

Full Length Research Article
Structure-Based Virtual Screening of Antiviral Compounds Targeting the Norovirus RdRp Protein
Leena S. Alqahtani1, Ahmad Salah Alkathiri2, Abdulrahman Alzahrani3, Rashed Mohammed Alghamdi4, Waad Abdulrahmman Alamri5, Mohammad Azhar Kamal6, Ahmed Hamdan Aloufi7, Ali Saeed Alamri8, Qamre Alam9*
Adv. life sci., vol. 11, no. 2, pp. 488-492, May 2024
*– Corresponding Author: Qamre Alam (alamqa2022@gmail.com)
Authors' Affiliations
2. Department of Health Promotion and Education, Faculty of Public Health & Health Informatics, Umm Al-Qura University, Makkah – Kingdom of Saudi Arabia
3. Department of Applied Medical Sciences, Applied College, Al-Baha University, Al-Baha City – Kingdom of Saudi Arabia
4. Department of Laboratory Medicine, Faculty of Applied College, Al-Baha University – Kingdom of Saudi Arabia
5. Department of Genetic Medicine, Faculty of Medicine, King Abdulaziz University, Jeddah – Kingdom of Saudi Arabia
6. Department of Pharmaceutics, College of Pharmacy, Prince Sattam Bin Abdulaziz University, Al-Kharj – Kingdom of Saudi Arabia
7. Department of Pathology and Laboratory Medicine, Imam Abdulrahman bin Faisal Hospital Ministry of National Guard Health Affairs, Dhahran – Kingdom of Saudi Arabia
8. Molecular Pathology Lab Department of Pathology and Laboratory Medicine, Ministry of National Guard Hospital and Health Affairs (MNGHA) – Kingdom of Saudi Arabia
9. Molecular Genomics and Precision Medicine Department, ExpressMed Diagnostics and Research, Block, Zinj – Kingdom of Saudi Arabia
[Date Received: 23/10/2023; Date Revised: 13/11/2023; Date Available Online: 18/04/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Human noroviruses (NV) are the primary etiological organisms causing acute gastroenteritis around the world, causing severe morbidity and imposing a significant economic burden. The RNA-dependent RNA polymerase (RdRp) is essential for viral replication and could be a promising target for anti-NV therapeutics. Despite the discovery of a few NV RdRp inhibitors, the majority of these pharmaceuticals have demonstrated limited efficacy in inhibiting viral replication in cellular models.
Methods: In this study, computational screening of antiviral compounds was conducted targeting the NV RdRp protein. The assessment was based on binding poses and the key residues of RdRp involved in interactions with compounds.
Results: The compounds namely, Ribavirin, BMS806, Dihydromyricetin, R7935788, and LY2784544 were found to bind the RdRp protein with high affinity. Notably, these compounds displayed significantly lower binding affinities compared to the positive control, PPNDS. In addition, these compounds exhibited many RdRp protein binding residues that were also present in the PPNDS.
Conclusion: The results presented here suggest that these compounds have the potential to be used as inhibitors of NV RdRp in the development of antiviral medications. Nevertheless, due to the computational nature of this study, it is imperative to do experimental validation.
Keywords: Noroviruses; RdRp; Virtual screening; Antiviral Compounds
Introduction![]()
Human noroviruses (NV), which belong to the Caliciviridae family, are the leading cause of acute gastroenteritis globally. They cause major disease and place a significant financial load [1,2]. The management of NV infections is inherently challenging due to their capacity to quickly spread through food and water, their wide genetic variety, and their amazing tolerance to many environmental circumstances [3]. The absence of effective diagnosis, therapy options specific to NV, and preventive measures like immunizations exacerbates the situation [4,5]. In order to tackle this significant problem, it is imperative to create small-molecule treatments and preventive measures that possess anti-NV capabilities, alongside efficient immunization strategies.
According to the World Health Organization, infection prevention and control (IPC) is a systematic and evidence-based approach that aims to reduce the negative impact of infections on patients and healthcare workers. IPC, as a subject of epidemiology, significantly contributes to the social sciences, infectious diseases, and global health by providing crucial measurements and solutions. The effectiveness of infection prevention and control (IPC) interventions is a significant public health concern that contributes to patient safety and the reformation of healthcare systems. The utilization of effective IPC interventions is intricately connected to the prevention of healthcare-associated illnesses, as well as the management of global epidemics and pandemics.
The human norovirus (NV) genome has three open reading frames (ORFs) and has a size of 7.7 kb. ORF1 encodes polyproteins that are subsequently cleaved into non-structural proteins, including viral proteases, RNA-dependent RNA polymerase (RdRp), and VPg-like protein [6]. The RdRp is notable for its critical role in viral replication and promise as a target for anti-NV medicines [7,8]. Although certain NV RdRp inhibitors have been discovered, most of these medications have demonstrated limited efficacy in halting viral propagation within living organisms. This is mostly attributed to their restricted pharmacological properties and incapacity to penetrate cellular membranes [9,10]. Through the use of intensive computational approaches, the purpose of this study was to identify novel RdRp inhibitors that could be used for the treatment of gastroenteritis.
Methods![]()
Retrieval and preparation of RdRp
The crystal structure (3D structure) of RdRp (PDB ID: 4LQ3) was obtained from the Protein Data Bank. This 3D structure was determined by X-ray diffraction method with 2.60 Å [11]. The co-crystal ligand (PPNDS), water molecules, and other heteroatoms were extracted from the monomeric unit of 3D structure. The cleaned 3D structure was subsequently 'prepared' and saved in pdb format using Discovery Studio.
Compound library preparation
In order to find a potential antiviral agent that targets the RdRp of NV, a collection of 752 antiviral compounds that are structurally diverse, medicinally active, and cell permeable were selected for the virtual screening in this study. These compounds have been identified as Targets of HCV Protease, HIV Protease, Integrase, and Reverse Transcriptase, amongst other enzymes; also, this collection contains a number of drugs that have been granted approval by the FDA. All the compounds were downloaded in ‘sdf’ and prepared for docking experiments using Discovery Studio 2021.
Structure-Based Virtual Screening of Antiviral Compounds Targeting the Norovirus RdRp Protein
Computational modeling is important in hit discovery and lead optimization, specifically through the use of structure-based virtual screening (VS) [12]. Docking, the primary computational technique used in VS, has been extensively researched and widely used in drug discovery over the last decade [13]. For this study, the prepared antiviral compound library was screened against the RdRp protein of NV using the PyRx0.8 tool [14]. The grid center coordinates were defined as X = -19.743, Y = -26.041, and Z = -5.212. The co crystal ligand of 4LQ3 (PPNDS) was selected as positive control for this study.
Results![]()
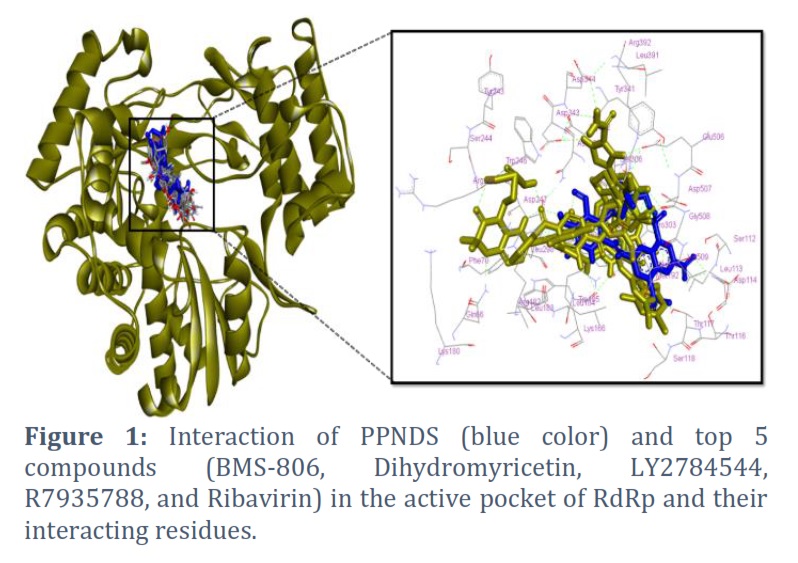
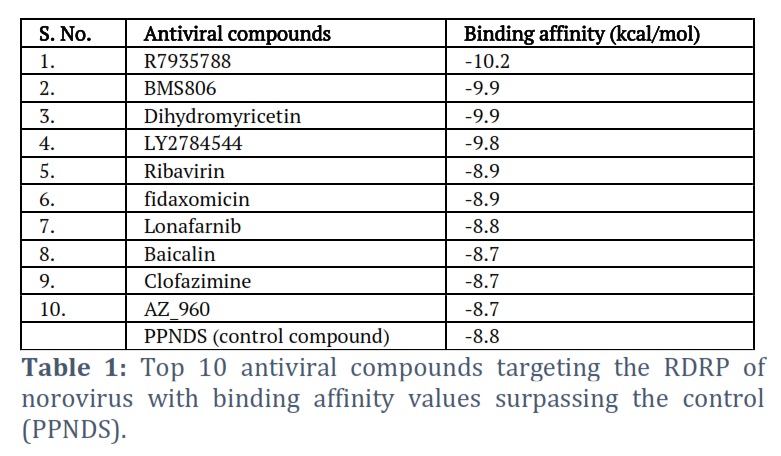
Using computational screening approaches, this study seeks probable inhibitors of the NV RdRp protein among substances known to have antiviral activity against other infectious diseases and viruses. In comparison to the control molecule, it was determined that the majority of the compounds had considerable binding affinities and the potential to decrease RdRp activity, indicating their potential as treatment options. A total of 71 compounds have been identified to have a higher binding affinity than the control, but in this study, only the top 10 are reported (Table 1), and a detailed interaction analysis was performed on the top 5 molecules (Figure 1).
The comprehensive interaction analysis of the top five compounds was conducted, and they compared them to the control compound in terms of their binding poses and the residues that interact with the most crucial residues of the target protein. The fact that all five of the compounds that were selected, as well as the control (shown in blue in Figure 1), are located within the same grid and share the majority of the same residues is made readily apparent by the visual inspection of their binding poses (Figure 1).
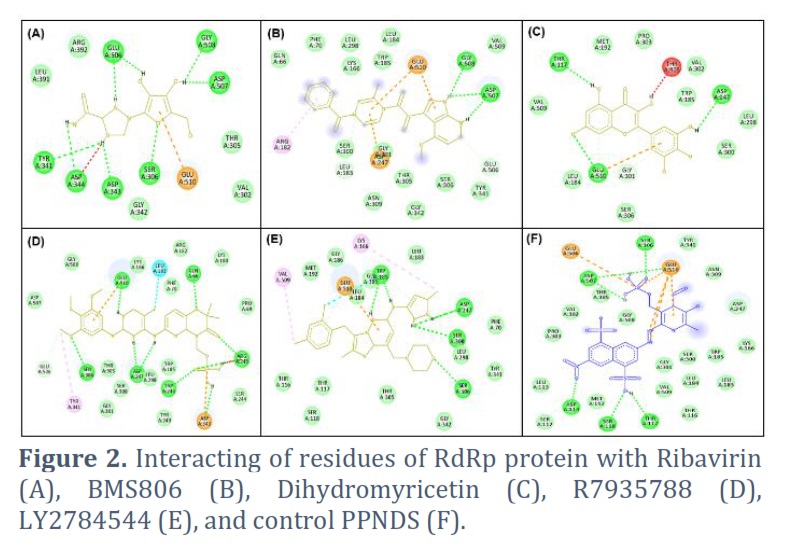
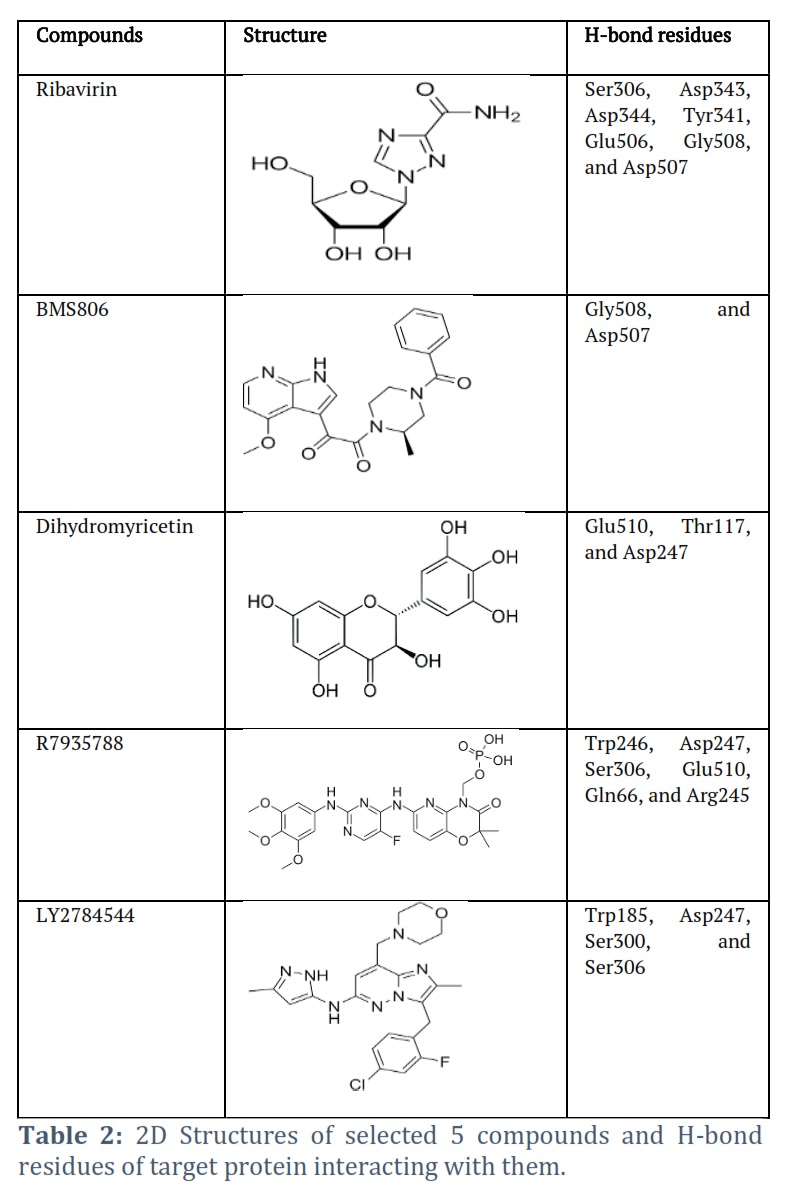
Ribavirin was found to interact with Glu510, Ser306, Gly342, Asp343, Asp344, Tyr341, Leu391, Arg392, Glu506, Gly508, Asp507, Thr305, and Val302 residues of RdRp protein. The Ser306, Asp343, Asp344, Tyr341, Glu506, Gly508, and Asp507 residues of RdRp protein were observed to H-bonded with Ribavirin (Figure 2A). BMS806 interacted with Asp247, Gly301, Leu183, Ser300, Arg182, Gln66, Phe70, Leu298, Lys166, Trp185, Leu184, Glu510, Gly508, Asp507, Val509, Glu506, Tyr341, Ser306, Gly342, Thr305, and Asn309 of RdRp protein. The Gly508, and Asp507 residues of RdRp protein were H-bonded with BMS806 (Figure 2B). Dihydromyricetin was found to interact with Ser306, Gly301, Glu510, Leu184, Val509, Thr117, Met192, Pro303, Thr305, Val302, Trp185, Asp247, Leu298, and Ser300 residues of RdRp protein. The Glu510, Thr117, and Asp247 residues of RdRp protein were observed to H-bonded with Dihydromyricetin (Figure 2C). R7935788 interacted with Tyr341, Ser306, Glu506, Asp507, Gly508, Glu510, Lys166, Leu183, Phe70, Gln66, Arg182, Gln66, Lys180, Pro69, Arg245, Ser244, Asp343, Tyr243, Trp246, Trp185, Leu298, Asp247, Ser300, Gly301, and Thr305 residues of RdRp protein. The Trp246, Asp247, Ser306, Glu510, Gln66, and Arg245 residues of RdRp protein were H-bonded with R7935788 (Figure 2D). Further, LY2784544 was found to interact with Thr305, Thr117, Ser118, Thr116, Val509, Met192, Gly186, Glu510, Leu184, Gly301, Trp185, Lys166, Leu183, Asp247, Ser300, Phe70, Leu298, Tyr341, Ser306, and Gly342 residues of RdRp protein. The Trp185, Asp247, Ser300, and Ser306 were H-bonded with LY2784544 (Figure 2E).
The co-crystallized ligand (PPNDS) was found to interact with Glu506, Ser306, Glu510, Tyr341, Asn309, Asp247, Lys166, Trp185, Leu183, Ser300, Leu184, Thr116, Val509, Gly301, Thr117, Ser118, Met192, Asp114, Ser112, Leu113, Pro303, Val302, Gly508, Thr305, and Asp507 residues of RdRp protein (Figure 2F). Structures of selected compounds has been shown in Table 2.
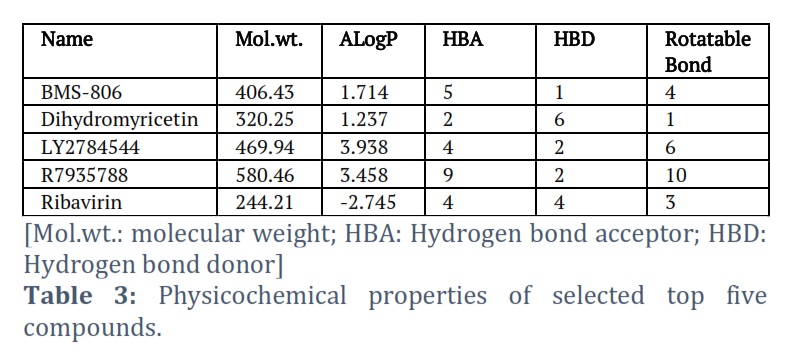
The physicochemical characteristics of selected five compounds have been presented in Table 3.
Figures & Tables
This study employs computational screening methodologies to identify potential NV RdRp protein inhibitors among substances with established antiviral efficacy against various infectious diseases and viruses. The investigation showed most of these compounds had significant binding affinities, implying their ability to inhibit RdRp activity and thus their potential as therapeutic agents. Seventy-one of the screened compounds had higher binding affinities than the control molecule. However, this study focuses only on the top ten compounds, and a thorough interaction analysis was performed on the top five compounds.
In order to gain a deeper understanding of the interacting residues between RdRp and the identified hit compounds, an analysis was conducted by re-docking the co-crystallized ligand (PPNDS) with RdRp. This analysis revealed that Glu506, Ser306, Glu510, Tyr341, Asn309, Asp247, Lys166, Trp185, Leu183, Ser300, Leu184, Thr116, Val509, Gly301, Thr117, Ser118, Met192, Asp114, Ser112, Leu113, Pro303, Val302, Gly508, Thr305, and Asp507 residues played a crucial role in the binding of PPNDS to RdRp. Interestingly, the hit compounds (Ribavirin, BMS806, Dihydromyricetin, R7935788, and LY2784544) were found to bind to these same RdRp residues, indicating that these compounds interacted within the same RdRp pocket as PPNDS.
The ligand binding efficiency with the target protein can be elucidated by considering the minimal binding affinity obtained from docking [15-18]. In comparison to the positive control PPNDS, the hit compounds Ribavirin, BMS806, Dihydromyricetin, R7935788, and LY2784544 exhibit significantly lower binding affinities. The results suggest a strong interaction between the RdRp protein and these hit compounds.
R7935788, frequently referred to as Fostamatinib (R788), is a highly effective inhibitor of spleen tyrosine kinase, with an IC50 value of 41 nM. It exerts a potent inhibitory effect on Syk, while having minimal influence on Lyn. Moreover, it exhibits approximately one-fifth of the potency compared to Flt3. Significantly, Fostamatinib has progressed to Phase 3 clinical trials, representing a noteworthy advancement in its development [19].
BMS-378806 is a selective inhibitor that hinders the binding between HIV-1 gp120 and the CD4 receptor. The CD4 receptor is the specific target of this binding. During viral research testing, it exhibited an exceptional EC50 range of 0.85-26.5 nM. BMS-378806 demonstrates its potential as a potent anti-HIV-1 medication by selectively inhibiting the interaction between gp120 and the CD4 receptor [20]. Ampelopsis grossedentata synthesizes dihydromyricetin, which is a naturally-occurring flavonoid antioxidant. It is alternatively referred to as Ampelopsin and Ampeloptin [21].
LY2784544, often referred to as Gandotinib, is a highly potent JAK2 inhibitor with an exceptional IC50 value of 3 nM. When comparing it to JAK2V617F, it exhibits a selectivity that is 8 times more than JAK1 and 20 times greater than JAK3, suggesting exceptional effectiveness in suppressing JAK2V617F. Gandotinib has progressed to Phase 2 of clinical trials, signifying a substantial advancement in the medicine's development and assessment [22]. Finally, Ribavirin, a synthetic analogue of guanosine, has been shown to have a broad spectrum of antiviral activity against both DNA viruses as well as RNA viruses [23].
Human NV is the most common pathogen responsible for acute gastroenteritis worldwide. RdRp plays an important role in viral replication and may be a promising target for the development of anti-NV therapeutics. Here, antiviral compounds were screened against the NV RdRp protein. Ribavirin, BMS806, Dihydromyricetin, R7935788, and LY2784544 were discovered to bind strongly to RdRp protein. Notably, these compounds had much lower binding affinities than PPNDS. These compounds also shared several RdRp protein binding residues with PPNDS. These results suggest that these compounds could be used as future NV RdRp inhibitors in the development of antiviral medications.
Conflict of Interest
The authors declare that there is no conflict of interest.
LSA, ASA and QA; Conceptualization, Data collection/curation, writing the manuscript, AA, RMA and WAA; Data collection/curation, analysis, MAK, AHA and ASA; extensive editing of the first draft.
![]() References
References
- Koo HL, Ajami N, Atmar RL, DuPont HL. Noroviruses: The leading cause of gastroenteritis worldwide. Discovery Medicine, (2010); 10(50): 61-70.
- Lopman BA, Steele D, Kirkwood CD, Parashar UD. The Vast and Varied Global Burden of Norovirus: Prospects for Prevention and Control. PLOS Medicine, (2016); 13(4): e1001999.
- Hall AJ. Noroviruses: the perfect human pathogens? The Journal of Infectious Diseases, (2012); 205(11): 1622-1624.
- Kaufman SS, Green KY, Korba BE. Treatment of norovirus infections: moving antivirals from the bench to the bedside. Antiviral Research, (2014); 105: 80-91.
- Rocha-Pereira J, Neyts J, Jochmans D. Norovirus: targets and tools in antiviral drug discovery. Biochemical Pharmacology, (2014); 91(1): 1-11.
- Ebenezer O, Jordaan MA, Damoyi N, Shapi M. Discovery of Potential Inhibitors for RNA-Dependent RNA Polymerase of Norovirus: Virtual Screening, and Molecular Dynamics. International Journal of Molecular Sciences, (2020); 22(1): 171.
- Yates MK, Seley-Radtke KL. The evolution of antiviral nucleoside analogues: A review for chemists and non-chemists. Part II: Complex modifications to the nucleoside scaffold. Antiviral Research, (2019); 162: 5-21.
- Mastrangelo E, Pezzullo M, Tarantino D, Petazzi R, Germani F, et al. Structure-based inhibition of Norovirus RNA-dependent RNA polymerases. Journal of Molecular Biology, (2012); 419(3-4): 198-210.
- S MAK, Hosen MA, Ahmad S, El Bakri Y, Laaroussi H, et al. Potential SARS-CoV-2 RdRp inhibitors of cytidine derivatives: Molecular docking, molecular dynamic simulations, ADMET, and POM analyses for the identification of pharmacophore sites. PLoS One, (2022); 17(11): e0273256.
- Bassetto M, Van Dycke J, Neyts J, Brancale A, Rocha-Pereira J. Targeting the Viral Polymerase of Diarrhea-Causing Viruses as a Strategy to Develop a Single Broad-Spectrum Antiviral Therapy. Viruses, (2019); 11(2): 173.
- Tarantino D, Pezzullo M, Mastrangelo E, Croci R, Rohayem J, et al. Naphthalene-sulfonate inhibitors of human norovirus RNA-dependent RNA-polymerase. Antiviral Research, (2014); 102: 23-28.
- Kireev D. Structure-Based Virtual Screening of Commercially Available Compound Libraries. Methods in Molecular Biology, (2016); 1439: 65-76.
- Kontoyianni M. Docking and Virtual Screening in Drug Discovery. Methods in Molecular Biology, (2017); 1647: 255-266.
- Dallakyan S, Olson AJ. Small-molecule library screening by docking with PyRx. Methods in Molecular Biology, (2015); 1263: 243-250.
- Sait KHW, Mashraqi M, Khogeer AA, Alzahrani O, Anfinan NM, et al. Molecular docking analysis of HER-2 inhibitor from the ZINC database as anticancer agents. Bioinformation, (2020); 16(11): 882-887.
- Sait KHW, Alam Q, Anfinan N, Al-Ghamdi O, Malik A, et al. Structure-based virtual screening and molecular docking for the identification of potential novel EGFRkinase inhibitors against ovarian cancer. Bioinformation, (2019); 15(4): 287-294.
- Sayed Murad HA, M MR, Alqahtani SM, B SR, Alghamdi S, et al. Molecular docking analysis of AGTR1 antagonists. Bioinformation, (2023); 19(3): 284-289.
- I JH, Alsharif FH, Aljadani M, Fahad Alabbas I, Faqihi MS, et al. Molecular docking analysis of KRAS inhibitors for cancer management. Bioinformation, (2023); 19(4): 411-416.
- Baluom M, Grossbard EB, Mant T, Lau DT. Pharmacokinetics of fostamatinib, a spleen tyrosine kinase (SYK) inhibitor, in healthy human subjects following single and multiple oral dosing in three phase I studies. British Journal of Clinical Pharmacology, (2013); 76(1): 78-88.
- Lin PF, Blair W, Wang T, Spicer T, Guo Q, et al. A small molecule HIV-1 inhibitor that targets the HIV-1 envelope and inhibits CD4 receptor binding. Proceedings of the National Academy of Sciences of the United States of America, (2003); 100(19): 11013-11018.
- Liu D, Mao Y, Ding L, Zeng XA. Dihydromyricetin: A review on identification and quantification methods, biological activities, chemical stability, metabolism and approaches to enhance its bioavailability. Trends in Food Science & Technology, (2019); 91: 586-597.
- Ma L, Clayton JR, Walgren RA, Zhao B, Evans RJ, et al. Discovery and characterization of LY2784544, a small-molecule tyrosine kinase inhibitor of JAK2V617F. Blood Cancer Journal, (2013); 3(4): e109.
- Beaucourt S, Vignuzzi M. Ribavirin: a drug active against many viruses with multiple effects on virus replication and propagation. Molecular basis of ribavirin resistance. Current Opinion in Virology, (2014); 8: 10-15.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0