r.t age, gender, and outcome
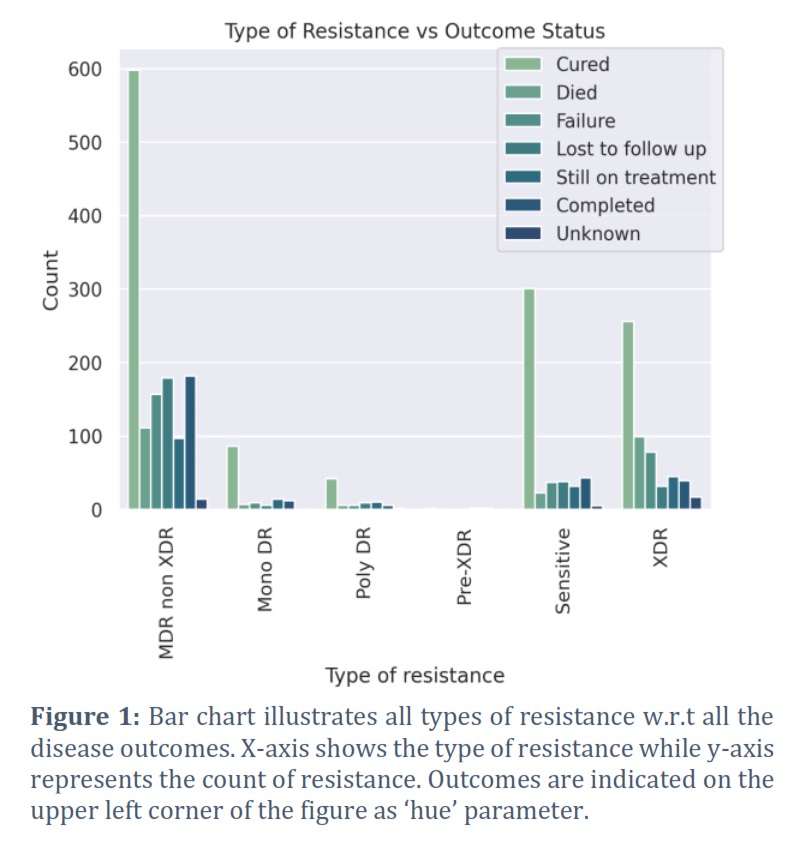
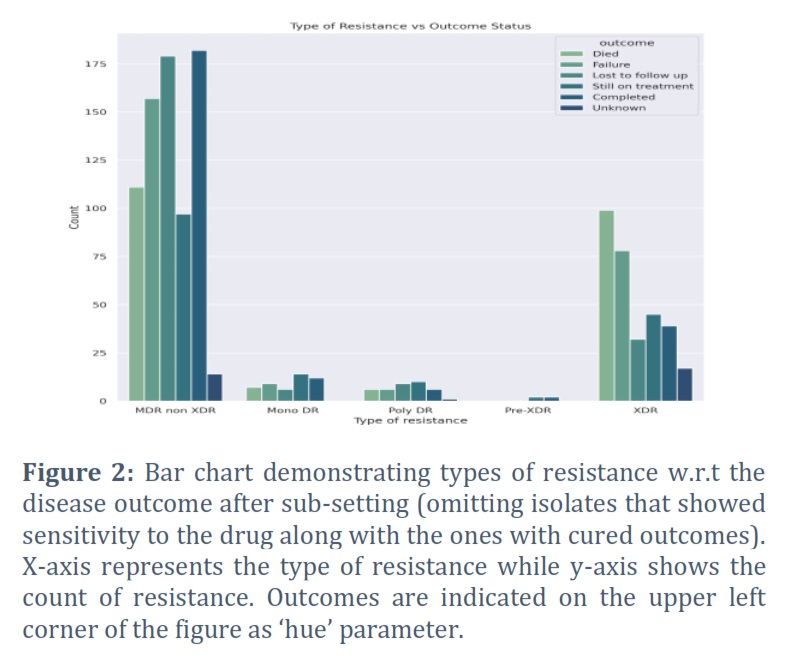
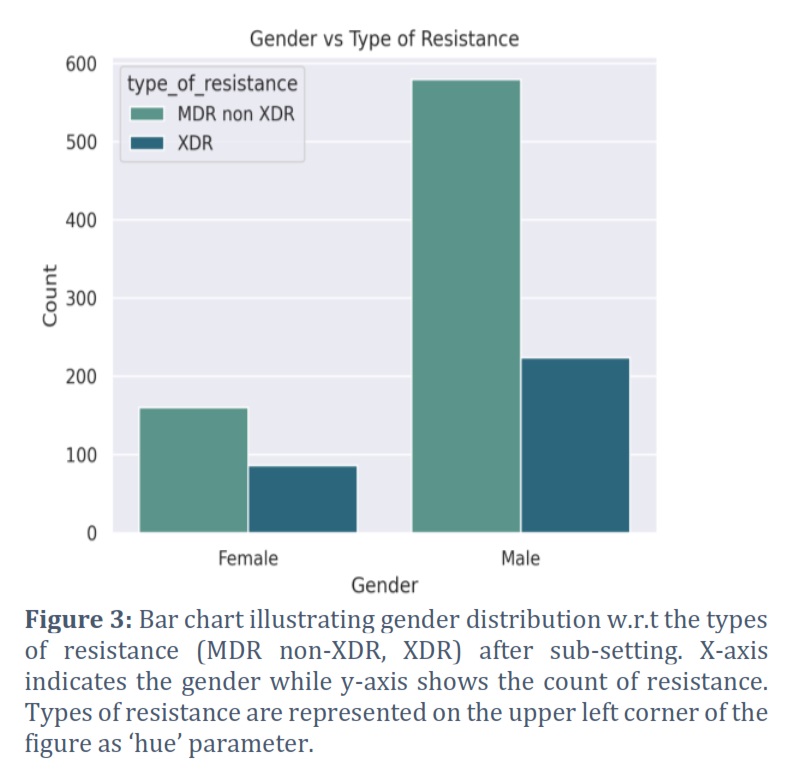
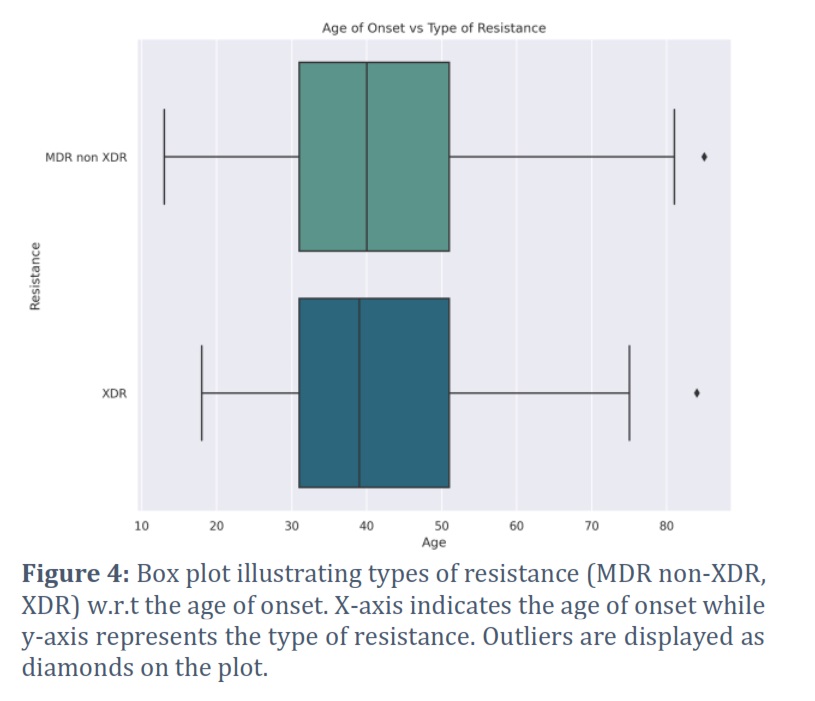
In order to identify drug resistant isolates of TB, various demographic features present in the dataset (2,602 observations) were analyzed and shortlisted by using pattern recognition techniques. Type of resistance which refers to the resistance exhibited by TB strain, age of onset which corresponds to the age at which TB was diagnosed, gender of the patient, and final outcome of the patient's treatment were the selected features as they were identified with some prominent patterns and regularities in the data. Furthermore, to better understand how different types of bacterial resistance influence outcome of the disease, all the types of resistance (pre-XDR, XDR, sensitive, mono DR, poly DR, and MDR non XDR) were visualized w.r.t all of the disease outcomes (cured, died, failure, lost to follow up, still on treatment, completed, and unknown) using a bar chart, as shown in Figure 1. As we were not concerned with those isolates which showed sensitivity towards the drugs and those which were cured, the dataset was initially subset accordingly and then plotted again by filtering out such isolates (Figure 2). It can be clearly visualized that XDR and MDR non-XDR are the most frequent TB resistance among all the isolates, linked to the worst outcomes. Subsequently, isolates with XDR and MDR non-XDR were separately plotted with outcome of the disease to visualize and analyze outcomes associated with these types of resistance by bar chart plot (Figure 4). After final subsetting of the dataset, it can be observed that more than 100 isolates having MDR non-XDR TB died while more than 150 isolates were associated with treatment failure. However, the count of XDR TB is comparatively less than MDR non-XDR TB. Around 75 determinants of treatment failure were observed in patients with XDR TB whereas around 98 isolates were of those patients which died with XDR TB. Moreover, gender distribution was also checked w.r.t the type of resistance to indicate which gender is predominantly being affected by both XDR and MDR non-XDR TB. It can be analyzed from the bar chart that males are significantly being influenced by both types of resistance, as compared to females. XDR TB is present in around 220 males while the count of females is less than 100. However, around 570 males were found with MDR non-XDR TB, whereas only 160 females possessed MDR non-XDR TB. Additionally, the patient's age of onset was then compared with the resistance of bacteria by visualizing through a comparative box plot (Figure 4). This was done to analyze the onset age distribution among the isolates of the patient having MDR non-XDR and XDR as the types of resistance. Age around 40 is the median, corresponding to both MDR non-XDR and XDR TB. Moreover, the age of patients with XDR TB ranges from 18-75 years while the age for MDR non-XDR TB patients ranges from 13-82 years. Majorly, both types of resistance range from ages between 32-52 years as demonstrated by the box plot. The rest of the graphs are provided as supplementary files.
Identification of antimicrobial resistance genes:
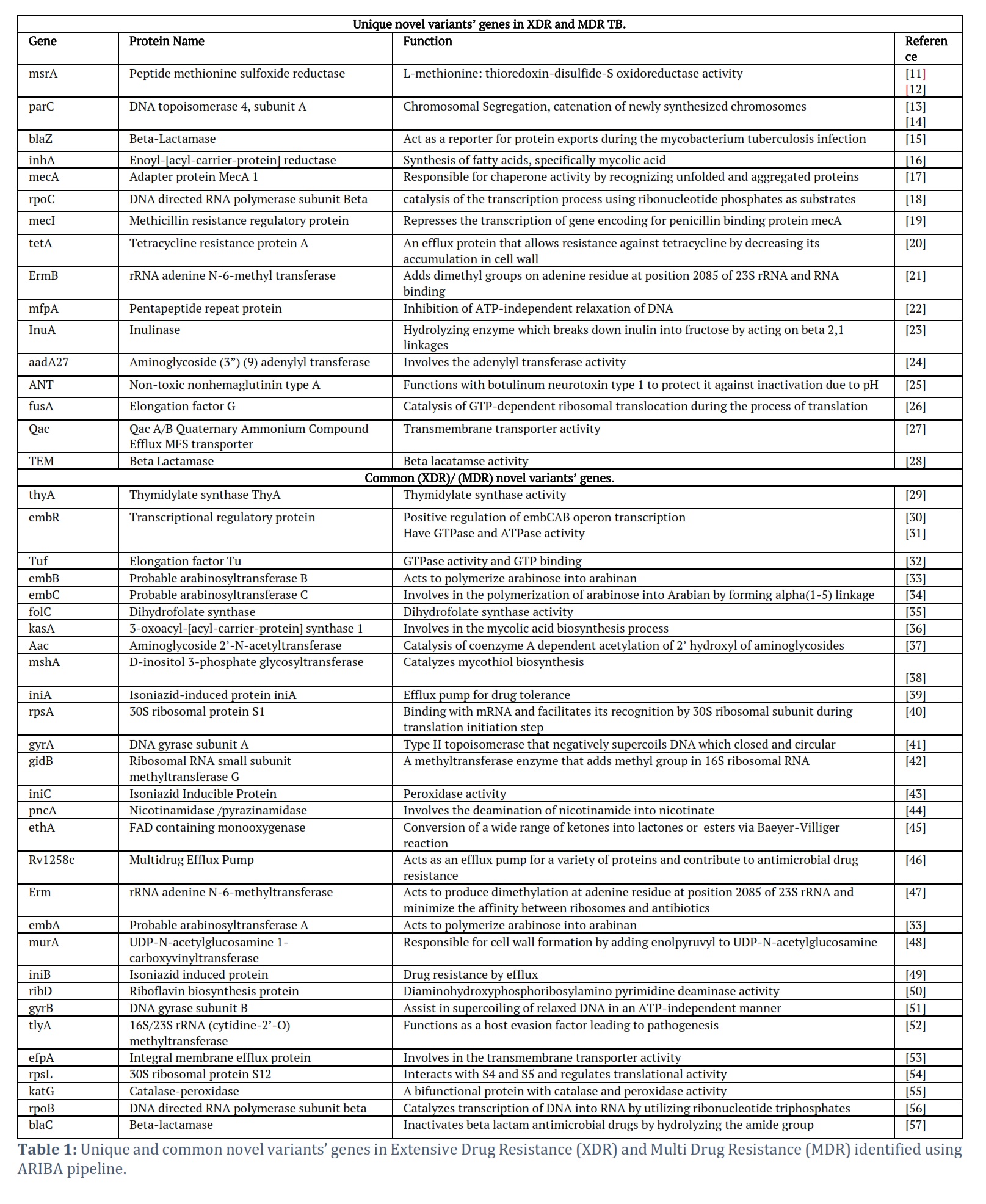
The majority of the cases of MDR and XDR were around age 40 (Figure 4) despite the gender. Utilizing XDR and MDR isolates samples, an investigation was conducted to identify the genetic elements responsible for antibiotic resistance using the whole genome sequencing pipeline, Ariba. This analysis unveiled previously known and newly discovered variants of these resistance genes. The novel variants were specifically selected for further scrutiny, identifying 47 unique variants in MDR samples and 36 unique variants in XDR samples. A comparative analysis was also performed to identify the shared novel variants in XDR and MDR novel resistance genes, revealing a set of unique variants exclusive to each category. Notably, one unique novel variant was exclusively found in the XDR samples, while 16 were exclusively identified in the MDR samples, Moreover, 31 variants were common between the XDR and MDR samples as given in Table 1.
Figures & Tables
The present study aimed to identify and analyze multi drug resistant isolates of TB based on various demographic features, including type of resistance, age of onset, gender, and treatment outcome. A dataset comprising 2,602 observations was subjected to pattern recognition techniques to identify relevant features. The types of resistance, such as pre-XDR, XDR, sensitive, mono DR, poly DR, and MDR non-XDR, were visualized in relation to different treatment outcomes using bar charts. By focusing on the most severe cases, the dataset was sub-set to exclude sensitive and cured isolates. The analysis revealed that XDR and MDR non-XDR were the most prevalent types of TB resistance and were associated with unfavorable outcomes.
Specifically, a significant number of patients with MDR non-XDR TB experienced death or treatment failure. In terms of gender distribution, males were more affected by both XDR and MDR non-XDR TB compared to females. Gender does not appear significant difference on drug resistance tuberculosis. Numerous studies have reported that females had a higher risk than males for MDR-TB, but our study did not find any significant difference between the sexes in this regard [58,59]. Analysis in one of the study showed that there was no evidence of an association between sex and risk of MDR/RR-TB in TB patients both globally and nationally in the majority (81%, 86 out of 106) of countries, with an overall random-effects weighted M:F risk ratio of 1.04 (95% CI 0.97–1.11) [60]. Age of onset analysis through box plots demonstrated that the median age for both types of resistance was around 40, with the age range varying slightly between XDR and MDR non-XDR TB cases. A case-control study conducted in Bangladesh confirmed this finding in the 25-44 age group, among whom MDR-TB was significantly more common [61]. Similarly, it has been found in another study that (63.5%) of the MDR-TB cases were from 15 to 44 years of age and were marginally statistically associated with MDR-TB [62]. Several studies showed the absence of statistically significant difference in the proportion of any resistance by age [63]. Supplementary files provide additional graphs for reference. These findings contribute to understanding the demographic patterns and outcomes associated with different types of drug-resistant TB. The shortlisted XDR and MDR strains were then analyzed through ARIBA pipeline to identify novel variants in each isolate.
The study's significant findings shed light on the novel variants associated with MDR and XDR strains, potentially playing a crucial role in tuberculosis drug resistance. The investigation successfully identified novel variants specifically linked to MDR and XDR, suggesting their direct or indirect involvement in the development of drug resistance. A total of 31 common genes with novel variants were observed in MDR and XDR samples. One gene was exclusive to XDR strains and 16 genes were exclusive to MDR strains.
In this study, multidrug-resistant isolates of tuberculosis (TB) were analyzed based on various demographic features, including resistance type, age of onset, gender, and treatment outcome. The analysis of a dataset comprising 2,602 observations revealed that extensively drug-resistant (XDR) and multidrug-resistant non-XDR (MDR non-XDR) TB were the most prevalent types of resistance and were associated with unfavorable treatment outcomes. Males were more affected by XDR and MDR non-XDR TB compared to females. The age of onset for both resistance types was around 40, with a slight variation between XDR and MDR non-XDR TB cases. Additionally, the study identified novel genetic variants associated with MDR and XDR strains of Mtb. Novel variants associated with MDR and XDR strains were identified, indicating their potential role in tuberculosis drug resistance. Among these variants, 31 common genes with novel variants were found in both MDR and XDR samples. The TEM gene was exclusive to XDR strains, and 16 genes were exclusive to MDR strains. Genes such as rpoB, rpoC, rpoZ, rpoA which have shown relevance to Mtb drug resistance and have gained novel variants, should be studied further. However, mutations at codon 864, 887, and 898 (V864I, Q887K, and A898T) were identified in rpoC which are novel variants. The results highlight potential therapeutic targets for controlling antimicrobial resistance in Mtb. However, further investigations are needed to fully understand the mechanisms and implications of these novel variants in drug resistance.
Conflict of Interest
The authors declare that there is no conflict of interest.
Acknowledgment
The data used in this research was taken from TB portals. The TB Portals website is freely available at https://tbportals.niaid.nih.gov/.
SF conceived the idea, AT and SF designed the experiments and AT carried out the computations. Both authors have accepted responsibility for the entire content of this manuscript and approved its submission.
![]() References
References
- Natarajan A, Beena PM, Devnikar AV, Mali S. A systemic review on tuberculosis. Indian Journal of Tuberculosis, (2020); 67(3): 295-311.
- Bagcchi S. WHO's global tuberculosis report 2022. The Lancet Microbe, (2023); 4(1):e20.
- Gill A, Ugalde I, Febres-Aldana CA, Tuda C. Fluoroquinolone resistant tuberculosis: A case report and literature review. Respiratory Medicine Case Reports, (2019);27:100829.
- Phyu AN, Aung ST, Palittapongarnpim P, Htet KK, Mahasirimongkol S, Aung HL, Chaiprasert A, Chongsuvivatwong V. Distribution of Mycobacterium tuberculosis Lineages and Drug Resistance in Upper Myanmar. Tropical Medicine and Infectious Disease, (2022); 7(12):448.
- Gabrielsson J, Meibohm B, Weiner D. Pattern recognition in pharmacokinetic data analysis. Journal of the American Association of Pharmaceutical Scientists (2016); 18:47-63.
- Cervantes J, Yokobori N, Hong BY. Genetic identification and drug-resistance characterization of Mycobacterium tuberculosis using a portable sequencing device. A pilot study. Antibiotics, (2020); 9(9):548.
- Hunt M, Mather AE, Sánchez-Busó L, Page AJ, Parkhill J, et al. ARIBA: rapid antimicrobial resistance genotyping directly from sequencing reads. Microbial genomics, (2017); 3(10): e000131.
- Huang Y, Niu B, Gao Y, Fu L, Li W. CD-HIT Suite: a web server for clustering and comparing biological sequences. Bioinformatics, (2010); 26(5):680-2.
- Langmead B, Salzberg SL. Fast gapped-read alignment with Bowtie 2. Nature methods, (2012); 9(4):357-9.
- Li H, Handsaker B, Wysoker A, Fennell T, Ruan J, Homer N, Marth G, Abecasis G, Durbin R, 1000 Genome Project Data Processing Subgroup. The sequence alignment/map format and SAMtools. Bioinformatics, (2009); 25(16):2078-9.
- John GS, Brot N, Ruan J, Erdjument-Bromage H, Tempst P, Weissbach H, Nathan C. Peptide methionine sulfoxide reductase from Escherichia coli and Mycobacterium tuberculosis protects bacteria against oxidative damage from reactive nitrogen intermediates. Proceedings of the National Academy of Sciences, (2001); 98(17):9901-6.
- Lee WL, Gold B, Darby C, Brot N, Jiang X, De Carvalho LP, Wellner D, St. John G, Jacobs Jr WR, Nathan C. Mycobacterium tuberculosis expresses methionine sulphoxide reductases A and B that protect from killing by nitrite and hypochlorite. Molecular microbiology, (2009); 71(3):583-93.
- Hiasa H. The Glu-84 of the ParC subunit plays critical roles in both topoisomerase IV− Quinolone and Topoisomerase IV− DNA interactions. Biochemistry, (2002); 41(39):11779-85.
- Sheng D, Chen X, Li Y, Wang J, Zhuo L, Li Y. ParC, a new partitioning protein, is necessary for the active form of ParA From Myxococcus pMF1 plasmid. Frontiers in Microbiology, (2021); 11(1):623699.
- McCann JR, McDonough JA, Pavelka MS, Braunstein M. β-Lactamase can function as a reporter of bacterial protein export during Mycobacterium tuberculosis infection of host cells. Microbiology, (2007); 153(10):3350–9.
- Gurvitz A, Hiltunen JK, Kastaniotis AJ. Function of heterologous Mycobacterium tuberculosis InhA, a type 2 fatty acid synthase enzyme involved in extending C20 fatty acids to C60-to-C90 mycolic acids, during de novo lipoic acid synthesis in Saccharomyces cerevisiae. Applied and Environmental Microbiology, (2008); 74(16):5078-85
- Schlothauer T, Mogk A, Dougan DA, Bukau B, Turgay K. MecA, an adaptor protein necessary for ClpC chaperone activity. Proceedings of the National Academy of Sciences, (2003); 100(5):2306-11.
- Lee YH, Nam KH, Helmann JD. A mutation of the RNA polymerase β′ subunit (rpoC) confers cephalosporin resistance in Bacillus subtilis. Antimicrobial agents and chemotherapy, (2013); 57(1):56-65.
- García-Castellanos R, Mallorquí-Fernández G, Marrero A, Potempa J, Coll M, Gomis-Ruth FX. On the transcriptional regulation of methicillin resistance: MecI repressor in complex with its operator. Journal of Biological Chemistry, (2004); 279(17):17888-96.
- Blokpoel MC, Murphy HN, O'Toole R, Wiles S, Runn ES, Stewart GR, Young DB, Robertson BD. Tetracycline-inducible gene regulation in mycobacteria. Nucleic acids research, (2005); 33(2): e22-.
- He W, Jiang K, Qiu H, Liao L, Wang S. 16-membered ring macrolides and erythromycin induce ermB expression by different mechanisms. BMC microbiology, (2022); 22(1):152.
- Mérens A, Matrat S, Aubry A, Lascols C, Jarlier V, Soussy CJ, Cavallo JD, Cambau E. The pentapeptide repeat proteins MfpAMt and QnrB4 exhibit opposite effects on DNA gyrase catalytic reactions and on the ternary gyrase-DNA-quinolone complex. Journal of bacteriology, (2009); 191(5):1587-94.
- Singh RS, Chauhan K, Kennedy JF. A panorama of bacterial inulinases: production, purification, characterization and industrial applications. International journal of biological macromolecules, (2017); 96(1):312-322.
- Stern AL, Van der Verren SE, Näsvall J, Gutiérrez-de-Terán H, Selmer M. Structural mechanism of AadA, a dual-specificity aminoglycoside adenylyltransferase from Salmonella enterica. Journal of Biological Chemistry, (2018); 293(29):11481-90.
- Gu S, Rumpel S, Zhou J, Strotmeier J, Bigalke H, Perry K, Shoemaker CB, Rummel A, Jin R. Botulinum neurotoxin is shielded by NTNHA in an interlocked complex. Science, (2012); 335(6071):977-81.
- Farrell DJ, Castanheira M, Chopra I. Characterization of global patterns and the genetics of fusidic acid resistance. Clinical infectious diseases, (2011); 52(suppl_7):S487-92.
- Wassenaar T, Ussery D, Nielsen L, Ingmer H. Review and phylogenetic analysis of qac genes that reduce susceptibility to quaternary ammonium compounds in Staphylococcus species. European Journal of Microbiology and Immunology, (2015); 5(1):44-61.
- Zhu J, He M, Xu W, Li Y, Huang R, Wu S, Niu H. Development of TEM-1 β-lactamase based protein translocation assay for identification of Anaplasma phagocytophilum type IV secretion system effector proteins. Scientific Reports, (2019); 9(1):4235.
- Hunter JH, Gujjar R, Pang CK, Rathod PK. Kinetics and ligand-binding preferences of Mycobacterium tuberculosis thymidylate synthases, ThyA and ThyX. PloS one, (2008); 3(5):e2237.
- Sharma K, Gupta M, Krupa A, Srinivasan N, Singh Y. EmbR, a regulatory protein with ATPase activity, is a substrate of multiple serine/threonine kinases and phosphatase in Mycobacterium tuberculosis. The FEBS journal, (2006); 273(12): 2711-21.
- Sharma K, Gupta M, Pathak M, Gupta N, Koul A, Sarangi S, Baweja R, Singh Y. Transcriptional control of the mycobacterial embCAB operon by PknH through a regulatory protein, EmbR, in vivo. Journal of bacteriology, (2006); 188(8):2936-44.
- Morse JC, Girodat D, Burnett BJ, Holm M, Altman RB, Sanbonmatsu KY, Wieden HJ, Blanchard SC. Elongation factor-Tu can repetitively engage aminoacyl-tRNA within the ribosome during the proofreading stage of tRNA selection. Proceedings of the National Academy of Sciences, (2020); 117(7):3610-20.
- Escuyer VE, Lety MA, Torrelles JB, Khoo KH, Tang JB, Rithner CD, Frehel C, McNeil MR, Brennan PJ, Chatterjee D. The role of the embA and embB gene products in the biosynthesis of the terminal hexaarabinofuranosyl motif of Mycobacterium smegmatisarabinogalactan. Journal of Biological Chemistry, (2001); 276(52):48854-62.
- Berg S, Kaur D, Jackson M, Brennan PJ. The glycosyltransferases of Mycobacterium tuberculosis—roles in the synthesis of arabinogalactan, lipoarabinomannan, and other glycoconjugates. Glycobiology, (2007); 17(6):35R-56R.
- Chakraborty S, Gruber T, Barry III CE, Boshoff HI, Rhee KY. Para-aminosalicylic acid acts as an alternative substrate of folate metabolism in Mycobacterium tuberculosis. Science, (2013); 339(6115):88-91.
- Kremer L, Dover LG, Carrère S, Nampoothiri KM, Lesjean S, Brown AK, Brennan PJ, Minnikin DE, Locht C, Besra GS. Mycolic acid biosynthesis and enzymic characterization of the β-ketoacyl-ACP synthase A-condensing enzyme from Mycobacterium tuberculosis. Biochemical Journal, (2002); 364(2):423-30.
- Aínsa JA, Pérez E, Pelicic V, Berthet FX, Gicquel B, Martín C. Aminoglycoside 2′‐N‐acetyltransferase genes are universally present in mycobacteria: characterization of the aac (2′)‐Ic gene from Mycobacterium tuberculosis and the aac (2′)‐Id gene from Mycobacterium smegmatis. Molecular microbiology, (1997); 24(2):431-41.
- Newton GL, Koledin T, Gorovitz B, Rawat M, Fahey RC, Av-Gay Y. The glycosyltransferase gene encoding the enzyme catalyzing the first step of mycothiol biosynthesis (mshA). Journal of Bacteriology, (2003); 185(11):3476-9.
- Colangeli R, Helb D, Sridharan S, Sun J, Varma‐Basil M, Hazbón MH, Harbacheuski R, Megjugorac NJ, Jacobs Jr WR, Holzenburg A, Sacchettini JC. The Mycobacterium tuberculosis iniA gene is essential for activity of an efflux pump that confers drug tolerance to both isoniazid and ethambutol. Molecular microbiology, (2005); 55(6):1829-40.
- Sørensen MA, Fricke J, Pedersen S. Ribosomal protein S1 is required for translation of most, if not all, natural mRNAs in Escherichia coli in vivo. Journal of molecular biology, (1998); 280(4): 561-9.
- Aubry A, Pan XS, Fisher LM, Jarlier V, Cambau E. Mycobacterium tuberculosis DNA gyrase: interaction with quinolones and correlation with antimycobacterial drug activity. Antimicrobial agents and chemotherapy, (2004); 48(4):1281-8.
- Mikheil DM, Shippy DC, Eakley NM, Okwumabua OE, Fadl AA. Deletion of gene encoding methyltransferase (gidB) confers high-level antimicrobial resistance in Salmonella. The Journal of antibiotics, (2012); 65(4):185-92.
- Zeng S, Soetaert K, Ravon F, Vandeput M, Bald D, Kauffmann JM, Mathys V, Wattiez R, Fontaine V. Isoniazid bactericidal activity involves electron transport chain perturbation. Antimicrobial agents and chemotherapy, (2019); 63(3):10-128.
- hang H, Deng JY, Bi LJ, Zhou YF, Zhang ZP, Zhang CG, Zhang Y, Zhang XE. Characterization of Mycobacterium tuberculosis nicotinamidase/pyrazinamidase. The FEBS journal, (2008); 275(4):753-62..
- Fraaije MW, Kamerbeek NM, Heidekamp AJ, Fortin R, Janssen DB. The prodrug activator EtaA from Mycobacterium tuberculosis is a Baeyer-Villiger monooxygenase. Journal of Biological Chemistry, (2004); 279(5):3354-60.
- Jia H, Chu H, Dai G, Cao T, Sun Z. Rv1258c acts as a drug efflux pump and growth controlling factor in Mycobacterium tuberculosis. Tuberculosis, (2022); 133:102172.
- Dzyubak E, Yap MN. The expression of antibiotic resistance methyltransferase correlates with mRNA stability independently of ribosome stalling. Antimicrobial agents and chemotherapy, (2016); 60(12):7178-88.
- Kumar S, Parvathi A, Hernandez RL, Cadle KM, Varela MF. Identification of a novel UDP-N-acetylglucosamine enolpyruvyl transferase (MurA) from Vibrio fischeri that confers high fosfomycin resistance in Escherichia coli. Archives of microbiology, (2009); 191:425-9.
- Alland D, Kramnik I, Weisbrod TR, Otsubo L, Cerny R, Miller LP, Jacobs Jr WR, Bloom BR. Identification of differentially expressed mRNA in prokaryotic organisms by customized amplification libraries (DECAL): the effect of isoniazid on gene expression in Mycobacterium tuberculosis. Proceedings of the National Academy of Sciences, (1998); 95(22):13227-32.
- Fassbinder F, Kist M, Bereswill S. Structural and functional analysis of the riboflavin synthesis genes encoding GTP cyclohydrolase II (ribA), DHBP synthase (ribBA), riboflavin synthase (ribC), and riboflavin deaminase/reductase (ribD) from Helicobacter pylori strain P1. FEMS microbiology letters, (2000); 191(2):191-7.
- Madhusudan K, Nagaraja V. Mycobacterium smegmatis DNA gyrase: cloning and overexpression in Escherichia coli. Microbiology, (1995); 141(12):3029-37.
- Rahman MA, Sobia P, Dwivedi VP, Bhawsar A, Singh DK, Sharma P, Moodley P, Van Kaer L, Bishai WR, Das G. Mycobacterium tuberculosis TlyA protein negatively regulates T helper (Th) 1 and Th17 differentiation and promotes tuberculosis pathogenesis. Journal of Biological Chemistry, (2015); 290(23):14407-17.
- Doran JL, Pang Y, Mdluli KE, Moran AJ, Victor TC, Stokes RW, Mahenthiralingam E, Kreiswirth BN, Butt JL, Baron GS, Treit JD. Mycobacterium tuberculosis efpA encodes an efflux protein of the QacA transporter family. Clinical Diagnostic Laboratory Immunology, (1997); 4(1):23-32.
- Holberger LE, Hayes CS. Ribosomal protein S12 and aminoglycoside antibiotics modulate A-site mRNA cleavage and transfer-messenger RNA activity in Escherichia coli. Journal of Biological Chemistry, (2009); 284(46):32188-200.
- Singh R, Wiseman B, Deemagarn T, Jha V, Switala J, Loewen PC. Comparative study of catalase-peroxidases (KatGs). Archives of biochemistry and biophysics, (2008); 471(2):207-14.
- Zhou YN, Lubkowska L, Hui M, Chen S, Strathern J, Jin DJ, Kashlev M. Isolation and characterization of RNA polymerase rpoB mutations that alter transcription slippage during elongation in Escherichia coli. Journal of Biological Chemistry, (2013); 288(4):2700-10.
- Flores AR, Parsons LM, Pavelka Jr MS. Genetic analysis of the β-lactamases of Mycobacterium tuberculosis and Mycobacterium smegmatis and susceptibility to β-lactam antibiotics. Microbiology, (2005); 151(2):521-32.
- Rifat M, Milton AH, Hall J, Oldmeadow C, Islam MA, Husain A, Akhanda MW, Siddiquea BN. Development of multidrug resistant tuberculosis in Bangladesh: a case-control study on risk factors. PloS one, (2014); 9(8):e105214.
- Lomtadze N, Aspindzelashvili R, Janjgava M, Mirtskhulava V, Wright A, Blumberg HM, Salakaia A. Prevalence and risk factors for multidrug-resistant tuberculosis in the Republic of Georgia: a population-based study. The International journal of tuberculosis and lung disease, (2009); 13(1):68-73.
- McQuaid CF, Horton KC, Dean AS, Knight GM, White RG. The risk of multidrug-or rifampicin-resistance in males versus females with tuberculosis. European Respiratory Journal, (2020); 56(3): 2000626.
- Mulu W, Mekkonnen D, Yimer M, Admassu A, Abera B. Risk factors for multidrug resistant tuberculosis patients in Amhara National Regional State. African health sciences, (2015); 15(2):368-77.
- De Martino M, Lodi L, Galli L, Chiappini E. Immune response to Mycobacterium tuberculosis: a narrative review. Frontiers in pediatric, (2019); 7: 350.
- Mehari K, Asmelash T, Hailekiros H, Wubayehu T, Godefay H, Araya T, Saravanan M. Prevalence and factors associated with multidrug-resistant tuberculosis (MDR-TB) among presumptive MDR-TB patients in Tigray Region, Northern Ethiopia. Canadian Journal of Infectious Diseases and Medical Microbiology, (2019); 2019(1):1-8.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0