

Full Length Research Article
Whole genome sequence of Lactiplantibacillus plantarum strain HA9 isolated from conventional Iraqi cheese: First report
Hasanain Qasim Mezaal Al-kaabi*, Rahman Laibi Chelab
Adv. life sci., vol. 11, no. 2, pp. 438-443, May 2024
*– Corresponding Author: Hasanain Qasim Mezaal Al-kaabi (hasaninqasim00.bio@utq.edu.iq )
Authors' Affiliations
[Date Received: 14/09/2023; Date Revised: 15/03/2024; Date Available Online: 18/04/2024]
Editorial Note on Version of Record
31 May 2025: This article has been corrected. See https://doi.org/10.62940/als.v13i0.4275 for more information.
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Lactiplantibacillus plantarum is a renowned probiotic known for its ability to enhance health when ingested in sufficient quantities. While L. plantarum typically exhibits unique characteristics, variations in strains and environments can occur. Therefore, it is crucial to conduct genetic identification of the strain before incorporating it into products or administering it for health benefits.
Methods: After being isolated from classic artisanal milk cheeses, the bacterial DNA extraction kit from GENEAID, Korea, was used, and strain HA9’s genomic DNA was isolated. Then, the highly conserved section of ribosomal RNA was amplified with universal primers. The results were confirmed by a blast analysis of the NCBI database. The genome of HA9 was sequenced using the Illumina and PacBio Sequel II technologies (HN00194138; Psomagen/USA). The Prokaryotic Dynamic Programming Gene-Finding Algorithm software was applied to forecast coding genes. The Pathosystems Resource Integration Center constructed the L. plantarum genome.
Result: According to the findings, the genome of the L. plantarum HA9 strain has a length of 3,321,986 bp and a GC percentage of 44.40. This assembled genome contained a total of 1,420 coding proteins and 82 contigs, as discovered by the Comprehensive Genome Analysis. The isolate had been submitted to the NCBI and received the HA9 genome sequence (accession ID: JAUTDL000000000.1).
Conclusion Microbes’ whole-genome sequencing plays a crucial role in clarifying their probiotic properties and enables accurate identification down to the strain level. The authors argue that further research should be done on probiotics’ whole genomes, with a special emphasis on second-generation strains.
Keywords: Lactiplantibacillus plantarum; Probiotics; Whole genome sequencing
Introduction![]()
Lactiplantibacillus plantarum, previously known as Lactobacillus plantarum, is a type of lactic acid bacterium commonly encountered in various fermented foods, such as artisanal milk cheeses. L. plantarum is one of the predominant microbial species engaged in fermentation, which is a procedure used historically to create artisanal milk cheeses [1].
Depending on the type of milk used, the region, and the particular cheese-making methods utilized by the artisanal cheese manufacturers, isolates of L. plantarum from traditional artisanal milk cheeses might differ. Different L. plantarum strains or isolates may have unique traits, such as the capacity to create enzymes. L. plantarum strains isolated from various cheeses had various proteolytic and lipolytic capabilities as well as various levels of resistance to stress [2]. Some strains of L. plantarum are regarded as a probiotic, which means that when taken in sufficient levels, it gives health advantages. It is thought to strengthen immune system function, increase gut microbial balance, and support digestive health [3]. Low pH values, high salt concentrations, and a range of temperatures are only a few environmental variables to which L. plantarum is very tolerant. Due to its versatility, it may live and flourish in many gut and food conditions. For instance, according to [4], L. plantarum can withstand pH levels as low as 2.51, bile concentrations as high as 0.3%, pancreatin concentrations as high as 0.5%, NaCl concentrations as high as 5%, and temperatures as low as 4°C to 45°C [5]. Due to these qualities, L. plantarum is able to colonize a variety of fermented foods as well as resist the challenging circumstances of the gastrointestinal system. In response to several environmental cues including pH, bile, and oxygen, L. plantarum may also alter the expression of its genes. Due to its adaptability, L. plantarum is able to retain its functioning and viability in a variety of conditions [6]. The L. plantarum creates bacteriocins, which are antimicrobial peptides that can stop the growth of other hazardous bacteria. The capacity of these bacteriocins to outcompete harmful bacteria and aid in food preservation makes it especially beneficial [7]. L. plantarum has a flexible metabolic route that enables it to ferment several types of carbohydrates and generate a range of byproducts, such as carbon dioxide, ethanol, and other organic acids [8]. The L. plantarum is capable of forming biofilms. According to [9], the creation of biofilms can be advantageous in some situations, such as boosting adherence to probiotic colonization in the gut or enhancing during fermentation processes. L. plantarum has the capacity to produce a number of vitamins, including several B-group vitamins, which may improve the nutritional content of food overall and be advantageous to the host [10]. Despite the fact that L. plantarum often demonstrates these traits, there may be variances between different strains and depending on the particular circumstances it faces. As a result, the L. plantarum genome may exhibit structural variants (SVs), particularly in genes involved in metabolism, stress tolerance, biofilm formation, and virulence [11]. The phenotypic traits of L. plantarum strains, such as their glucose metabolism, ethanol tolerance, anti-fungal activity, and interactions with other microorganisms, can also be affected by these genetic differences [12]. For instance, the L. plantarum B2 strain has a mutation in the FMN-aptamer of the riboswitch, which regulates the expression of the rib operon. The strain benefits from the increased production and accumulation of riboflavin and FMN as sources of vitamin B2, but this mutation also decreases the riboswitch’s regulatory activity and makes the strain more susceptible to roseoflavin, a riboflavin analog that prevents bacterial growth [13]. Thus, it is essential to establish the genetic identity of the strain both before and throughout manufacturing.
The antimicrobial susceptibility profile of the strain and its ability to spread AMR genes or virulence factors to other bacteria are important criteria in establishing a probiotic’s safety for use in people and/or animals. Any bacterium employed as a probiotic must be demonstrated to be devoid of AMR and virulence factors, both of which can be transmitted by ingestion, and all probiotics should have their antimicrobial susceptibility profiles assessed [14,15]. If a probiotic is shown to be the cause or is suspected of being the source of any invasive illness, knowledge of AMR patterns can help clinicians make treatment options. Additionally, understanding a microbe’s AMR phenotype can aid in the development of innovative multi-component treatment regimens, which is crucial in light of the growing body of research supporting the simultaneous administration of probiotics and antibiotics [16-18].
The goals of this study were to determine the genome sequence of L. plantarum isolated from classic artisanal milk cheese and to confirm their taxonomic, strain-level identities and more details.
Methods![]()
L. plantarum HA9 culture conditions
Lactic acid bacteria (LAB) were isolated from classic artisanal milk cheeses. Isolation was done using, with minor modifications, a previously published approach [19]. In short, we plated each sample on MRS agar after diluting and homogenizing 1 gram of each sample in MRS broth. Then incubated the plates anaerobically at 37 °C for 48 hours and selected colonies with different morphologies. The colonies were purified by sub-culturing and confirmed them as Lactobacillus spp. based on their catalase-negative, gram-positive, rod-shaped appearance. The bacterial isolates stored in MRS broth with 20% glycerol.
Extraction of genomic DNA from L. plantarum HA9
Using the bacterial DNA extraction kit from GENEAID, Korea, genomic DNA was isolated from strain HA9 [20]. Thermo Fisher Scientific’s NanoDrop 2000 spectrophotometer and Life Technologies’ Qubit 3.0 were used for DNA quantification and concentration measurements, respectively, and 0.8% (w/v) agarose gel electrophoresis was used to assess the purity of the DNA. Then, the highly conserved section of ribosomal RNA was amplified with universal primers 27F and 1492R, the PCR product was sequenced with these primers [21], and the results were confirmed by a blast analysis of the NCBI database [22].
Whole-genome sequence analysis of L. plantarum HA9
The genome of L. plantarum HA9 was sequenced using the Illumina and PacBio Sequel II technologies (HN00194138; Psomagen/USA). After passing quality control (QC), the DNA from the samples was randomly fragmented and ligated with 5′ and 3′ adapters to produce a library. The generated NGS library will then be sequenced. Following the completion of the sequencing, the raw data was analyzed for parameters such as GC (percentage), total bases, and total reads. To reduce analytical biases, quick QC (https://www.bioinformatics.babraham.ac.uk/projects/fastqc/) and quality filtering were utilized. We used the phred quality score (Q20(%) and Q30(%) to assess data quality at the end of each cycle [23].
Genome annotation of L. plantarum HA9
Coding genes were predicted using Prodigal (Prokaryotic Dynamic Programming Gene-finding Algorithm) software [24] Filtering for proteins with the highest sequence similarity using annotations with e 1e-5, the Diamond software [25] was employed to contrast expected coding sequences in order to provide functional annotation information. The HMMER software [26] was utilized for comparing these sequences with protein group models in order to identify the families that scored the highest. The predicted code sequence regions were compared against the KEGG (Kyoto Encyclopedia of Genes and Genomes), COG (Cluster of Orthologous Groups of proteins), GO (Gene Ontology), CAZy (Carbohydrate-Active Enzymes Database), and NR (non-redundant) databases in order to complete the foundational functional prediction and annotation. Using the Circus (v0.69) program [27] many data sets were shown in one single genomic circular map.
Genome analysis
PATRICS, a sophisticated genome analysis service, constructed L. plantarum genome [28]. The L. plantarum reference genome mapped sequencing reads. Functional genes in subsystem categories were annotated in the genome, mapping results were compared, conserved and unique sequencing properties were identified, and high-quality maps were created to verify and display the described features. Bioinformatics techniques removed duplicates, found variants, and mapped filtered data reads to the reference genome [29-31].
Results![]()
The quality of the HA9 DNA
To guarantee the concentration and quality of DNA, the bioinformatics tools provided by PATRIC were employed. The genome isolates of strain HA9 displayed a coarse consistency score of 97.8, was complemented by a fine consistency score of 96.1, and achieved a checkM completeness rating of 100. As a result, this genome exemplified exceptional overall quality.
Genomic characteristics of HA9
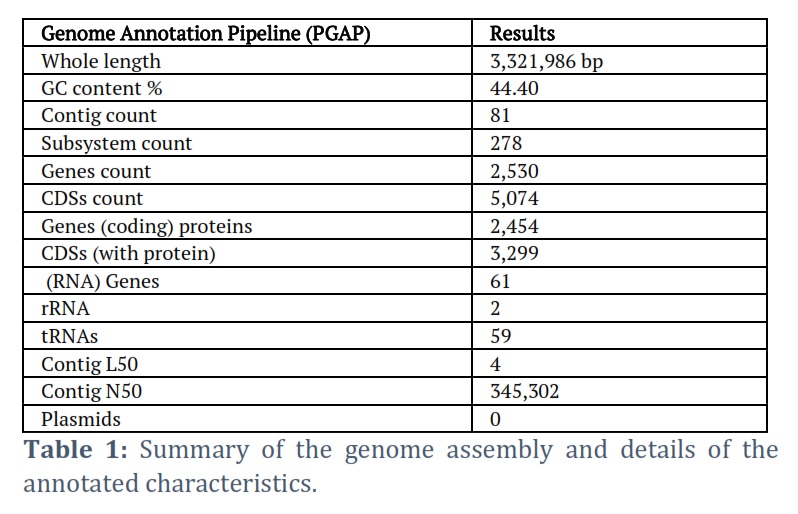
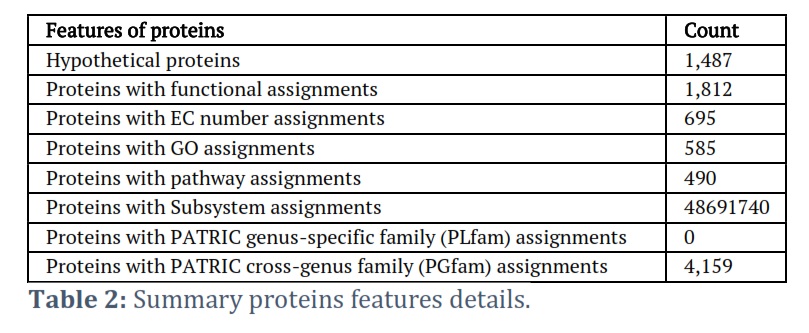
The cutting-edge PacBio Sequel II sequencing technology was employed to unveil the entire genome sequence of L. plantarum strain HA9. This breakthrough resulted in NCBI receiving the HA9 genome sequence (accession ID: JAUTDL000000000.1). Impressively, the HA9 genome spans 3,321,986 base pairs, consisting of a singular circular chromosome with a GC content of 44.40%, and notably, no plasmids were detected. Table 1 encapsulates the anticipated 3,138 genes, along with 2 rRNAs (5S, 16S, and 23S) and 59 tRNAs present in this comprehensive genome. Meanwhile, the strain HA9 whole-genome circle map is shown in Figure 1, meticulously delineating key genomic features such as homologous genes, GC content, genomic islands, gene distribution across forward and antisense strands, and COG functional classification of genes, and more intricate details. Noteworthy is the genome’s high quality, substantiated by annotation statistics and comparison with PATRIC genomes of the same species. Following meticulous read filtering, the phred quality scores of bases exceeded Q20 and Q30 at an impressive rate of 97.8% and 94.0%, respectively. Furthermore, Comprehensive Genome Analysis unveiled 81 contigs spanning 3,314,413 base pairs, housing 1,420 coding proteins in this meticulously assembled genome, as detailed in Table 2. The genome’s GC content averages 44.3%, with Figure 1 providing a schematic representation of GC content and GC skew analysis. Each protein subsystem within the genome contributes to specific biological activities or structural complexes, with a striking 278 genome-specific subsystems found across annotation. Figure 2 offers a comprehensive overview of the genome’s subsystem organization, highlighting its intricate biological functionality and structural complexity.
Phylogenetic Analysis
For phylogenetic research, PATRIC offers representative and reference genomes. Based on 16S rRNA sequences, Figure 3 shows a neighbor-joining phylogenetic tree; Figure 4 shows the nearest reference and typical genotypes found during the research.
Figures & Tables

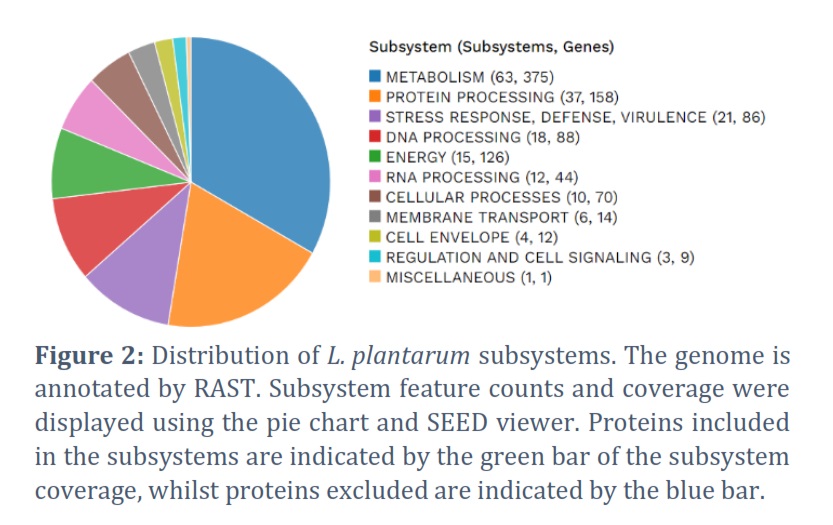
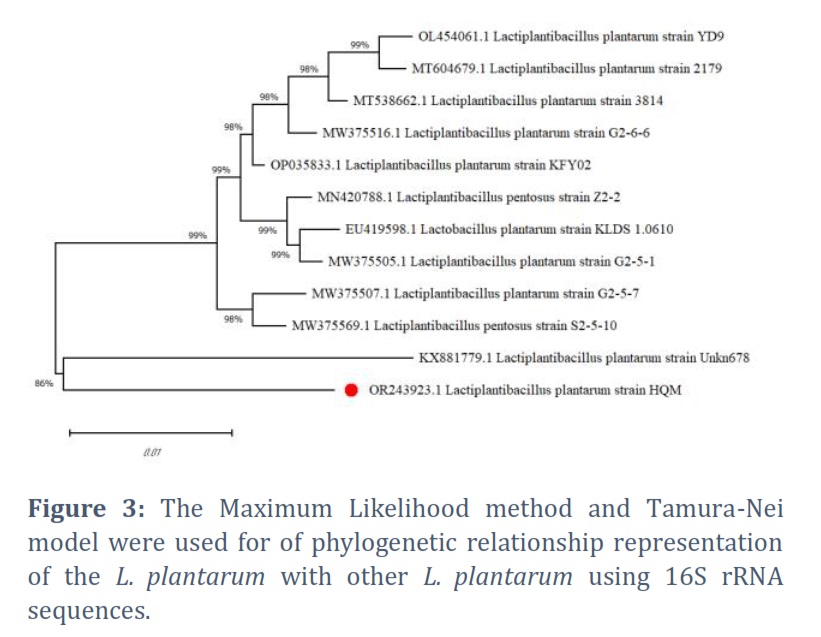
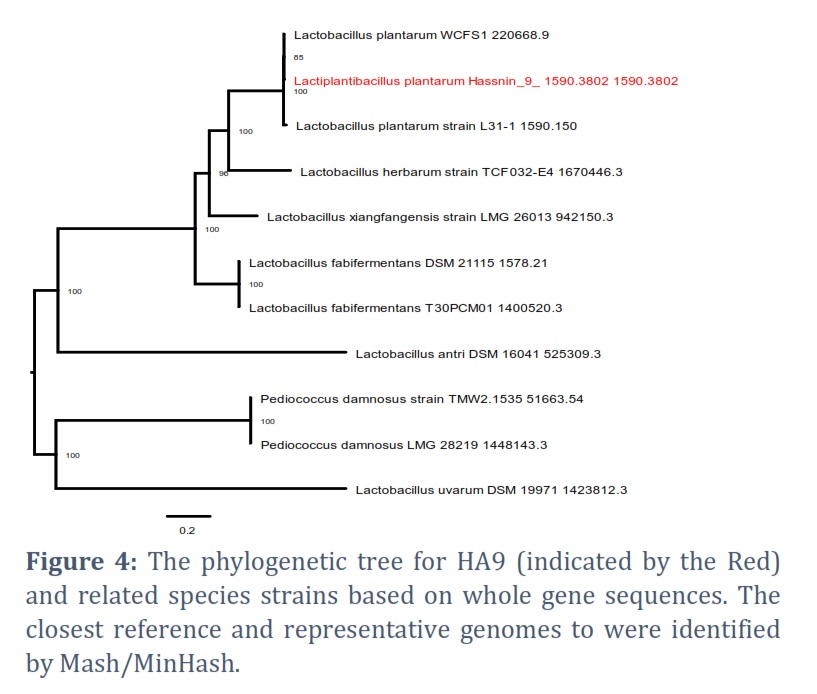
16S rRNA analysis was used to identify the HA9 strain and classify it as L. plantarum. This taxonomy corresponds to the species commonly present in sauerkraut [32].
The size of the HA9 genome and the proportion of its GC content were identical to those previously reported for L. plantarum strains. Within the lactobacilli group, the genome of the L. plantarum species is among the largest genomes overall, ranging in size from 2.91 to 3.7 Mb. The whole genome of L. plantarum has a median length of 3.27272 Mb, a median protein counts of 2997, and a GC content of 44.5%, according to the NCBI database. It is considered that the large genomes of L. plantarum are directly related to the environmental flexibility and extraordinary adaptability it possesses to a wide variety of settings [33].
The HA9 strain of L. plantarum occupies a distinct position on the evolutionary tree when compared to other strains of the same species. Its closest counterparts include strains such as L. plantarum WCFS1, which is among the most extensively researched lactobacilli. Notably, the genome of L. plantarum WCFS1 was unveiled many years ago. This particular strain of L. plantarum demonstrates remarkable resilience and a relatively high survival rate when passing through the digestive system, and it can be cultivated to high densities. Additionally, it is easily modifiable genetically [34]. Furthermore, the genome of L. plantarum WCFS1 has not been found to include any transferable antibiotic resistance genes [35]. To understand the distinctions between HA9 and WCFS1 better, further comprehensive comparative genomic investigations are required.
A viable technique to enrich fermented meals and boost the health-promoting impact of probiotics is to make use of starter cultures that produce vitamins or probiotics [36]. On the other hand, future investigations both in silico and in vitro should shed light on the manner in which several biochemical biosynthesis processes are linked to the genetic ability to biosynthesize vitamins. We also evaluated genetic data linked to safety evaluation, like genes that encode toxins and virulence factors, however we were unable to find any risk-associated sequences in the genome of L. plantarum HA9.
Acknowledgments
Authors acknowledge Medical and science colleges/Misan University and Science college for their support.
Conflict of Interest
The authors declare that there is no conflict of interest.
Al-kaabi H. Q. and Chelab R. L. designed the study and performed the experiments. In addition, both authors analyzed the data and wrote the manuscript.
![]() References
References
- Milioni C, Martínez B, Degl’Innocenti S, Turchi B, Fratini F, et al. A novel bacteriocin produced by Lactobacillus plantarum LpU4 as a valuable candidate for biopreservation in artisanal raw milk cheese. Dairy science & technology, (2015); 95: 479-494.
- Surve S, Shinde DB, Kulkarni R. Isolation, characterization and comparative genomics of potentially probiotic Lactiplantibacillus plantarum strains from Indian foods. Scientific Reports, (2022); 12(1): 1940.
- Hsu H-Y, Liao Y-C, Lin S-H, Lin J-S, Lee C-C, et al. Safety assessment of Lactiplantibacillus plantarum TWK10 based on whole-genome sequencing, phenotypic, and oral toxicity analysis. Microorganisms, (2022); 10(4): 784.
- Fidanza M, Panigrahi P, Kollmann TR. Lactiplantibacillus plantarum–nomad and ideal probiotic. Frontiers in microbiology, (2021); 6(12): 712236.
- Nath S, Sikidar J, Roy M, Deb B. In vitro screening of probiotic properties of Lactobacillus plantarum isolated from fermented milk product. Food Quality and Safety, (2020); 4(4): 213-223.
- Garcia-Gonzalez N, Battista N, Prete R, Corsetti A. Health-promoting role of Lactiplantibacillus plantarum isolated from fermented foods. Microorganisms, (2021); 9(2): 349.
- Flórez AB, Mayo B. Genome analysis of Lactobacillus plantarum LL441 and genetic characterisation of the locus for the lantibiotic plantaricin C. Frontiers in Microbiology, (2018); 91916.
- Wieczorek MN, Drabińska N. Flavour generation during lactic acid fermentation of brassica vegetables—literature review. Applied Sciences, (2022); 12(11): 5598.
- García-Reyes RA, García-Cancino A, Arrevillaga-Boni G, Espinoza-Monje M, Gutiérrez-Zamorano C, et al. Identification and Characterization of Probiotic Lactiplantibacillus plantarum BI-59.1 Isolated from tejuino and Its Capacity to Produce Biofilms. Current Microbiology, (2023); 80(7): 220.
- Echegaray N, Yilmaz B, Sharma H, Kumar M, Pateiro M, et al. A novel approach to Lactiplantibacillus plantarum: From probiotic properties to the omics insights. Microbiological Research, (2022); 127289.
- Di Cerbo A, Palmieri B, Aponte M, Morales-Medina JC, Iannitti T. Mechanisms and therapeutic effectiveness of lactobacilli. Journal of clinical pathology, (2016); 69(3): 187-203.
- Yu AO, Goldman EA, Brooks JT, Golomb BL, Yim IS, et al. Strain diversity of plant‐associated Lactiplantibacillus plantarum. Microbial Biotechnology, (2021); 14(5): 1990-2008.
- Ripa I, Ruiz-Masó JÁ, De Simone N, Russo P, Spano G, et al. High vitamin B2 production by the Lactiplantibacillus plantarum B2 strain carrying a mutation in the aptamer P1 helix of the FMN-riboswitch that regulates expression of the bacterial rib operon. BioRxiv, (2021); 2021.2005. 2020.444923.
- Gheziel C, Russo P, Arena MP, Spano G, Ouzari H-I, et al. Evaluating the probiotic potential of Lactobacillus plantarum strains from Algerian infant feces: towards the design of probiotic starter cultures tailored for developing countries. Probiotics and antimicrobial proteins, (2019); 11113-123.
- Sakurai K, Kawasaki H. Genetic variation during long-term preservation of bacteria in public culture collections. International Journal of Systematic and Evolutionary Microbiology, (2018); 68(5): 1815-1821.
- McFarland LV, Evans CT, Goldstein EJ. Strain-specificity and disease-specificity of probiotic efficacy: a systematic review and meta-analysis. Frontiers in medicine, (2018); 7(5)124.
- Sansotta N, Peroni DG, Romano S, Rugiano A, Vuilleumier P, et al. The good bugs: the use of probiotics in pediatrics. Current Opinion in Pediatrics, (2019); 31(5): 661-669.
- Yelin I, Flett KB, Merakou C, Mehrotra P, Stam J, et al. Genomic and epidemiological evidence of bacterial transmission from probiotic capsule to blood in ICU patients. Nature medicine, (2019); 25(11): 1728-1732.
- Cui X, Shi Y, Gu S, Yan X, Chen H, et al. Antibacterial and antibiofilm activity of lactic acid bacteria isolated from traditional artisanal milk cheese from Northeast China against enteropathogenic bacteria. Probiotics and antimicrobial proteins, (2018); 10(4): 601-610.
- Arseneau JR, Steeves R, Laflamme M. Modified low-salt CTAB extraction of high-quality DNA from contaminant-rich tissues. Mol Ecol Resour, (2017); 17(4): 686-693.
- Frank JA, Reich CI, Sharma S, Weisbaum JS, Wilson BA, et al. Critical evaluation of two primers commonly used for amplification of bacterial 16S rRNA genes. Applied and environmental microbiology, (2008); 74(8): 2461-2470.
- Sayers EW, Beck J, Bolton EE, Bourexis D, Brister JR, et al. Database resources of the national center for biotechnology information. Nucleic acids research, (2021); 49(D1): D10.
- Bankevich A, Nurk S, Antipov D, Gurevich AA, Dvorkin M, et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. Journal of computational biology, (2012); 19(5): 455-477.
- Hyatt D, Chen GL, Locascio PF, Land ML, Larimer FW, et al. Prodigal: prokaryotic gene recognition and translation initiation site identification. BMC Bioinformatics, (2010); 11119.
- Buchfink B, Xie C, Huson DH. Fast and sensitive protein alignment using DIAMOND. Nat Methods, (2015); 12(1): 59-60.
- Eddy SR. A new generation of homology search tools based on probabilistic inference. Genome Inform, (2009); 23(1): 205-211.
- Krzywinski M, Schein J, Birol I, Connors J, Gascoyne R, et al. Circos: an information aesthetic for comparative genomics. Genome Research, (2009); 19(9): 1639-1645.
- Gillespie JJ, Wattam AR, Cammer SA, Gabbard JL, Shukla MP, et al. PATRIC: the comprehensive bacterial bioinformatics resource with a focus on human pathogenic species. Infection and immunity, (2011); 79(11): 4286-4298.
- Brettin T, Davis JJ, Disz T, Edwards RA, Gerdes S, et al. RASTtk: a modular and extensible implementation of the RAST algorithm for building custom annotation pipelines and annotating batches of genomes. Scientific reports, (2015); 5(1): 8365.
- Li H, Durbin R. Fast and accurate long-read alignment with Burrows–Wheeler transform. Bioinformatics, (2010); 26(5): 589-595.
- McKenna A, Hanna M, Banks E, Sivachenko A, Cibulskis K, et al. The Genome Analysis Toolkit: a MapReduce framework for analyzing next-generation DNA sequencing data. Genome research, (2010); 20(9): 1297-1303.
- Zheng J, Wittouck S, Salvetti E, Franz CM, Harris HM, et al. A taxonomic note on the genus Lactobacillus: Description of 23 novel genera, emended description of the genus Lactobacillus Beijerinck 1901, and union of Lactobacillaceae and Leuconostocaceae. International journal of systematic and evolutionary microbiology, (2020); 70(4): 2782-2858.
- Evanovich E, de Souza Mendonça Mattos PJ, Guerreiro JF. Comparative genomic analysis of Lactobacillus plantarum: an overview. International journal of genomics, (2019); 9(4): 452-465.
- Pell LG, Horne RG, Huntley S, Rahman H, Kar S, et al. Antimicrobial susceptibilities and comparative whole genome analysis of two isolates of the probiotic bacterium Lactiplantibacillus plantarum, strain ATCC 202195. Scientific Reports, (2021); 11(1): 15893.
- Kleerebezem M, Boekhorst J, van Kranenburg R, Molenaar D, Kuipers OP, et al. Complete genome sequence of Lactobacillus plantarum WCFS1. Proceedings of the National Academy of Sciences U S A, (2003); 100(4): 1990-1995.
- Sanger F, Air GM, Barrell BG, Brown NL, Coulson AR, et al. Nucleotide sequence of bacteriophage φX174 DNA. nature, (1977); 265(5596): 687-695.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0