

Full Length Research Article
Alleviation of heat stress by ACC Deaminase and Exopolysaccharide Producing Plant-Growth-Promoting Rhizobacteria in Solanum melongena L.
Sami Hannachi1,2*, Karim Naghmouchi3,4, Angelo Signore5
Adv. life sci., vol. 11, no. 2, pp. 513-524, May 2024
*– Corresponding Author: Sami Hannachi (sami.hannachi2025@gmail.com)
Authors' Affiliations
2. Department of Plants and Crops, Faculty of Bioscience Engineering, Ghent University, Ghent – Belgium
3. College of Clinical Pharmacy, Department of Pharmaceutical Chemistry, Al Baha University Kingdom of Saudi Arabia
4. Université de Tunis El Manar, Faculté des Sciences de Tunis, LR01ES05 Biochimie et Biotechnologie, 2092, Tunis – Tunisie
5. Department of Agricultural and Environmental Science, University of Bari Aldo Moro, Via Amendola Bari – Italy
[Date Received: 14/12/2023; Date Revised: 27/01/2024; Date Available Online: 18/04/2024]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: The current study was carried out to assess the alternative role of high temperature tolerant plant growth promoting rhizobacteria (PGPR) in alleviation of heat stress adverse effect in eggplant.
Methods: PGPR was assessed for PGP potentialities and investigated for1‐aminocyclopropane‐1‐carboxylate (ACC deaminase), and exopolysaccharide under normal and high temperature environment. The impact of PGPR application on plant physiology and biochemistry was investigated under normal and high temperature environments and plant growth regulators were evaluated using High Performance Liquid Chromatography (HPLC).
Results: A significant impediment for morphological and physiological parameters of eggplant cultivars was observed during heat stress in absence of PGPR inoculation. The results showed that ACC‐deaminase produced by PGPR boosted up significantly the extracellular polymeric substances (EPS) accumulation, conversion of ACC into α‐ketobutyrate and ammonia, and reduce consequently the impact of high temperature on eggplant development.
Conclusion: PGPR inoculation showed an alternative strategy to improve eggplant growth and development under high temperature condition in order to preserve agriculture sustainability and healthy crops.
Keywords: ACC‐deaminase; Extracellular polymeric substances; Heat stress; PGPR; Solanum melongena
Introduction![]()
Plants are subjected to stressful environmental conditions caused by several abiotic and biotic stresses [1]. Extreme temperature, droughts, and high carbon dioxide are considered as serious detrimental environmental factors alerting plant morphological, biochemical and physiological processes. These parameters could be reversible or irreversible, depending on their duration and sternness and consequently affecting crop growth and productivity under global warming change [2-4]. It is expected that greenhouse gases will increase the Earth temperature by 0.3°C each ten years. Heat stress constitutes a critical threat for our sustainable farming system so huge and urgent effort has to be exerted to overcome the increasing damage of this abiotic stress on various crop species [5]. Elevated temperature is considered as a serious threat for various plant functioning processes such as plant germination, growth, physiology, reproduction, and yield [6, 7].
The vulnerability of plants to high temperature may depend on their developmental stage and the intensity of heat stress and its persistence [5, 8]. An elevated temperature ranging from 30 to 40°C may contribute to the disruption of plant germination kinetics [9]. Furthermore, it has been reported that plant reproduction is the most vulnerable plant function to heat stress for several crops such as lentil [10], Chickpea [11] and rice [12, 13]. Under stressful conditions (e.g., drought, salinity, heat) proline accumulation is known to be an osmoprotective strategy of membranes and proteins, developed by plants to keep their optimal growth and reproductive functions [14, 15]. Besides, proline has a key role in the effective scavenging of accumulated free radicals and is well known for its contribution in the cell osmotic adjustment under abiotic stresses [16, 17]. The onset of heat stress and its sternness impede the proline pathway and sugar transport during the plants reproductive stage thereby disrupting fruit formation and development [18].
Plant growth promoting rhizobacteria (PGPR) growing in the rhizosphere and rhizoplane of the plants are one of the most effective biological strategies developed by plants to counteract abiotic stress [19, 20]. PGPR are provided with 1-aminocyclopropane-1-carboxylate deaminase (ACC deaminase) activity [21, 22]. Consequently, the application of beneficial bacteria becomes a crucial practice aiming at enhancing the production of plant growth regulators thus improving nutrient uptake and inhibiting plant pathogens development [23, 24]. Under abiotic stress, PGPR has been reported in several crops such as cucumber [25], mung bean [26], wheat [27] and tomato [5].
The production of various plant growth hormones (indole‐3‐acetic acid, cytokinins, gibberellic acid and abscisic acid) and other growth-improving molecules (antibiotics, siderophore, nitrogenize) is considered an effective plant response to mitigate the negative effect of abiotic stress [28-30]. The impact of stress-induced plant oxidative damage can be alleviated by PGPR through the production of antioxidants, thus increasing the amount of abscisic acid (ABA) and reducing the free radical’s accumulation [23]. It has been reported that PGPR inoculation improves root growth and change root architecture, thereby promoting root surface area and enhancing mineral absorption accumulation [31, 32]. In addition, PGPR treatment is closely related to optimal shoot development, thus keeping the plant healthy and improving its productivity [26]. Eggplant (Solanum melongena L.) is considered as one of the most important vegetable crops with high economic value worldwide [33]. Several rhizospheric bacteria including Burkholderia, Bacillus, Azotobacter, Azospirillum, Enterobacter, Pseudomonas, and Serratia have been well known by their effective potential in pant growth promotion, mineral uptake improvement, plant hormone optimization and avoidance of pathogenic action in various solanaceous vegetable crops including eggplant [34-36].
In this paper we focused on screening heat tolerant PGPR strains provided with various PGP traits and assessing their impact on heat stress mitigation. To interpret the cultivar responses, we investigated growth, physiological and biochemical parameters of eggplant in a greenhouse experiment.
Methods![]()
Rhizospheric soil sampling and PGPR isolation
Two F1 hybrids of root samples (‘Black Beauty’& ‘Bonica’) eggplant cultivars were collected from 105 days old eggplant rhizosphere cultivated in the Department of Plant physiology and Biotechnology, National Institute of Agronomic Research, Tunis, Tunisia (36◦500 N, 10◦110 E). The highest and lowest average of Tunis temperature in summer was 25 and 40 °C, respectively. Sample soil characteristics zone was pH 8.5, electrical conductivity (EC) 0.35 dS m−1 and organic matter concentration 0.85%. The isolation of PGPR was achieved via serial dilution protocol, then placed on Luria‐Bertani (LB) agar medium and finally incubation at 37 °C [37]. The obtaining of pure culture was performed by collecting the selective colonies after being streaked and diluted several times.
High Temperature Tolerance Assay
The High temperature tolerance of PGPR strains was assessed on population density at temperatures varying from 40 to 60°C (40, 45, 50, 55, and 60°C) in LB medium. The bacterial strain inoculation was achieved in 200 mL sterilized flasks containing 100 mL LB medium and maintained at different temperatures ranging from 40 to 60 °C using shaking incubator (120 rpm). An incubation of 24 h was followed by recording the inoculated bacterial culture optical density at λ= 600 nm through a spectrophotometer (Agilent 8453 UV–visible Spectroscopy System).
Screening of in vitro traits of PGPR
Phosphate solubilizing microorganisms are routinely screened by a plate assay method using Pikovskaya (PVK) agar. Then the inoculation of the bacterium was conducted on PVK agar plates, and the incubation was achieved for one week at 30 °C [38]. The production of indole-3-acetic acid (IAA) was assessed by calorimetry [39] and the estimation of ammonia production was determined in peptone water following the method of Ahmad et al. [40]. The screening for siderophore production was determined via Chromeazurol S (CAS) agar media according to Schwyn et al. [41]. The quantification of hydrogen cyanide (HCN) production was performed following the protocol of Castric et al., [42] by streaking bacterial isolates on agar medium culture supplemented with glycine (4.4 g/L). After being soaked in solution containing 0.5% picric acid and 2% sodium carbonate a Whatman No. 1 filter was kept in the plate lid and then the incubation was conducted during seven days at 30 ± 0.1°C. HCN production was investigated through the occurrence of one of two colors (dark or light brown).
Evaluation of ACC deaminase producing PGPR strains
Qualitative and quantitative estimation
We investigated the ACC‐deaminase production as a nitrogen source for the PGPR strain according to the protocol of Glick et al. [43]. The growth of the bacterial isolate was performed in 5 mL of Tryptic Soy Broth (TSB) medium during 24 hours at 28°C with uninterrupted quaking at 120 rpm. A centrifugation of cells was conducted for 5 min at 3000 g, then rinsed with sterilized water and maintained in 0.1M Tris‐HCl (pH 7.5). Finally, we inoculated the bacterial strain in petri plates containing Dworkin and Foster salt medium culture added or not with ACC [44]. Plates containing ammonium sulphate were used as a positive control. The incubation was conducted at 28 ± 2°C for three days. The growth of strains isolates on ACC-added plates were compared with controls (the negative and positive one) [44]. After qualitative assay, a quantitative estimation of ACC deaminase activity was performed following the protocol of Penrose and Glick [45].
Exopolysaccharide production evaluation
The PGPR ability of EPS production was investigated according to the protocol of Mu’minah and Subair [46].
Extracellular Enzyme Activities
The determination of extracellular enzyme activities was carried out to assess enzyme bacterium potentialities. Protease assay, amylase assay and pectinase test were performed according to the protocol of Kazempour [47], Adebiyi and Akinyaanju [48] and Namasivayam et al. [49], respectively.
For catalase assay, a sample of bacterial strain was taken, deposited on microscopic slide and made smear using autoclaved loop. After adding some H2O2 (3%) drops an occurrence of gas bubbles was noticed [50]. The positive indication of catalase was evidenced by increasing generation of bubbles, whereas a low bubbles appearance indicated the negative result for catalase assay.
PGPR microbial identification
Biochemical traits of PGPR strains were tested using bacterial isolation kits QTS‐24 miniaturized recognition process (Biology Laboratory, Hail University, Saudi Arabia) according to the method of Yasmin and Bano [51]. Bacterial isolated were supplemented the QTS wells and then incubated at 37 °C for 24 hours. Later on, the QTS wells inoculated with bacterial culture were added with reagents following the QTS‐24 kit manual and finally observations were made.
Quantitative evaluation of Plant Growth Regulators
The quantitative assessment of bacterial strain IAA, gibberellins and kinetin was conducted according to the protocol of Saber et al. [53]. The growth of bacterial strain was performed in King’s B broth medium in presence or absence of heat stress in quaking incubator (SHKE480HP, Thermo Fisher Scientific, Kansas City, USA) at 155 rpm for 5 days. Then the obtained supernatant was kept, and the rest was thrown. The reduction of the supernatant volume to 20 mL was carried out via a rotary evaporator (Yamato RE500, Yamato Scientific, Japan). The supernatant pH adjustment to 2.8 was achieved using 1N HCl. The primary extraction phase was conducted by centrifugation 3 times at 800× g (IEC HN‐SII, Thermo Fisher Scientific, USA) during 5 min using ethyl acetate (1:1; v/v). The upper constituent of ethyl acetate was taken following each centrifugation. The adjustment of pH water phase to7 was done with 1N NaOH and the extraction was performed with water‐saturated N‐butanol (0.4 ml) during 5 min at 800× g for 3 times. The obtained higher butanol portion was kept, and the rest was removed. After that the mixture was evaporated at 55 °C and the dilution (in 2.0 mL of MeOH:H3P04; 99.9:1; v/v) of the obtained ethyl acetate and N‐butanol phases were carried out. After dilution and centrifugation of the extract during 10 min at 12,000x g (SorvallBiofuge Pico, Kendro Laboratory Products, Germany), the particles of bacteria were eliminated. The quantification of IAA, gibberellic acid (GA3) and kinetin was performed using high performance liquid chromatography (HPLC) processed with Waters 2487 Dual λ absorbance detector and column (Vydac 218Tp C18 5 μm). The detection was performed at 214 nm. The identification of IAA, GA3 and kinetic on chromatograms and the determination of their concentration were made using the analytical grade hormones (Sigma Aldrich, USA).
Greenhouse Assay
Seed sterilization and treatment with PGPR strain
Two F1 hybrids (‘Bonica’ and ‘Black Beauty’) eggplant cultivars were used as plant material. The seeds surface was sterilized with 70% ethanol and rinsed with distilled water. Then, they are dipped in a solution of 0.02% Dreft (5–15% non-ionic surfactants, 15–30% anionic surfactants) and 5% HazTab (1,3,5 Dichloro-Triazine-Trione dihydrate-dichloro sodium) for 20 min and followed by a second dipping in a solution of mercuric chloride (0.5%) during 10 min [54]. The assumed PGPR strain was inoculated and cultivated in LB media which is a mixture of NaCl (10 g) (Sigma Aldrich), tryptone (10 g) (Sigma Aldrich) and yeast extract (5 g) (Sigma Aldrich) in of H2O (1 liter) during 24 h and then the mixture was centrifuged during 10 min at 3000 rpm.
Experimental setup
The study was conducted at the Department of Plant Physiology and Biotechnology (National Institute of Agricultural Research, Tunisia).Four treatments were applied in our experiment.T0 = control (absence of bacteria / normal temperature), T1 = treated plant with PGPR strain, T2 = plants under normal temperature and T3 = plants inoculated with bacteria under high temperature. A factorial completely random design was used in all analyses with five replicates for every treatment and variety. Seeds of eggplant were sown into plug trays with peat as substrate and placed in a growth chamber with the following parameters T 25 °C, RH 70%, photon flux density of 150 μmol m−2 s−1 and photoperiod of 16 h. Later on, the seedlings were transplanted into 2 L plastic pots at the occurrence of the second true leaf. Plants were shifted to a greenhouse with a minimum temperature setpoint of 21 °C (located at 51°02′N, 03°42′E) and were fertigated with 250 mL of full-strength Hoagland’s solution [55], twice a week. Plants growing in the presence/absence of bacterial treatment were subjected to high temperature (42 °C) at flowering phase. Then plants were exposed to high temperature during 6 h/day in a growth chamber until reaching the fruit formation phase. Following the application of heat stress plants were transferred one more time to greenhouse for recuperation (temp. 25 ± 2 °C). 45 days after flowering stage four plants were taken randomly, cleaned with sterile demineralized water then kept for further analysis. Plants were harvested 105 days after sowing. The harvested roots plants were washed with sterilized water to achieve the elimination of roots debris and kept for future investigation.
Plant Growth and Relative Water Content (RWC) measurements
Using the randomly collected plants we measured root and shoot length. Fresh weight (FW) was determined. The surface area of the leaf was measured by a leaf area meter. Dry weight (DW) was evaluated after 48 h drying at 60 °C in a forced draft oven. The assessment of leaf RWC was achieved following the method of Barrs et al. [56]. We measured the leaf fresh weight (FW), turgid weight (TW), and dry weight (DW), and we calculated RWC following the formula: RWC (%) = [(FW − DW)/(TW − DW)] × 100.
PGPR assessment effect on photosynthetic pigments in the presence/absence of heat stress
The pigments extraction was conducted using 0.20 g fresh weight supplemented by 80% acetone (v/v). The mixture was maintained in the dark atmosphere at room temperature until the tissue of leaf was totally bleached. The centrifugation of the obtained extract was achieved at 5000× g during 5 min and then supplemented by 80% acetone to reach a final volume of 15 mL. The measurement of extracts absorbance was carried out at optical density of 645 and 663 nm via a spectrophotometer (UV-2550, Shimadzu, Japan). Finally, every pigment concentration was estimated as described by Arnon [57] and Lichtenthaler [58] and expressed in µg g-1 fresh weight (FW).
For protein extraction and quantification fresh plant tissue were homogenized with 50 mM potassium phosphate buffer (pH 7) (3 mM MgCl2, 1 mM EDTA and 1.0% PVP, w/v). The homogenates were centrifuged at 10,000 rpm during 20 min at 4 °C. Protein content determination was performed following Afridi et al. [59] and bovine serum albumin was used as standard. Proline quantification was achieved according to Bates et al. [60]. Briefly, we extracted 500 mg of plant tissue using 3% (w/v) sulfosalicylic acid and Proline concentration was determined.
For enzymatic assay, we measured the guaiacol peroxidase (POD), superoxide dismutase (SOD) and Catalase (CAT) activities according to the protocols of Chance and Maehly [61], Lu et al. [62] and Aebi (1984) [63], respectively. Results were stated as EU mg−1 of protein.
Statistical analyses
Data were subjected to a two-way analysis of variance (ANOVA) approach for the two cultivars based on bacteria (presence /absence of bacteria inoculation) and thermic level (presence /absence of high temperature stress). All analysis was achieved using a completely random design. The identification of statistically significant dissimilarities between the treatments and varieties was carried out using the software of SPSS Statistics 25 (IBM SPSS Statistics). Means comparison was performed through Tukey’s multiple range test (p = 0.05).
Results![]()
Isolation of bacteria and assessment of their tolerance potential for heat stress
The isolation bacteria from roots and their purification were performed. Then bacterial isolates were subjected to heat tolerance screening up to 60°C. Only 13, 8 and 3 strains succeeded to survive at 50°C, 55 °C and 60°C, respectively. Based on its superiority in term of PGP activities and elevated temperature resistance, only one bacterial strain was chosen from these three strains that were able to survive at 60°C.
Morphological, physiological and biochemical characterization of the selected bacteria
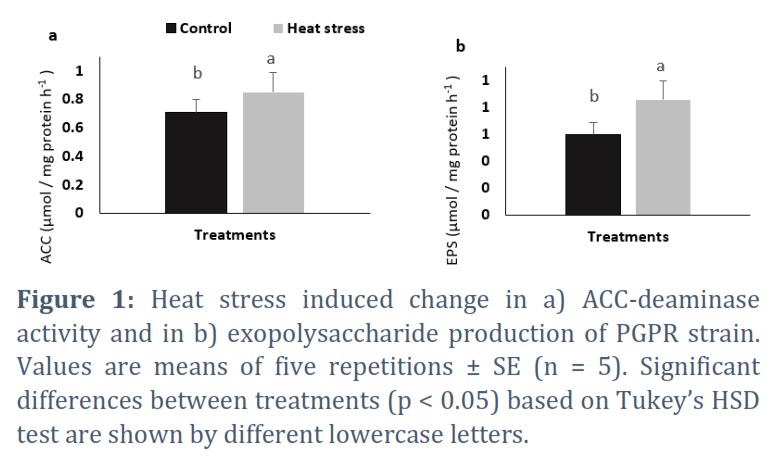
The selected bacterial strain was specified as Gram positive, and rod shaped through conducting Gram staining method. Moreover, the selected bacterial strain was investigated via microbial identification QTS‐24 kits (Biology department laboratory, college of Science, Hail University). Morphological traits were examined using compound microscope and naked eye. Furthermore, biochemical characteristics evaluation (notably PGRP activities) was achieved for the studied bacterial strain. Our results showed that the strain exhibited positivity for catalase, amylase, pectinase and protease synthesis, but was negative for hydrogen cyanide and siderophore production. The selected strain showed ACC‐deaminase activity and exopolysaccharide production under control (normal temperature) and stressful environment (elevated temperature). The isolated strain was recorded to exhibit higher growth in presence/absence of nitrogen source. The selected bacteria grown on DF medium free of ACC precursor exhibited lower growth when compared to positive control and plates containing ACC. The bacterial strain grown under high temperature condition (presence of heat) was found to be more efficient as ACC deaminase and EPS producer than under normal temperature (absence of heat) (Figure 1).
The analysis of PGPR hormone accumulation under normal and heat stress condition was studied via High performance liquid chromatography (HPLC). Recorded Gibberellic acid (GA) concentration was 21.7 μg/mL under normal temperature and 16.2 μg/mL under elevated temperature. While monitoring the IAA production, the selected bacterial strain showed a concentration of 0.60 and 0.49 μg/mL in absence and in presence of heat stress, respectively. Further, the kinetin quantification showed a concentration of 49.1 and 30.2 μg/mL under normal and heat condition, respectively.
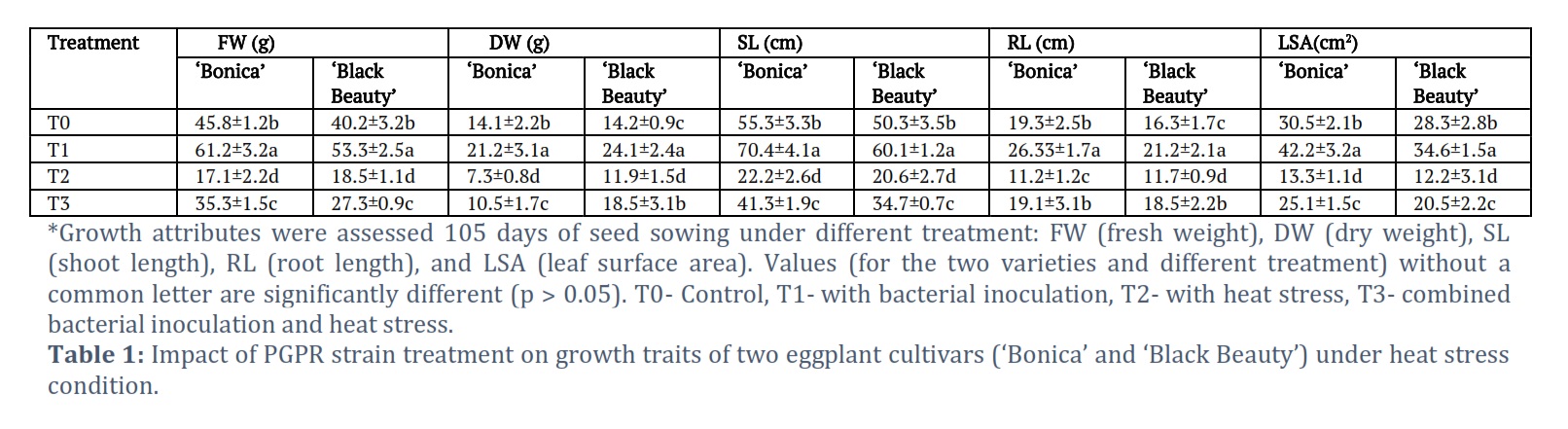
Impact of PGPR strain inoculation on plant biomass
PGPR strain inoculation (T1) enhanced fresh and dry biomass in ‘Bonica’ and ‘Black Beauty’ by 33.6%, 50.3%, 32.5% and 69.7%, respectively when compared to non-treated control (T0) under normal temperature. Whereas heat stress reduced fresh and dry biomass in ‘Bonica’ and ‘Black Beauty’ by 62.6%, 48.2%, 53.9% and 16.19%, respectively when compared to un-inoculated control (T0). Further, bacterial inoculation under heat stress rose fresh and dry biomass in ‘Bonica’ and ‘Black Beauty’ by 26.4%, 50.3%, 32.5% and 69.7%, respectively when compared to heat stress condition without bacterial inoculation (T2). In absence of heat stress, the bacterial treatment (T1) rose shoot length by 26.8% and 19.5% in ‘Bonica’ and ‘Black Beauty’ when compared to the non-treated control (T0) under no heat stress. However, high temperature decreased shoot length by 59.8% and 59.04% in ‘Bonica’ and ‘Black Beauty’ when compared to un-inoculated control (T0), respectively (Table 1). In addition, bacterial treatment under high temperature (T3) increased the shoot length by 86.03% and 68.4% in ‘Bonica’ and ‘Black Beauty’, respectively when compared to bacterial un-treatment under heat stress condition (T2). The same trend was obtained when dealing with root length. Bacterial inoculation under normal temperature (T1) increased root length in ‘Bonica’ by 36.3% and in ‘Black Beauty’ by 30.06% in comparison to un-inoculated control (T0). Moreover, bacterial treatment under high temperature (T3) enhanced root length in ‘Bonica’ by 70.5% and in ‘Black Beauty’ by 58.11% when compared to un-inoculated heat stress condition (T2).
Likewise, the T1 (bacterial inoculation under normal temperature) boosted-up leaf surface area in ‘Bonica’ by 38.3% and in ‘Black Beauty’ by 22.2% when compared to T0 (untreated control). In addition, T3 (bacterial treatment under high temperature) increased leaf surface area by 88.7% and 68.03% in ‘Bonica’ and ‘Black Beauty’, respectively when compared to T2 (un-inoculated heat stress condition).
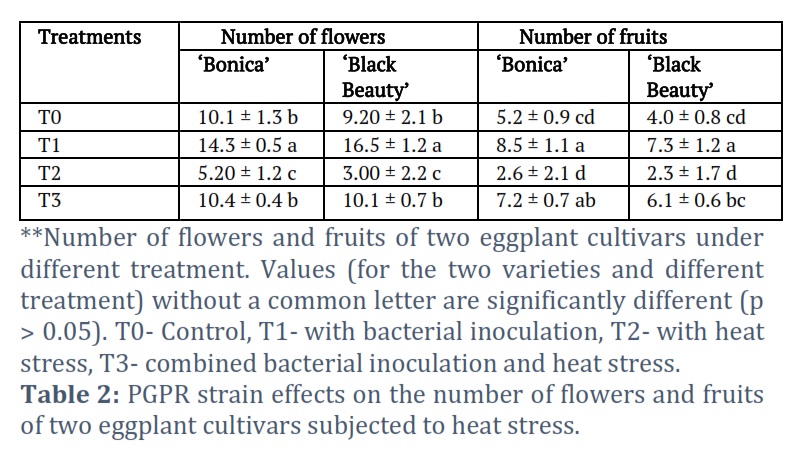
Also, bacterial inoculation (T1 and T3) contributed to superior activity in other PGPR traits such as flower numbers and numbers of fruits in both cultivars (‘Bonica’ and ‘Black Beauty’) during normal and heat stress conditions in comparison to T0 and T2 (un‐inoculated treatments) (Table 2). The quantitative estimation showed a significant decrease (p=0.05) in the number of flowers of ‘Bonica’(48.5%)in absence of bacterial treatment under heat stress when compared to control (T0). In addition, the exposition of ‘Bonica’ to combined normal temperature and bacterial treatment rose the number of flowers by 41.5% in comparison to control. Furthermore, T3 (the combined bacterial inoculation and heat stress) increased flowers number by 50% when compared to T2 (heat stress without bacterial inoculation). Moreover, in ‘Black Beauty’, the number of flowers was found to be reduced by 67.4% when exposed to high temperature without bacterial inoculation (T2) in comparison to control condition (T0). However, bacterial treatment generated an elevation of 44.2% in flowers number if compared with control condition (Table 2). Added to that, bacterial treatment (T3) boosted up the number of flowers by 9.8% in comparison to control and by 236.6% when compared to T2 (high temperature with bacterial inoculation). It was found that the selected bacterial inoculation enhanced fruits number under normal and stressful environment (T1 and T3) in comparison to control and non-treated plants exposed to high temperature (T2). The non-treated ‘Bonica’ plants, cultivated under high temperature (T2) exhibited 50% decrease in number of fruits in comparison to control (T0), and under normal temperature the plants with no bacterial treatment showed a greater fruit number (+38.2%) compared to control (T0). Bacterial treatment under high temperature (T3) increased the number of
fruit of 38.4% and 173.9% in comparison to control (T0) and non-treated plants under high temperature (T2), respectively. The non-treated ‘Black beauty’ plants subjected to high temperature (T2) exhibited lower fruit production in comparison to control (T0), and the treated plants subjected to normal temperature (T3) produced a greater number of fruit (80.5% more) compared to the control (T0).Combined bacterial treatment and high temperature (T3) boosted-up the number of fruits by 52.5% and 165.2% in comparison to non-treated ‘Black beauty’ exposed to high temperature condition (T1), respectively.
Leaf Relative Water Content (RWC)
RWC levels showed a reduction under normal and high temperature in the cultivars (‘Bonica’ and ‘Black Beauty’). Bacterial treatment (T1) increased RWC by 34.1% and 53.6% in ‘Bonica’ and ‘Black Beauty’, respectively, in comparison to control. Furthermore, associated bacterial treatment and high temperature (T3) boosted up RWC by 23.7% and 36.3% in ‘Bonica’ and ‘Black Beauty’, respectively, when compared to control.
Leaf Chlorophyll (chl) Contents
The exposure of to both varieties to treatment T2 (non-treated plants under high temperature) reduced both Chla and Chlb and carotenoid content, when compared to control (T0). In addition, bacterial treatment, significantly enhanced Chla and Chlb concentration and carotenoid accumulation in both cultivars under normal and high temperature condition.
The application of treatment T1 (bacterial inoculation under normal temperature) increased Chla and Chlb and carotenoid contents in ‘Bonica’ by 17.8%, 16.6% and 116.3%, respectively, in comparison to control (T0) . Treatment T3 (bacterial treatment under high temperature) augmented Chla and Chlb and carotenoid contents of ‘Bonica’ by 209.5%, 150.7% and 153.9%, respectively, when compared to treatment T2 (no bacterial inoculation with high temperature. The same trend was obtained with ‘Black Beauty’ cultivar. Bacterial treatment (T1) rose Chla and Chlb and carotenoid concentration by 28.4%, 25.6% and 117.4%, respectively relative to control (T0). Added to that bacterial treatment under high temperature (T3) increased leaf Chla and Chlb and carotenoid content by 204.1%, 66.6% and 380.5%, respectively when compared to T2 (non-treated plants under high temperature).
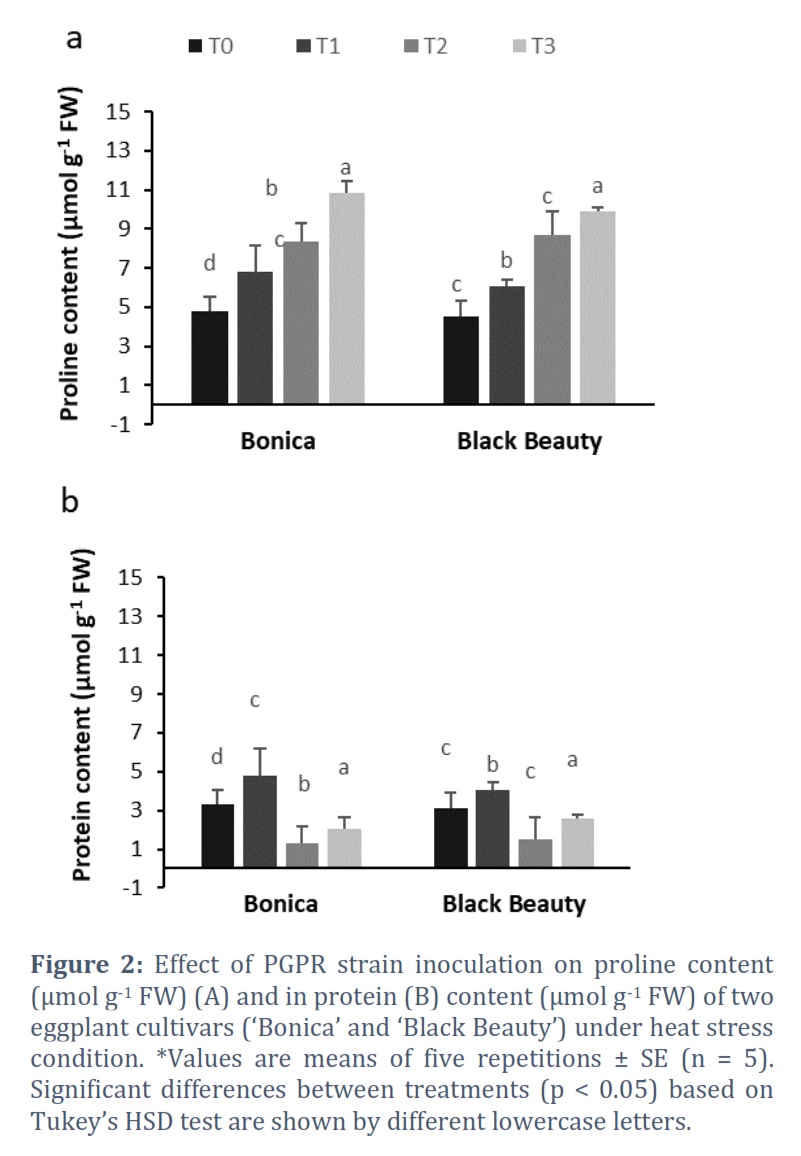
Protein and Proline Contents
Bacterial inoculation boosted up proline and protein biosynthesis in both cultivars under normal and high temperature (Figure 2). Under normal temperature, bacterial treatment enhanced proline and protein accumulation in ‘Bonica’ by 70.1% and 44.84%, respectively in comparison to control (T0). When subjected to bacterial treatment and high temperature, proline and protein content for ‘Bonica’ rose reached approximately 30.1% and 57.6%, respectively, in comparison to non-treated plants under high temperature. Under normal conditions, bacterial treatment (T1) boosted up proline and protein content in ‘Black Beauty’ by 33.5% and 30.3% respectively, relative to control (T0). Treatment T3 (bacterial treatment under heat stress) augmented proline and protein accumulation in ‘Black Beauty’ by 12.6% and 73.3%, respectively, when compared to treatment T2.
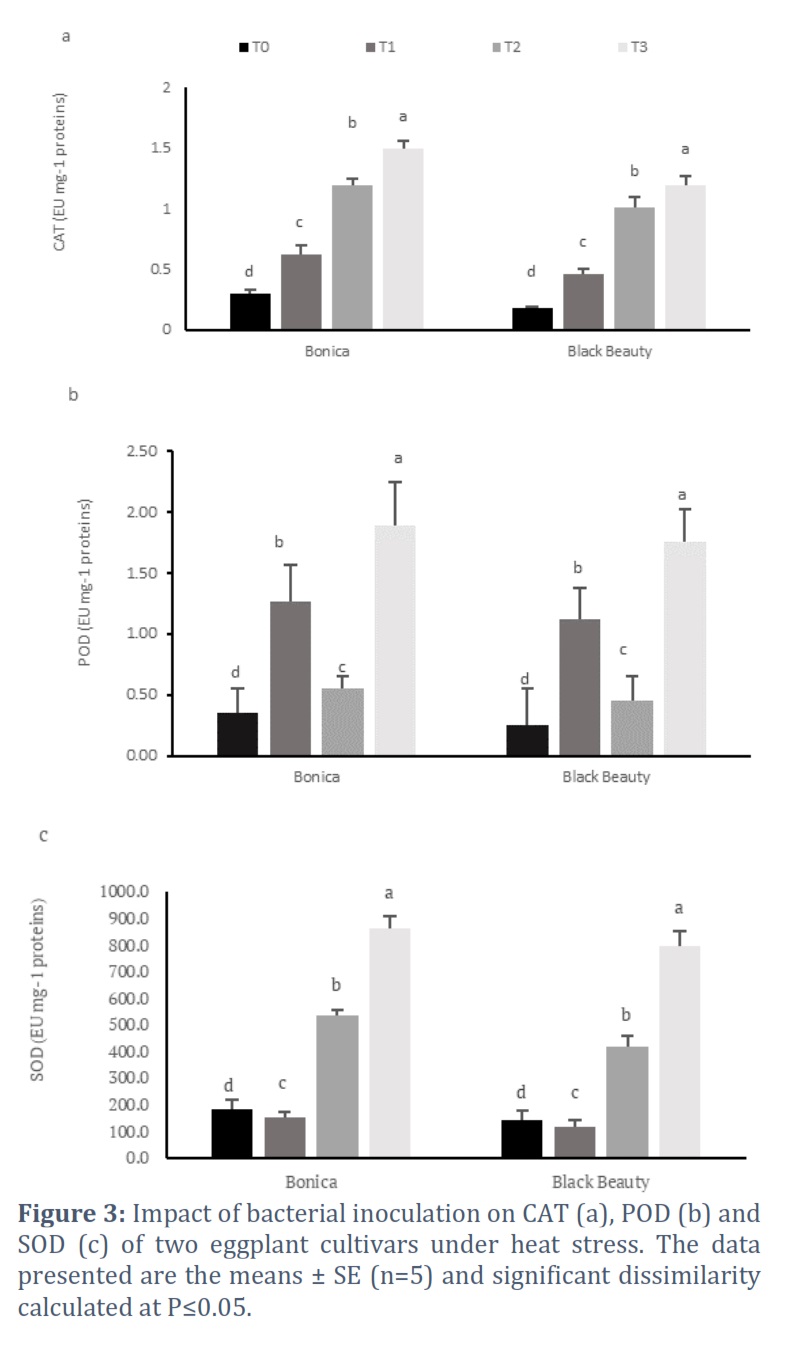
Induced change in the activity of antioxidant enzymes pathway
Figure 3 showed the impact of bacterial strain inoculation on the antioxidant enzymes pathway in eggplant cultivars. Results demonstrated that the exposition of both varieties to heat stress and bacterial treatment enhanced significantly the activity of all studied antioxidant enzymes (CAT: catalase, POD: peroxidase and SOD: superoxide dismutase).
The treatment T2 (non-inoculated plants under high temperature) rose CAT, POD and SOD activities 3-, 1.6 and 2.9-fold, respectively in ‘Bonica’ in comparison to control (T0) (Figure 3). Similarly, the treatment T1 (combined normal temperature and bacterial treatment) increased CAT and POD activities 2- and 3.6 -fold, respectively, in ‘Bonica’ relative to control (T0). Likewise, treatment T3 (bacterial treatment under heat stress), enhanced CAT, POD and SOD activities 1.3-, 3.4 and 1.6-fold, respectively in ‘Bonica’ compared to T2 (non-treated ‘Bonica’ plants under high temperature). When subjected to treatment T2 (untreated plants under heat stress), ‘Black Beauty’ increased CAT, POD and SOD activities by 5.6-, 1.8 and 2.9-fold, respectively in comparison to control (T0) (Fig.3). Added to that, treatment T1 (associated normal temperature and bacterial inoculation) boosted up CAT and POD and SOD activities 0.6- and 5 -fold, respectively in ‘Bonica’ in comparison to control (T0). In addition, our results showed that under treatment T3 (high temperature coupled bacterial treatment), the CAT, POD and SOD activities of ‘Black Beauty’ plants were found to be strengthened 1.2-, 3.9 and 1.9-fold, respectively, relative to treatment T2 (untreated plants under heat stress) (Figure 3).
Figures & Tables
During their development, plants established many suitable strategies to minimize the negative impacts of stressful conditions [64, 65]. The PGPR inoculation showed a distinguished contribution in the amelioration of the plant healthy status and high capacity in tolerating several abiotic and biotic threats [66, 67, 68]. Due to greenhouse effect generated by increased levels of carbon dioxide, chlorofluorocarbon, the occurrence of global warming was combined with numerous potential critical dangers [5, 69, 70]. The selected bacterial strain that exhibited higher potential was assessed in term of heat tolerance level and plant growth performance in presence or in absence of heat stress.
The present study deals with the aptitude of heat tolerant PGPR strain in mitigation of heat stress and amelioration of plant growth parameters in two eggplant cultivars through the generation of ACC‐deaminase, EPS and enzymes activities. Hence, an investigation was carried out to assess the efficacy of application of PGPR strain to mitigate the adverse impact of heat stress in eggplant in the current study. Under stressful conditions, plants showed that they may enhance the amount of ethylene in their tissues. Remarkably, Ethylene content boosted up in cells under stress conditions [71, 72, 73, 74]. Under abiotic stress condition, a high ethylene production engendered root growth impediment and plant development disruption. The ACC deaminase-producing bacterial strain contributed to ethylene generation mitigation, thus converting ACC into ammonia and α-ketobutyrate and alleviated the negative effect on plant development parameters under heat stress. The inoculation with selected PGPR isolates has contributed to the improvement of plant growth features when subjected to abiotic stresses [5, 67, 68]. Moreover, bacterial inoculation ameliorates plant mineral nutrition and root development [5, 75]. It is noteworthy that under normal and stressful environment, PGPR strain has the capacity to produce exopolysaccharides (EPS). Exopolysaccharide (EPS) production deeply plant tolerance to abiotic stress through improving water permeability, roots mineral nutrition, soil integrity, plant biomass production [76]. Our results regarding investigation of PGR concentration (plant growth regulators) such as IAA, GA, and kinetin are in agreement with previous findings reported [77].
Our results are also in line with earlier findings that has been stated by Spaepen et al., [78] and Mukhtar et al., [5]. Various bacterial strains have contributed to plant growth improvement and generated changes in Arabidopsis thaliana morphology. Further, the results dealing with the PGPR strain producing gibberellin and its impact on eggplant cultivars are in confirmation with previous reports showing that the treatment with gibberellins‐producing strain Promicromonospora sp. (SE188) boosted up gibberellins amount in plant tissue [79]. The authors findings [5, 80] are in accordance with our results indicating that the treatment of plants by bacterial strains caused a cytokinin production and improved shoot development and fruit generation, apart from boosting up plant capacity to counteract abiotic stress.
The treatment with PGPR was found to be efficient in mitigating the negative impacts of heat stress on plant growth parameters and its productivity [81]. Eggplant cultivar ‘Black Beauty’ exhibited higher susceptibility to heat stress in comparison to ‘Bonica’ when subjected to bacterial inoculation. Our finding agrees with previous results stated by Daim et al. [82] working on wheat and by Mukhtar et al. [5] working on tomato. In the present work, bacterial strains inoculation was efficient in mitigating the various impacts of high temperature on all studied growth parameters in the two eggplant cultivars.
Under heat stress condition, bacterial application ameliorated the two eggplant cultivars biomass in comparison to plants grown in absence of bacteria. The bacterial inoculation was also efficient in alleviation of heat stress in sorghum [81], in finger millet [67] and in tomato [5]. Earlier studies showed that under PGPR inoculation plant growth parameters were found to be improved due to the buildup of plant growth regulators in plant cells, thus enhancing cells growth and improving their efficacy in water and nutrient uptake [81]. It is well known that in tropical and temperate region, flowering phase and fruit formation are deeply affected by heat stress. Wahid et al. [83] reported that high temperature conditions can seriously affect flowers number. Additionally, under heat stress condition, plants exhibited lower fruit formation due to less carbohydrates and growth regulators production [84]. Our current results indicate that under normal and high temperature conditions, flowers and fruits number was boosted up by bacterial application in the two cultivars in comparison to those grown without bacterial inoculation. Under high temperature, ‘Bonica’ exhibited an increased Chlb amount in comparison to normal condition. This might be resulted from a more efficient photosynthetic rate generated by the application of PGPR, which was declined in plant subjected to high temperature in absence of bacterial inoculation in comparison to plants growing under normal environment without bacterial application [5, 85]. Besides, our findings are in agreement with some previously reported results [86] showing that the inoculation with Pseudomonas putida (P. putida) increased the shoot chlorophyll concentration in canola plant. Likewise, it has been stated [87, 88] that the abiotic stress alleviation was achieved through the Brevibacterium sp., (FAB3) inoculation which improved the chlorophyll concentration thus contributing to the plant productivity amelioration.
It is well known that plant antioxidant machinery is considered as feasible tool reflecting plant tolerance level under stressful environment. Plant antioxidant system including SOD, POD and CAT has a vital role in scavenging H2O2 under abiotic stress condition, thus leading to plant tolerance [89-91]. The bacterial application improved the antioxidant enzymes activity in both cultivars when subjected to high temperature and normal environment condition. Plant antioxidant defense system is crucial in mitigating oxidative stress engendered by high oxygen free radicals’ production [87, 92]. The bacterial inoculation boosted up SOD activity leading to a more scavenging capacity of reactive oxygen species abiotic stress. SOD and POD enhance the conversion of O2– to H2O2. Then CAT convert H2O2 into H2O and oxygen [59, 93]. The elimination of H2O2 by APX and CATis playing an important role in plant tolerance under abiotic stress condition [94]. Proline is an efficient plant osmo-protectant which deeply improve stress tolerance [95, 96]. Under heat stress, high level of proline buildup in leaves is considered as a protective system preserving cell membrane permeability and affecting tissues water exchange. Under high temperature and normal condition, the two cultivars showed higher proline accumulation in presence of bacterial inoculation in comparison with absence of bacterial application. Furthermore, it has been stated that under high temperature condition, P. putida inoculation led to a significant amelioration of proline buildup in wheat [97, 98]. Our current results agree with those of Afridi et al. [59], as they emphasized plant metabolites accumulation in bacterial inoculated sorghum under high temperature conditions. The present study investigated the putative potential process of high temperature tolerant PGP, ACC‐deaminase and EPS producing PGPR that lead to heat stress tolerance in two eggplant cultivars.
The application of PGPR strain might be considered as one of the most efficient biological approaches used by plant to mitigate heat stress and improve all the plant physiological and biochemical parameters. The use of high temperature resistant PGPR, ACC‐deaminase and EPS producing PGPR strain showed an effective amelioration in (1) plant physiology (SL and RL, FW and DW, and LSA) and (2) biochemistry (RWC, scavenging system) of two eggplant cultivars investigated under high stress. The studied bacterial strain is provided with an effective potential in plant growth traits promotion via IAA synthesis and phosphate solubilization, thus alleviating the heat stress adverse effects and improving plant productivity. The present study showed that a putative PGPR strain exhibited a key role in minimizing the adverse impacts of high temperature stress on plant development. Consequently, the production and use of PGPR compounds for counteracting heat stress negative effects seem to be crucial. The strategy dealing with adoption of PGPR becomes a must in order to preserve agriculture sustainability and healthy crops. The present study serves as useful basis supporting the potential integrative application of bacterial strain as bio-inoculant for suppression of heat stress damage in the field.
Acknowledgements
We thank Editage (http://www.editage.cn) for English language editing.
Author Contributions
SH conceived and designed research. SH conducted experiments. SH analyzed data. SH wrote the manuscript. KN and AS revised the manuscript. All authors read and approved the manuscript.
Conflict of Interest
The author declare that there is no conflict of interest.
![]() References
References
- Bita CE, Zenoni S, Vriezen WH, Mariani C, Pezzotti M, Gerats T. Temperature stress differentially modulates transcription in meiotic anthers of heat-tolerant and heat-sensitive tomato plants. BMC Genomics, (2011); 12: 384.
- Grover M, Ali SZ, Sandhya V, Rasul A, Venkateswarlu B. Role of microorganisms in adaptation of agriculture crops to abiotic stresses. World Journal of Microbiology and Biotechnology, (2011); 27: 1231–1240.
- Carbonell‐Bojollo R, Veroz‐Gonzalez O, Ordoñez‐Fernandez R, Moreno‐Garcia M, Basch G, Kassam A, Repullo‐Ruiberriz de Torres MA, Gonzalez‐Sanchez EJ. The effect of conservation agriculture and environmental factors on CO2 emissions in a rainfed crop rotation. Sustainability, (2019); 11: 3955.
- Zhang X, Rong X, Cai M, Meng Q. Collaborative optimization of emissions and abatement costs for air pollutants and greenhouse gases from the perspective of energy structure: An empirical analysis in Tianjin. Sustainability, (2019); 11:3872.
- Mukhtar T, Rehman SU, Smith D, Sultan T, Seleiman MF, Alsadon AA, Saad MA. Mitigation of heat stress in Solanum lycopersicum L. by ACC-deaminase and exopolysaccharide producing Bacillus cereus: effects on biochemical profiling. Sustainability, (2020); 12: 2159.
- Fahad S, Bajwa AA, Nazir U, Anjum SA, Farooq A, Zohaib A, Ihsan MZ. Crop production under drought and heat stress: Plant responses and management options. Frontiers in Plant Science, (2017); 8: 1147.
- Khan MA, Asaf S, Khan AL, Ullah I, Ali S, Kang SM, Lee IJ. Alleviation of salt stress response in soybean plants with the endophytic bacterial isolate Curtobacterium sp. SAK1. Annals of Microbiology, (2019); 69: 797-808.
- Barnabas B, Katalin J, Feher A. The effect of drought and heat stress on reproductive processes in cereals. Plant, Cell & Environment, (2008); 31: 11–38.
- Prasad PVV, Djanaguiraman M. Response of floret fertility and individual grain weight of cwheat to high temperature stress: Sensitive stages and thresholds for temperature and duration. Functional Plant Biology, (2014); 41: 1261–1269.
- Sita K, Sehgal A, Kumar J, Kumar S, Singh S, Siddique KH, Nayyar H. Identification of high‐ temperature tolerant lentil (Lens culinaris Medik.) genotypes through leaf and pollen traits. Frontiers in Plant Science, (2017); 8: 1–26.
- Devasirvatham V, Gaur PM, Mallikarjuna N, Tokachichu RN, Trethowan RM, Tan DKY. Effect of high temperature on the reproductive development of chickpea genotypes under controlled environments. Functional Plant Biology, (2012); (12):1009-1018.
- Fahad S, Hussain S, Saud S, Khan F, Hassan S, Nasim W, Huang J. Exogenously applied plant growth regulators affect heat-stressed rice pollens. Journal of Agronomy and Crop Science, (2016); 202:139–150.
- Fahad S, Hussain S, Saud S, Tanveer M, Bajwa AA, Hassan S, et al. A biochar application protects rice pollen from high-temperature stress. Plant Physiology and Biochemistry, (2015); 96: 281–287.
- Mattioli R, Marchese D, D’Angeli S, Altamura MM, Costantino P, Trovato M. Modulation of intracellular proline levels affects flowering time and inflorescence architecture in Arabidopsis. Plant Molecular Biology, (2008); 66: 277–288. 16.
- Siddique A, Kandpal G, Kumar PJ. Proline accumulation and its defensive role under diverse stress condition in pants: An overview. Journal of Pure and Applied Microbiology, (2018); 12: 1655–1659.
- Zouari M, Hassena AB, Trabelsi L, Rouina BB, Decou R, Labrousse P. “Exogenous proline–mediated abiotic stress tolerance in plants: possible mechanisms,” in Osmoprotectant–Mediated Abiotic Stress Tolerance in Plants, eds M. Hossain, V. Kumar, D. Burritt, M. Fujita, and P. Mäkelä (Cham: Springer), (2019); 99–122.
- Zulfiqar F, Akram NA, Ashraf M. Osmoprotection in plants under abiotic stresses: new insights into a classical phenomenon. Planta, (2020); 251:3.
- Sato S, Kamiyama M, Iwata T, Makita N, Furukawa H, Ikeda H. Moderate increase of mean daily temperature adversely affects fruit set of Lycopersicon esculentum by disrupting specific physiological processes in male reproductive development. Annals of Botany, (2006); 97: 731–738.
- Shrivastava P, Kumar R. Soil salinity: A serious environmental issue and plant growth promoting bacteria as one of the tools for its alleviation. Saudi Journal of Biological Sciences, (2015) 22: 123–131.
- Turan M, Yildirim E, Kitir N, Unek C, Nikerel E, Ozdemir BS, Güne SA, et al. Beneficial role of plant Growth-Promoting Bacteria in vegetable production under abiotic stress. In Microbial Strategies for Vegetable Production, Springer: Singapore, (2017); pp 151–166.
- Tiwari G, Duraivadivel P, Sharma S, Hariprasad P. 1-Aminocyclopropane-1- carboxylic acid deaminaseproducingbeneficialrhizobacteriaameliorate the biomasscharacters of Panicum maximum Jacq. by mitigating drought and salt stress. Scientific Reports, (2018); 8, 17513.
- Singh SB, Gowtham HG, Murali M, Hariprasad P, Lakshmeesha TR, Murthy KN, et al. Plant growth promoting ability of ACC deaminase producing rhizobacteria native to Sunflower (Helianthus annuus L.). Biocatalysis and Agricultural Biotechnology, (2019); 18: 101089.
- Gill SS, Gill R, Trivedi DK, Anjum NA, Sharma KK, Ansari MW, et al. Piriformospora indica: Potential and significance in plant stress tolerance. Frontiers in Microbiology, (2016); 7: 332.
- Glick BR. Bacteria with ACC‐ deaminase can promote plant growth and help to feed the world. Microbiological Research, (2014); 169: 30–39.
- Wanget SS, Tuteja N. Reactive oxygen species and antioxidant machinery in abiotic stress tolerance in crop plants. Plant Physiology and Biochemistry, (2010); 48: 909–930.
- Sarma RK, Saikia R. Alleviation of drought stress in mung bean by strain Pseudomonas aeruginosa GGRJ21. Plant and Soil, (2014); 377: 111–126.
- Tiwari S, Singh P, Tiwari R, Meena KK, Yandigeri M, Singh DP, et al. Salt‐tolerant rhizobacteria‐mediated induced tolerance in wheat (Triticum aestivum) and chemical diversity in rhizosphere enhance plant growth. Biology and Fertility of Soils, (2011); 47:907–916.
- Bashan Y, de-Bashan LE. Bacteria/plant growth-promotion (ed Hillel, D.), (Elsevier Oxford), (2005); 103–115.
- Compant S, Duffy B, Nowak J, Clément C, Barka EA. Use of plant growth promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Applied and Environmental Microbiology Journal, (2005); 71: 4951–4959.
- Warnita W, Ardi A, Zulfa Y. Effect of mulch and indigenous rhizobacteria isolate on growth and yield of potato (Solanum tuberosum L.). Asian Journal of Agriculture and Biology, (2019); 239–245
- Zahir ZA, Ghani U, Naveed M, Nadeem SM, Asghar HN. Comparative effectiveness of Pseudomonas and Serratia sp. containing ACC‐deaminase for improving growth and yield of wheat (Triticum aestivum L.) under salt‐stressed conditions. Archives of Microbiology, (2009);191: 415–424.
- Vacheron J, Desbrosses G, Bouffaud ML, Touraine B, Moënne-Loccoz Y, Muller D, et al. Plant growth-promoting rhizobacteria and root system functioning. Frontiers in Plant Science, (2013); 4: 356.
- Chapman MA. Introduction: The importance of Eggplant. In Compendium of Plant Genomes, (2019); Springer: Singapore, pp 1–10.
- Artyszak A, Gozdowski D. The Effect of Growth Activators and Plant Growth-Promoting Rhizobacteria (PGPR) on the Soil Properties, Root Yield, and Technological Quality of Sugar Beet. Agronomy, Agron, (2020); 10: 1262.
- Kour D, Rana KL, Yadav N, Yadav AN, Kumar A, Meena VS, et al. Rhizospheric Microbiomes: Biodiversity, Mechanisms of Plant Growth Promotion, and Biotechnological Applications for Sustainable Agriculture. In Plant Growth Promoting Rhizobacteria for Agricultural Sustainability; Springer: Singapore, (2019); pp. 19–65.
- Gupta S, Kaushal R, Gupta S. Plant Growth Promoting Rhizobacteria: Bioresouce for Enhanced Productivity of Solanaceous Vegetable Crops. Acta Scientific Agriculture, (2017); 1: 10–15.
- Yasmin S, Hafeez FY, Mirza MS, Rasul M, Arshad HM, Zubair M, et al. Biocontrol of bacterialleaf blight of riceandprofiling of secondarymetabolitesproducedbyrhizospheric Pseudomonas aeruginosa BRp3. Frontiers in Microbiology, (2017); 8: 1895
- Gupta RR, Singal R, Shanker A, Kuhad RC, Saxena RKA. A modified plate assay for screening phosphate solubilizing microorganisms. The Journal of General and Applied Microbiology, (1994); 40:255–260.
- Loper J, Schroth M. Influence of bacterial sources of indole‐3‐acetic acid on root elongation of sugar beet. Phytopathology, (1986); 76: 386–389.
- Ahmad F, Ahmad I, Khan MS. Indole acetic acid production by the indigenous isolates of Azotobacter and fluorescent Pseudomonas in the presence and absence of tryptophan. Turkish Journal of Biology, (2005); 29: 29–34.
- Schwyn B, Neilands JB. Universal chemical assay for the detection and determination of siderophores. Analytical Biochemistry, (1987); 160: 47–56.
- Castric PA. Hydrogen cyanide, a secondary metabolite of Pseudomonas aeruginosa. Canadian Journal of Microbiology, (1975); 21: 613–618.
- Glick BR, Karaturovic DM, Newell PC. A novel procedure for rapid isolation of plant growth promoting pseudomonas. Canadian Journal of Microbiology, (1995); 41: 533–536.
- Dworkin M, Foster J. Experiments with some microorganisms which utilize ethane and hydrogen. Journal of Bacteriology, (1958); 75: 592.
- Penrose DM, Glick BR. Methods for isolating and characterizing ACC‐deaminase containing plant growth‐promoting rhizobacteria. Physiologia Plantarum, (2003); 118: 10–15.
- Mu’minah B, Subair HF. Isolation and screening Bacterial Exopolysaccharide (EPS) from potato rhizosphere in highland and the potential as a producer Indole Acetic Acid (IAA). Procedia Food Science, (2015); 3:74–81.
- Kazempour MN. Biological control of Rhizoctonia solani, the causal agent of rice sheath blight by antagonistic bacteria in greenhouse and field conditions. Journal of Plant Pathology, (2004); 3, 88–96.
- Adebiyi CAB, Akinyaanju JA.Thermophillic amylase producers from the soil. Niger. Journal of Science and Technology, (1998); 11: 30–38.
- Namasivayam E, John RD, Mariappan K, Jiji A, Kumar M, Jayaraj RL. Production of extracellular pectinase by Bacillus cereus isolated from market solid waste. Journal of Bioanalysis & Biomedicine, (2011); 3: 70–75.
- Hussain A, Kamran MA, Javed MT, Hayat K, Farooq MA, Ali N, et al. Individual and combinatorial application of Kocuria rhizophila and citric acid on phytoextraction of multi‐metal contaminated soils by Glycine max L. Environmental and Experimental Botany, (2019); 159: 23–33.
- Yasmin H, Bano A. Isolation and characterization of phosphate solubilizing bacteria from rhizosphere soil of weeds of khewra salt range. Pakistan Journal of Botany, (2011); 43:1663–1668.
- Ma Y, Oliveira RS, Nai F, Rajkumar M, Luo Y, Rocha I, et al. The hyper accumulator Sedum plumbizincicola harbors metal‐ resistant endophytic bacteria that improve its phytoextraction capacity in multi‐metal contaminated soil. Journal of Environmental Management, (2015); 156: 62–69.
- Saber MAF, Abdelhafez AA, Hassan EA, Ramadan EM. Characterization of fluorescent pseudomonads isolates and their efficiency on the growth promotion of tomato plant. Annals of Agricultural Sciences, (2015); 60: 131–140.
- Hannachi S, Van Labeke MC. Salt stress affects germination, seedling growth andphysiological responses differentially in eggplant cultivars (Solanum Melongena L.). Scientia Horticulturae, (2018); 228: 56-65.
- Hoagland DR, Arnon DI. The water-culture method for growing for plants without soil. Circular. California Agricultural Experiment Station, (1950); 347: pp1-32.
- Barrs HD, Weatherley PE. A re-examination of the relativeturgiditytechniqueforestimating water defcits in leaves. Australian Journal of Biological Sciences, (1962); 15:413.
- Arnon DI. Copper enzymes in isolated chloroplasts. Polyphenoloxidase in Beta vulgaris. Plant Physiology, (1949); 24(1):1-15
- Lichtenthaler HK, Wellburn AR. Determinations of total carotenoids and chlorophylls a and b of leaf extracts in different solvents. Biochemical Society Transactions, (1983); 603: 591–592.
- Afridi MS, Amna S, Mahmood T, Salam A, Mukhtar T, Mehmood S, et al. Induction of tolerance to salinity in wheat genotypes by plant growth promoting endophytes: Involvement of ACC deaminase and antioxidant enzymes. Plant Physiology and Biochemistry, (2019); 139: 569–577.
- Bates LS, Waldren RP, Teare ID. Rapid determination of free proline for water-stress studies. Plant and Soil, (1973); 39: 205–207.
- Chance B, Maehly AC. Assay of catalases and peroxidases. Methods in Enzymology, (1955); 2: 764–775.
- Lu T, Yu H, Li Q, Chai L, Jiang W. Improving plant growth and alleviating photosynthetic inhibition and oxidative stress from low‐light stress withexogenous GR24 in tomato (Solanum Lycopersicum L.) seedlings. Frontiers in Plant Science, (2019); 10: 490.
- Aebi H. Catalase in vitro. Methods in Enzymology, (1984); 105: 121–126.
- Meena KK, Sorty AM, Bitla UM, Choudhary K, Gupta P, Pareek A, et al. Abiotic stress responses and microbe-mediated mitigation in plants: the omics strategies. Frontiers in Plant Science, (2017); 8: 172.
- Gowtham HG, BrijeshSingh M, Murali N, Prasad SM, Mohammed Aiyaz M, Amruthesh KN, et al. Induction of drought tolerance in tomato upon the application of ACC deaminase producing plant growth promoting rhizobacterium Bacillus subtilis Rhizo SF 48. Microbiological Research, (2020); 234: 126422.
- Turner TR, James EK, Poole PS. The plant microbiome. Genome Biol, (2013); 14 (6): 209.
- Chandra D, Srivastava R, Sharma AK. Influence of IAA and ACC deaminase producing fluorescent Pseudomonads in alleviating drought stress in Wheat (Triticum aestivum). Agricultural Research, (2018); 7: 290–299.
- Tiwari G, Duraivadivel P, Sharma S, Hariprasad P. 1-Aminocyclopropane-1- carboxylic acid deaminase producing beneficial rhizobacteria ameliorate the biomass characters of Panicum maximum Jacq. by mitigating drought and salt stress. Scientific Reports, (2018); 8: 17513.
- Niñerola A, Ferrer‐Rullan R, Vidal‐Suñé A. Climate Change Mitigation: Application of Management Production Philosophies for Energy Saving in Industrial Processes. Sustainability, (2020); 12: 717.
- Colantoni A, Monarca D, Marucci A, Cecchini M, Zambon I, Di Battista F, et al. Solar Radiation distribution inside a greenhouse prototypal with photovoltaic mobile plant and effects on flower growth. Sustainability, (2018); 10: 855.
- Spain A, Alm E. Implications of microbial heavy metal tolerance in the environment. Reviews in Undergraduate Research, (2003); 2:1–6.
- Ali S, Charles TC, Glick BR. Amelioration of high salinity stress damage by plant growth‐promoting bacterial endophytes that contain ACC‐ deaminase. Plant Physiology and Biochemistry, (2014); 80: 160–167.
- Pan YJ, Liu L, Lin YC, Zu YG, Li LP, Tang ZH. Ethylene antagonizes salt-induced growth retardation and cell death process via transcriptional controlling of ethylene-, BAG-and senescence-associated genes in Arabidopsis. Frontiers in Plant Science, (2016); 7: 696.
- Wang H, Lin J, Chang Y, Jiang CZ. Comparative Transcriptomic Analysis Reveals That Ethylene/H2O2-Mediated Hypersensitive Response and Programmed Cell Death Determine the Compatible Interaction of Sand Pear and Alternaria alternata. Frontiers in Plant Science, (2017); 8:195.
- Shahzad SM, Arif MS, Riaz M, Iqbal Z, Ashraf M. PGPR with varied ACC‐ deaminase activityinduced different growthandyield response in maize (ZeamaysL.) under fertilized conditions. European Journal of Soil Biology, (2013); 57: 27–34.
- Bhagat N, Raghav M, Dubey S, Bedi N. Bacterial Exopolysaccharides: Insight into Their Role in Plant Abiotic Stress Tolerance. Journal of Microbiolog and Biotechnology, (2021); (8):1045-1059.
- Spaepen S, Vanderleyden J, Remans R. Indole‐3‐acetic acid in microbial and microorganism‐plant signaling. FEMS Microbiology Reviews, (2007); 31: 425–448.
- Spaepen S, Bossuyt S, Engelen K, Marchal K, Vanderleyden J. Phenotypical and molecular responsesof A. thaliana roots as a result of inoculation with the auxin producing bacterium Azospirillum brasilense. New Phytologist, (2014); 201: 850–861.
- Kang SM, Khan AL, You YH, Kim JG, Kamran M, Lee IJ. Gibberellin production by newly isolated strain Leifsoniasoli SE134 anditspotentialto promote plant growth. Journal of Microbiolog and Biotechnology, (2014); 24:106–112.
- Azcon R, Barea J. Synthesis of auxins, gibberellins and cytokinins by Azotobacter vinelandii and Azotobacter bijerinkii related to effects produced on tomato plants. Plant and Soil, (1975); 43: 609–619.
- Ali SZ, Sandhya V, Grover M, Linga VR, Bandi V. Effect of inoculation with a thermo‐tolerant plant growth promoting Pseudomonas putida strain AKMP7 on growth of wheat (Triticumspp.) under heat stress. Journal of Plant Interactions, (2011); 6: 239–246.
- Daim IAA, Bejai S, Meijer J. Improved heat stress tolerance of wheat seedlings by bacterial seed treatment. Plant and Soil, (2014); 379: 337–350.
- Wahid A, Gelani S, Ashraf M, Foolad M. Heat tolerance in plants: An overview. Environmental and Experimental Botany, (2007); 61: 199–223.
- Kinet JM, Peet MM. Tomato. In The Physiology of Vegetable Crops; Wien, H.C., Ed.; CAB International: Wallingford, UK; (1997); pp. 207–258.
- Rehman SU, Afzal W, Anjum T, Choudhry HJ, Ahmad SR, Aslam M. Screening and evaluation of indigenous halo‐tolerant microbes forsalt stress alleviation in celery (Apium graveolens). Soil & Environment, (2019); 38:42–47.
- Ali J, Mahmood T, Hayat K, Afridi MS, Ali F, Chaudhary HJ. Phytoextraction of Cr bymaize (Zea maysL.). The role of plant growth promoting endophyte and citric acid under polluted soil. Archives of Environmental Protection, (2018); 44: 73–82.
- Ansari FA, Ahmad I. Plant growth promoting attributesandalleviation of salinity stress tow heat by biofilm forming Brevibacterium sp. FAB3 isolated from rhizospheric soil. Saudi J Biol Sci, (2018); in press.
- Ahamd M, Hussain A, Akhtar FUZM, Zafar‐Ul‐Hye M, Iqbal Z, et al. Effectiveness of multi‐strainbiofertilizer in combinationwithorganic sources for improving the productivity of Chickpea in Drought Ecology. Asian Journal of Agriculture and Biology, (2017); 5: 228‐237.
- Zandalinas SI, Mittler R, Balfagón D, Arbona V, Gómez-Cadenas A. Plant adaptations to the combination of drought and high temperatures. Physiologia Plantarum, (2017); 162:2–12.
- Zhou R, Yu X, Kjær KH, Rosenqvist E, Ottosen C-O, Wu Z. Screening andvalidation of tomatogenotypesunder heat stress using Fv/Fm toreveal the physiological mechanism of heat tolerance. Environmental and Experimental Botany, (2015); 118:1–11.
- Xu HC, Tie C, Wang Z, He MR. Physiological basis for the differences of productive capacity among tillers in winter wheat. Journal of Integrative Agriculture, (2015); 14: 1958–1970.
- Raja V, Qadir SU, Alyemeni MN, Ahmad P. Impact of drought and heat stress individually and in combination on physio-biochemical parameters, antioxidant responses, and gene expression in Solanumly copersicum.3 Biotech, (2020); 10(5):1-18.
- Carvalho LSC, Vidigal PC, Amancio S. Oxidative stress homeostasis in grapevine (VitisviniferaL.). Frontiers in Environmental Science, (2015); 3:20.
- Koussevitzky S. Ascorbate peroxidase 1 plays a key role in the response of Arabidopsis thaliana to stress combination. Journal of Biological Chemistry, (2008); 283:34197–34203.
- Akram NA, Iqbal M, Muhammad A, Ashraf M, Al-Qurainy F, Shafiq S. Aminolevulinic acid andnitric oxide regulate oxidative defense and secondary metabolisms in canola (BrassicanapusL.) under drought stress. Protoplasma, (2017); 255:163–174.
- Alzahrani Y, Kuşvuran A, Alharby HF, Kuşvuran S, Rady MM. The defensive role of silicon in wheat against stress conditions induced by drought, salinity or cadmium. Ecotoxicol Environ Saf, (2018); 154:187–196.
- Ashraf M, Foolad MR. Roles of glycine betaine and proline in improving plant abiotic stress resistance. Environmental and Experimental Botany, (2007); 59: 206–216.
- Ikram R, Ali B. Co‐inoculation of auxinproducing PGPR andrhizobiaenhancedgrowth of Vignamungo (L.) under cadmium stress. Asian Journal of Agriculture and Biology, (2018); 6: 46‐54.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0