

Full Length Research Article
Protective immune response of recombinant Fiber-2 protein as subunit vaccine against Fowl-adenovirus-4 infection in Pakistan
Faiza Ather1, Muhammad Ashir Zia2*, Muhammad Salahuddin Shah1, Mudasser Habib1
Adv. life sci., vol. 11, no. 2, pp. 315-321, May 2024
*– Corresponding Author: Muhammad Ashir Zia (ashirzia68@gmail.com)
Authors' Affiliations
2. Department of Biological Sciences, Virtual University of Pakistan, Lahore, Punjab – Pakistan
[Date Received: 13/10/2022; Date Revised: 09/01/2024; Date Available Online: 18/04/2024]
Editorial Note: You are viewing the latest version of this manuscript having major changes in order of authors.
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Hydropericardium Syndrome (HPS) has emerged as a major viral disease of poultry industry causing huge economic losses in the recent past years. Its etiological agent, Fowl adenovirus-4 (FAdV-4) has been reported in the various countries including Pakistan. Various strategies including inactivated vaccines, are being devised to control the disease since its appearance. Subunit vaccines based on viral structural proteins have demonstrated more promising protection against FAdV-4 infection than commercial inactivated vaccines. Among different viral structural proteins, Fiber protein (mainly Fiber-2) has been as a suitable candidate for developing the recombinant subunit vaccine against HPS.
Methods: Considering the importance of the Fiber-2 gene, the pET28a expression vector was utilized to clone its open reading frame, which was subsequently expressed as an oligo-histidine tagged fusion protein in BL21 cells of Escherichia coli via the IPTG induction method. The expressed recombinant fiber-2 protein was purified using nickel (Ni2+) affinity chromatography and used as a subunit vaccine in broiler birds following challenge with pathogenic FAdV-4 isolate. The immunological response was evaluated using ELISA.
Results: The gene for the Fiber-2 protein was effectively cloned and expressed as a soluble 60 kDa protein, as detected by SDS-PAGE and western blot analysis. The protective efficacy of subunit vaccine was assessed by ELISA which showed the highest protection (80%) against the virus challenge than that of commercial inactivated vaccine (70%).
Conclusion: The recombinant fiber-2 protein was determined to be a good option for a recombinant subunit vaccination to control HPS.
Keywords: Poultry; Fowl-adenovirus-4; Subunit vaccine
Introduction![]()
Hydropericardium syndrome (HPS) is a poultry infectious disease that predominantly impacts broiler chickens within the age range of three to six weeks. The disease begins quickly and shows no significant clinical signs and symptoms [1]. A disease was detected in broilers in Pakistan between 1987 and 1988 and was designated Angara disease (after the location) Angara Goth, which is situated near Karachi, Pakistan [2]. After that, the disease erupted and spread all over the world. Since then, various strategies have been devised to control the disease. Recent years in Pakistan have witnessed FAdV outbreaks that have resulted in substantial losses for poultry flocks despite the administration of vaccinations as prescribed [3]. FAdVs have been classified into five molecular groups, such as FAdV-A-E. They are further categorized into twelve serotypes, such as FAdV-1 to 8a and 8b to 11, based on restriction enzymes, sequencing of particular genes, and serum cross neutralization tests [4,5]. The cause of HPS is Fowl adenovirus-4 (FAdV-4) of group C, which is a non-enveloped, icosahedral-shaped virion of 70-90 nm in diameter. FAdV-4 belongs to the family Adenoviridae and the genus Adenovirus [6-8].
The virus is about 43-46 kbp of DNA in size. Many viral proteins (structural and non-structural) are encoded by viral genome [9-11]. The viral capsid consists of structural proteins, which include Hexon, Penton base and two fibers having different lengths and various non-structural proteins [12]. Long fiber and short fiber are referred to as Fiber-1 and Fiber-2, respectively. Fiber-2 plays a key role in facilitating the attachment of the viral particles and the penetration of nuclear material into host cells and therefore it has a crucial influence on the pathogenicity of the virus [13]. The fibrous protein was found to consist of three regions namely tail, shaft and knob [14]. The hemagglutinating properties of adenoviruses depend on the sequence of the fiber knob [15]. The fiber knob is also involved in the synthesis and encapsidation of the fiber protein [16]. It also showed that different receptors were present on each fiber, suggesting that essentially both fibers are needed [17,18]. Fiber-2 protein consists of 479 amino acids and has epitopes against virulence determinants, cell surface binding receptors and tissue tropism [19]. Fiber 2 protein is one of the main structural proteins that is crucial for mediating FAdV-4 infection [20]. Fiber 2 stimulates fast CD4+ T-cell proliferation and offers greater immunity against avian adenovirus infection over the inactivated vaccine [21]. A recombinant fiber-2 protein expressed using the baculovirus system, has been shown to confer protection in susceptible birds [22].
Considering the importance of Fiber-2 proteins and limitations of baculovirus expression such as extensive manipulation to design plasmid, transfection process and use of cell culture [23], E. coli was used to generate the fiber-2 protein, which was then purified using metal affinity chromatography. The recombinant fiber-2 protein was used as subunit vaccine and its efficacy was analyzed by clinical observation and protection exerted following the virus challenge.
Methods![]()
Construction of Fiber-2 Expression vector
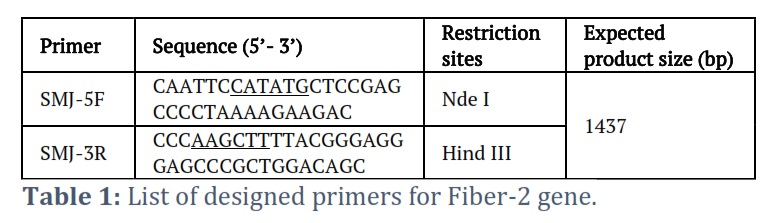
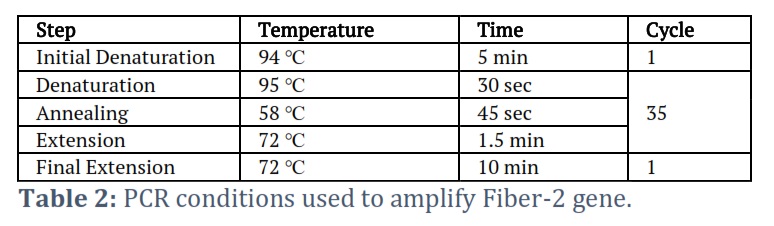
The DNA of Fowl Adenovirus-4 DNA (Group C) was extracted from the viral isolate (GenBank accession Number: HE649966.1) using Phenol chloroform extraction method [24]. Viral DNA was isolated from the liver of an infected chicken. To accomplish this, an equivalent volume (200µl) of lysis buffer containing 25µM Tris-HCl, 100µM NaCl, 0.5% SDS, and 25µg/ml Proteinase K was introduced into a 1.5 ml Eppendorf tube containing 200µl of semi-purified viral suspension (obtained by grinding infected liver in PBS and collecting supernatant by spinning the mixture). For an hour, the tube was kept at 37℃. Once the sample was at room temperature, 400 µl of Phenol-chloroform was added and mixed well. The mixture was then spun at 13000 rpm for 5 minutes. The supernatant was put into a new Eppendorf tube, and the NaCl content was set to 0.2M. To do this, the supernatant's overall volume was calculated, and a 1/10th volume of 2M NaCl solution was added. Two volumes of cooled ethanol were used to precipitate DNA for thirty minutes at -20°C. Centrifugation was used to recover the DNA for ten minutes at 13,000 rpm. The supernatant was disposed of while the DNA pellet was dried and dissolved in sterile water. The concentration of DNA was ascertained utilizing a Nanodrop spectrophotometer. The isolated DNA was subjected to PCR to amplify the Fiber-2 gene using the gene-specific primers incorporated with the Nde I (Fermentas, USA Cat # ER0581) and Hind III restriction sites (Fermentas, USA Cat # FD0505). The primers and PCR conditions used in the study are shown below (table 1).
The amplified DNA product and pET28a expression vector (Novagen Cat # 69864-3) were cleaved at the restriction sites. Buffers and enzymes from New England Biolab were employed in this process. A microfuge tube with 1 µg of DNA was filled with a 1X buffer (NEB buffer 10X, 5µL), appropriate for both restriction enzymes. 10 units (1 µg DNA) of each restriction enzyme was added and to prepare the reaction, 50 µL of nuclease-free sterile water was added. The enzymes were allowed to react in an incubation of 37℃ for 1-2 hours. Restricted insert and vector DNA fragments were mixed with 10 X ligation buffer and T4 DNA ligase and volume was made up to 20 µL with nuclease-free sterile water. The ligation process was maintained at 16℃ overnight. The enzymes and buffers of Fermentas were used for the ligation reaction. The resultant ligated product was transformed into E. coli competent cells (DH5α) as described in Molecular Cloning: A Laboratory Manual [25]. To perform this experiment, 200 µl of competent cells were frozen and thawed on ice for 10 minutes. The ligated products (0.2 – 0.5 µg DNA) were gently pipetted into the competent cells and mixed carefully. The vial was kept on ice for half an hour. A 30-second heat shock was administered in a water bath at 42ºC, followed by keeping it on ice for 2 minutes. Next, 500 µl of LB broth was added to the vial, and it was incubated for an hour at 37ºC while being shaken at 250 rpm. The mixture was spread to LB agar plates that had been pre-heated and contained a selective antibiotic (50µg/ml of Kanamycin). After 16 hours at 37℃, the plates were examined. The colonies that appeared on the plates were confirmed by colony PCR using primers specific to the gene. BigDye was used to analyze DNA sequences using an ABI 3100 capillary sequencer. DNA sequencing was done at Department of Biotechnology and cell biology, University of Connecticut, Storrs, CT. USA in order to deduce and analyze the amino acid sequence so that the correct ORFs and BLAST with already published sequences could be performed.
Expression and purification of Fiber-2 protein
The E. coli BL21 (DE3) cells were transformed with the recombinant Fiber-2 expression vector. A single transformed colony was inoculated into LB medium with kanamycin (50 µg/mL) and incubated at 37℃ overnight. Next day, the grown culture was diluted at 1:50 ratio into fresh LB media and incubated for 3 hours at 37℃ until OD600 was around 0.5. The new culture was then induced with 1 mM isopropyl-D-thiogalactoside (IPTG) and incubated for 4 hours at 37 ℃. Cells were extracted by centrifugation at 5000 rpm for 15 minutes at 4℃. The cell pellet was then dissolved into PBS.
The PBS dissolved-cells were broken down by sonication with 30 cycles (10 seconds ON and 20 seconds OFF) at 70% amplitude by using Qsonicator 500, Qsonica, CT, USA and then centrifuged at 13000 rpm for 20 minutes at 4℃ to separate supernatant and pellet fractions. The supernatant and pellet fractions were visualized using SDS-PAGE to analyze protein expression followed by Coomassie blue staining. As analyzed by SDS-PAGE, protein was expressed in supernatant as well as pellet. The expressed protein was purified using Nickle affinity chromatography under native/denaturing conditions. The aforementioned protein supernatant was put onto the Ni-NTA column, which had been pre-conditioned using binding buffer (Novagen). 5.25 mL of 1 X Ni NTA wash buffer (Novagen) was used to wash the column, and flowthrough was collected. Ultimately, 5 ml of 1 X Ni-NTA elute buffer (Novagen) was used to elute the bound protein. The eluted protein (5 aliquots of 1 ml volume) was then quantified using Nanodrop spectrophotometer. The fractions collected from the purification process were analyzed by SDS-PAGE.
Immunoblotting
Following separation on SDS-PAGE, the recombinant Fiber-2 protein was transferred onto the PVDF membrane using semi-dry blotter (Trans blot SD – Bio-Rad system). After that, the membrane was blocked for an hour at room temperature using 3% BSA in TBST. After washing twice with IX TBST, the membrane was incubated with mouse anti-histidine antibody prepared in blocking buffer for 1 hour. Three TBST washes and one TBS wash were performed on the membrane. The membrane was then treated with anti-mouse IgG (Sigma Aldrich, Cat # A3562) conjugated with alkaline phosphatase for 1 hr and was again washed with TBST 3-4 times. After washing BCIP/NBT substrate (bromo-chloro-indolyl-phosphate nitro blue tetrazolium) was mixed to identify the immune complexes.
Immunization and challenge studies
A group of thirty broiler chicks (one-day old) were bought from local hatchery (Al-Noor Chicks, Faisalabad) and raised under the protocols of the Institutional Animal Care and Use Committee (IACUC) of Animal Sciences Division, NIAB. They were split up into three groups, A, B, and C, each consisting of ten chickens (n = 10). On the fourteenth day, fiber-2 protein (25µg/mL dissolved in PBS and emulsified in Freund's complete adjuvant (FCA)) was subcutaneously administered into Group A birds. Group B broilers were injected subcutaneously with inactivated vaccine (Angara NIAB) [26] and Group C birds were injected with PBS as a control. Blood samples were collected from the birds of each group as day 0, 7, 14 and 21 after immunization to determine the antibody titer. On day 21 after vaccination, the birds were challenged with semi-purified virus (105.5 units of LD50/ml) [27] and observed clinically on daily basis for continuous 7 days. On day 21 after vaccination, the birds were challenged with semi-purified virus (105.5 units of LD50/ml) and observed clinically on daily basis for continuous 7 days. Clinical signs were observed on daily basis following challenge and assessed as follows: Normal (0), Mild depression (1), severe depression (2), Prostration/head down (3) and death (4) as described previously [28]. Mortality rates were calculated in all experimental groups and postmortem was performed to observe the postmortem lesions of HPS in each experimental group. Protection was confirmed by the absence of clinical signs and death in the vaccinated birds.
Enzyme-linked immunosorbent assay (ELISA)
The recombinant fiber-2 protein antigen was applied to ELISA plates at a concentration of 10 µg/ml, and the plates were then blocked using a blocking buffer (5% BSA). The plates were washed and incubated at 37 ℃ for one hour with the chicken sera of the experiment at a dilution of 1:100. After washing, the plate was incubated at 37 ℃ for one hour with AP conjugated anti-chicken IgG secondary antibodies. Following another wash of the plates, the p-nitrophenyl phosphate substrate was added to each well, and the plates were then left to incubate for 30 minutes in the dark at room temperature. After incubation, 3 M NaOH was used to terminate the reaction, and an ELISA reader was used to measure the colour intensity at 450 nm wavelength.
Results![]()
Analysis of fiber-2 gene sequence
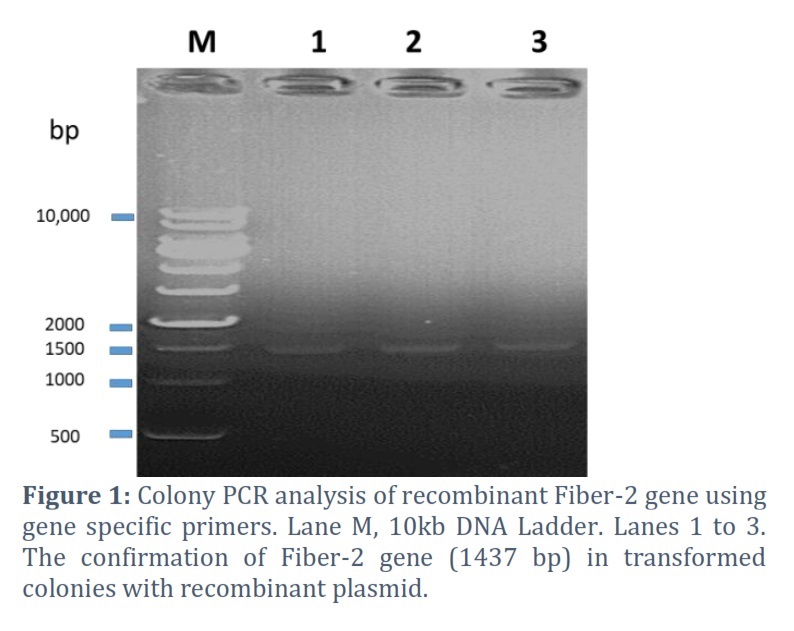
Gene code for Fiber-2 protein was amplified by PCR revealed the desired 1437 bp DNA product, which was then purified and cloned into pET28a expression vector. The insert's presence in the construct was further verified using colony PCR (Fig 1). DNA sequencing showed the accurate ORF of the fiber-2 gene in the construct. The deduced amino acid sequence revealed that Fiber-2 protein has 478 amino acid residues. Molecular studies also disclosed that fiber-2 gene has 99% similarity with FAdV-4 isolates originated from Indian regions.
Fiber-2 protein expression and purification
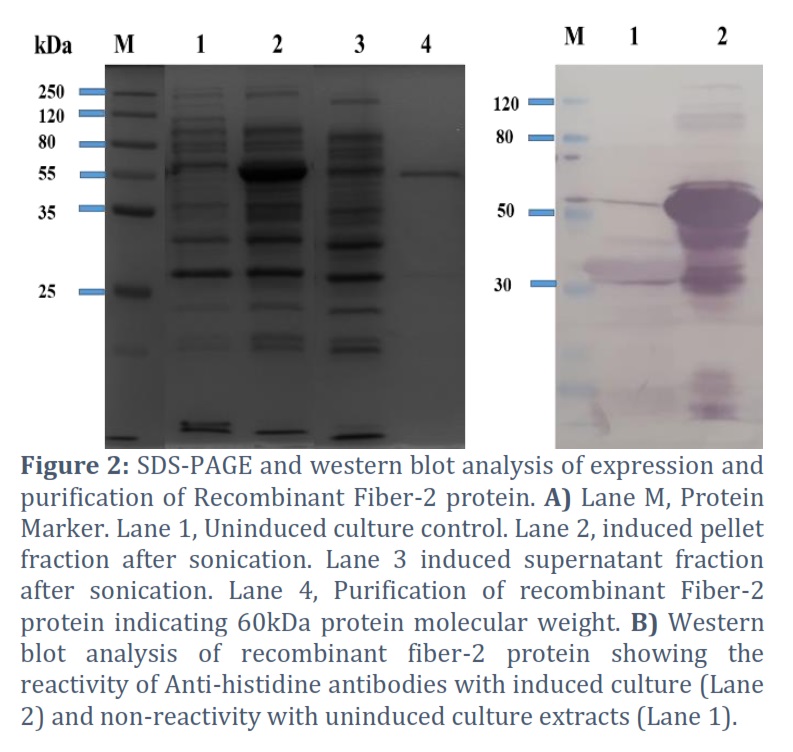
The recombinant Fiber-2 protein was successfully expressed in E. coli, yielding the predicted 60kDa protein on SDS-PAGE and Western blot (Fig 2A and B). Expression studies revealed that the recombinant Fiber-2 protein was expressed in supernatant as well as pellet fractions. The highest protein was found in inclusion bodies as insoluble proteins. It was also observed that by altering the factors like IPTG concentrations, temperature change and incubation time had no effect on the solubility of the recombinant fiber-2 protein (data not shown). The recombinant protein was purified using nickel affinity chromatography, revealing the presence of His-tagged recombinant fiber-2 in the eluted fractions with a molecular weight of 60 kDa. The obtained purified protein was quantified by nanodrop spectrophotometer which showed 1.5 mg/L of Fiber-2 protein from one-liter bacterial culture.
Immunogenicity of recombinant fiber-2 protein as subunit vaccine
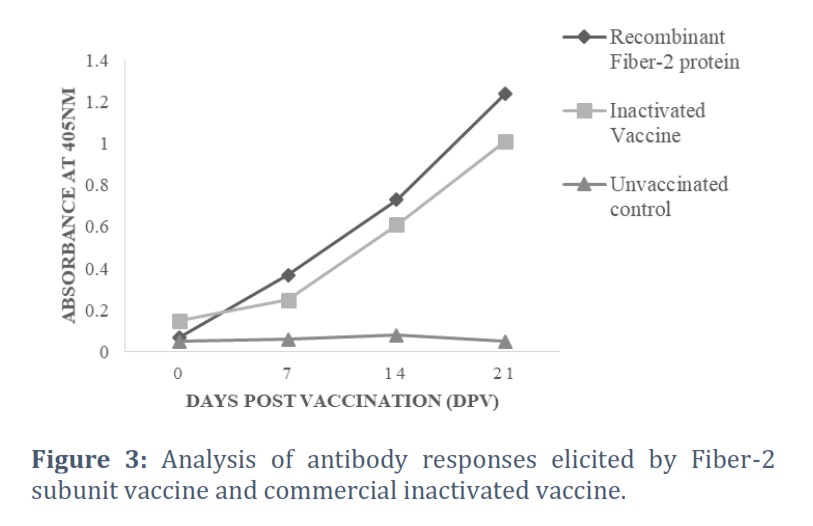
The antibody titers against Fiber-2 subunit vaccine and commercial inactivated vaccine were analyzed and compared at 0, 7, 14 and 21 days post vaccination (Fig. 3). It was found that production of antibodies was faster in the birds immunized with fiber-2 protein than those vaccinated with commercial vaccine (Fig 3). Moreover, high antibody titers were observed in birds vaccinated with Fiber-2 as compared with the birds immunized with commercial vaccine. This indicates that the fiber-2 protein produces more efficient immune response than commercial inactivated vaccine.
Protective efficacy of recombinant Fiber-2 protein
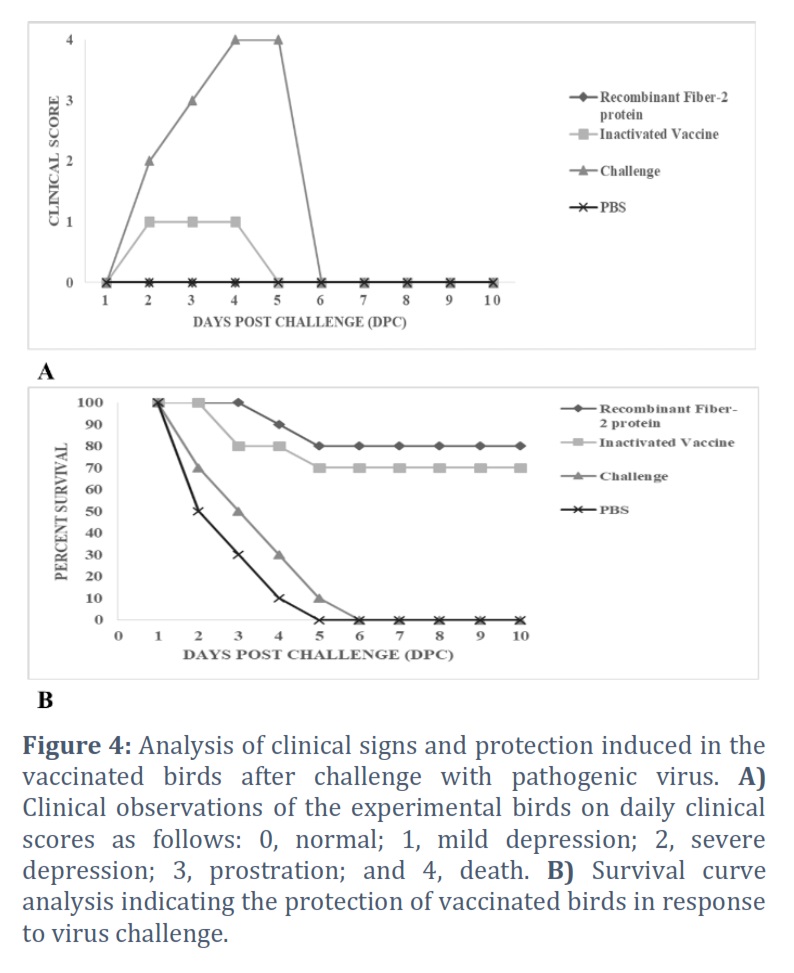
At 21 days after vaccination, the birds were challenged with pathogenic FAdV-4 virus strain and observed daily for ten days. In order to evaluate the protective efficacies of both vaccinated groups, a protection test was conducted. All the birds in the unvaccinated (PBS) and challenged group showed definite clinical signs such as severe depression, lameness, unable to walk or stand, and inappetence at two days post challenge. Mortality in the control group started at 3 dpc and reached 100 % at 5dpc (Fig 4A). Birds vaccinated with fiber-2 showed 80 % protection against the virus challenge than 70% protection induced by inactivated vaccine. The birds vaccinated with commercial vaccine also showed slight depression following virus challenge (Fig 4B).
Figures & Tables
In recent times, outbreaks of HPS have caused significantly greater bird mortality than earlier, milder outbreaks, thereby inflicting substantial economic losses on the poultry industry. Clinical investigations of samples collected from various poultry farms in Pakistan during the surveillance programme (2011-2016) revealed the presence of a mixed infection, which encompassed HPS as well [29]. Therefore, to minimize the mortality rates and infection status, efficient vaccination is very much needed. Many countries have attempted to develop an inactivated/autogenous vaccine and a live attenuated vaccine to control the HPS [30]. However, the incomplete inactivation of the virus and possible tumorigenicity can lead to further spread of the disease and other pathogens [28,31]. Routine immunization of chickens against HPS with inactivated FAdV-4 is observed to be effective in Pakistan, where it appears to reduce the severity of the disease. These inactivated vaccines have variable efficiencies [30]. Recent years in Pakistan have witnessed FAdV outbreaks that have resulted in substantial losses for poultry flocks despite the administration of vaccinations as prescribed [3]. Thus, subunit vaccines on the other hand, are more promising avoiding the risks associated with the traditional vaccines [32]. Fiber proteins play crucial roles in virus infectivity and pathogenicity. Being an important structural protein epitope, fiber 2 protein is considered as suitable candidate to develop safe and effective subunit vaccine [22]. E. coli [28], baculovirus [22] and yeast [33] expression hosts have been utilized to develop fiber 2 protein which confer protection against virus challenge. E. coli is preferred host due to its genetics, low cost, fast growth and high production yields of recombinant proteins [34]. As a result, we compared the immunization and protective efficacy of the fiber-2 protein derived from the Pakistani FAdV-4 isolate with that of the commercially available inactivated FAdV-4 vaccine in E. coli. Previous studies showed that the majority of FAdV-4's recombinant fiber 2 proteins produced as inclusion bodies in E. coli [28,32] which required dialysis and refolding prior to the purification. We used the supernatant fraction after the recombinant Fiber-2 protein was generated in E. coli, however our data also indicated that the protein was substantially expressed as inclusion bodies. In order to improve the solubility, different parameters such as IPTG concentrations, temperature and post induction time were altered solubility was not improved. Although the yield of recombinant protein after purification was low but enough to be used as subunit vaccine. Prior research also established the lowest amount of recombinant fiber-2 protein required per bird to offer defense against the virus challenge. It was found that the birds administered with 2.5 µg of recombinant fiber-2 or above provided protection against the virus challenge and produced high levels of antibodies [28]. In our study, we used 25 µg of recombinant fiber-2 protein which showed 80% protection against the virus challenge as compared with 70% protection provided by inactivated vaccine. The findings of our investigation are not entirely consistent with a recent study that demonstrated chicken immunization with fiber-2 protein provided 100% protection against a highly pathogenic serotype 4 of the fowl adenovirus (FAdV) [21]. Our findings demonstrated that birds immunized with fiber-2 protein exhibited higher antibody titers and faster antibody generation than birds immunized with inactivated FAdV-4 vaccine, suggesting that fiber-2 protein is more effective at eliciting protective immunity. These findings supported earlier findings that immunizing broilers with pure recombinant penton base protein produced a greater immunological response than using a traditional inactivated FAdV-4 vaccine [35]. Therefore, it is suggested that further research is necessary to improve protection.
In conclusion, our findings shown that immunization with recombinant fiber-2 protein (25 µg) can offer 80% protection against virulent FAdV-4 strain challenge. It induces the higher antibody levels than inactivated FAdV-4 vaccine. These results may contribute to the development of fiber-2 protein-based subunit vaccines that are both safe and effective in controlling HPS in Pakistan.
Acknowledgement
The study was supported by IAEA RAS 5085 project to MSS.
Conflict of Interest
The authors declare that there is no conflict of interest.
All authors contributed equally to this study.
![]() References
References
- Anjum A, Sabri M, Iqbal Z. Hydropericarditis syndrome in broiler chickens in Pakistan. The Veterinary Record, (1989); 124(10): 247-248.
- Jaffery M. A treatise on Angara disease (hydropericardium-pulmonary oedema-hepatonephritis syndrome). Journal of the Pakistan Medical Association, (1988); 34(1): 1-33.
- Wajid A, Basharat A, Shahid MA, Muntaha ST, Basit A, et al. Molecular characterization and phylogenetic analysis of fowl adenoviruses isolated from commercial poultry flocks in Pakistan during 2014-15. Pakistan Journal of Zoology, (2018); 50(5): 1863-1873.
- Zsák L, Kisary J. Grouping of fowl adenoviruses based upon the restriction patterns of DNA generated by BamHI and HindIII. Intervirology, (1984); 22(2): 110-114.
- Hess M. Detection and differentiation of avian adenoviruses: a review. Avian Pathology, (2000); 29(3): 195-206.
- Hess M, Raue R, Prusas C. Epidemiological studies on fowl adenoviruses isolated from cases of infectious hydropericardium. Avian Pathology, (1999); 28(5): 433-439.
- Anjum A. Experimental transmission of hydropericardium syndrome and protection against it in commercial broiler chickens. Avian Pathology, (1990); 19(4): 655-660.
- Mansoor MK, Hussain I, Arshad M, Muhammad G, Hussain MH, et al. Molecular characterization of fowl adenovirus serotype 4 (FAV-4) isolate associated with fowl hydropericardium-hepatitis syndrome in Pakistan. Pakistan Journal of Zoology, (2009); 41(4): 269-276.
- Griffin BD, Nagy É. Coding potential and transcript analysis of fowl adenovirus 4: insight into upstream ORFs as common sequence features in adenoviral transcripts. Journal of general virology, (2011); 92(6): 1260-1272.
- Davison AJ, Benkő M, Harrach B. Genetic content and evolution of adenoviruses. Journal of general virology, (2003); 84(11): 2895-2908.
- Marek A, Nolte V, Schachner A, Berger E, Schlötterer C, et al. Two fiber genes of nearly equal lengths are a common and distinctive feature of Fowl adenovirus C members. Veterinary Microbiology, (2012); 156(3-4): 411-417.
- Shah M, Ashraf A, Khan M, Rahman M, Habib M, et al. Fowl adenovirus: history, emergence, biology and development of a vaccine against hydropericardium syndrome. Archives of virology, (2017); 162(7): 1833-1843.
- Yin D, He L, Zhu E, Fang T, Yue J, et al. A fowl adenovirus serotype 4 (FAdV-4) Fiber2 subunit vaccine candidate provides complete protection against challenge with virulent FAdV-4 strain in chickens. Veterinary microbiology, (2021); 263109250.
- Zubieta C, Schoehn G, Chroboczek J, Cusack S. The structure of the human adenovirus 2 penton. Molecular cell, (2005); 17(1): 121-135.
- Pehler-Harrington K, Khanna M, Waters CR, Henrickson KJ. Rapid detection and identification of human adenovirus species by adenoplex, a multiplex PCR-enzyme hybridization assay. Journal of clinical microbiology, (2004); 42(9): 4072-4076.
- Henning P, Lundgren E, Carlsson M, Frykholm K, Johannisson J, et al. Adenovirus type 5 fiber knob domain has a critical role in fiber protein synthesis and encapsidation. Journal of general virology, (2006); 87(11): 3151-3160.
- Hess M, Cuzange A, Ruigrok RW, Chroboczek J, Jacrot B. The avian adenovirus penton: two fibers and one base. (1995); 252(4): 379-385.
- Tan PK, Michou A-I, Bergelson JM, Cotten M. Defining CAR as a cellular receptor for the avian adenovirus CELO using a genetic analysis of the two viral fibre proteins. Journal of general virology, (2001); 82(6): 1465-1472.
- Mase M, Nakamura K, Minami F. Fowl adenoviruses isolated from chickens with inclusion body hepatitis in Japan, 2009–2010. Journal of Veterinary Medical Science, (2012); 74(8): 1087-1089.
- Li L, Wang J, Chen P, Zhang S, Sun J, et al. Pathogenicity and molecular characterization of a fowl adenovirus 4 isolated from chicken associated with IBH and HPS in China. BMC veterinary research, (2018); 14(1): 1-8.
- Chen L, Yin L, Zhou Q, Li Q, Luo Y, et al. Immunogenicity and protective efficacy of recombinant fiber-2 protein in protecting SPF chickens against fowl adenovirus 4. Vaccine, (2018); 36(9): 1203-1208.
- Schachner A, Marek A, Jaskulska B, Bilic I, Hess M. Recombinant FAdV-4 fiber-2 protein protects chickens against hepatitis–hydropericardium syndrome (HHS). Vaccine, (2014); 32(9): 1086-1092.
- Yu X, Hua J, Zhang J, Ma LJAV. Expression of recombinant proteins in insect cells by their direct infection with Escherichia coli transformed with baculovirus bacmids. (2014); 5861-68.
- Ganesh K, Suryanarayana V, Raghavan R, Gowda S. Nucleotide sequence of L1 and part of P1 of hexon gene of fowl adenovirus associated with hydropericardium hepatitis syndrome differs with the corresponding region of other fowl adenoviruses. Veterinary microbiology, (2001); 78(1): 1-11.
- Sambrook J FE, Maniatis T. Molecular cloning: a laboratory Manual. Chapter: Book Name. 1989 of publication; Cold Spring Harbor Laboratory Press,Cold Spring Harbor, N.Y.
- Tufail S, Shah MA, Zafar M, Asif TA, Shehzad A, et al. Identification of potent epitopes on hexon capsid protein and their evaluation as vaccine candidates against infections caused by members of Adenoviridae family. Vaccine, (2021); 39(27): 3560-3564.
- Shah M, Ashraf A, Khan M, Rahman M, Habib M, et al. Molecular cloning, expression and characterization of 100K gene of fowl adenovirus-4 for prevention and control of hydropericardium syndrome. Biologicals, (2016); 44(1): 19-23.
- Ruan S, Zhao J, Yin X, He Z, Zhang G. A subunit vaccine based on fiber-2 protein provides full protection against fowl adenovirus serotype 4 and induces quicker and stronger immune responses than an inactivated oil-emulsion vaccine. Infection, Genetics and Evolution, (2018); 61: 145-150.
- Wajid A, Dimitrov KM, Wasim M, Rehmani SF, Basharat A, et al. Repeated isolation of virulent Newcastle disease viruses in poultry and captive non-poultry avian species in Pakistan from 2011 to 2016. Preventive veterinary medicine, (2017); 1421-6.
- Schachner A, Matos M, Grafl B, Hess M. Fowl adenovirus-induced diseases and strategies for their control–a review on the current global situation. Avian Pathology, (2018); 47(2): 111-126.
- Su Q, Hou L, Gao Y, Liu X, Cui Z, et al. Research Note: Molecular relationship of the fowl adenovirus serotype 4 isolated from the contaminated live vaccine and wild strains isolated in China, 2013-2018. Poultry Science, (2020); 99(12): 6643-6646.
- Wang X, Tang Q, Chu Z, Wang P, Luo C, et al. Immune protection efficacy of FAdV-4 surface proteins fiber-1, fiber-2, hexon and penton base. Virus research, (2018); 2451-6.
- Lai VD, Min K, Lai HTL, Mo J. Epidemiology of fowl adenovirus (FAdV) infections in South Korean chickens during 2013–2019 following introduction of FAdV-4 vaccines. Avian Pathology, (2021); 50(2): 182-189.
- Baneyx F. Recombinant protein expression in Escherichia coli. Current opinion in biotechnology, (1999); 10(5): 411-421.
- Shah M, Ashraf A, Rahman M, Khan M, Qureshi J. A subunit vaccine against hydropericardium syndrome using adenovirus penton capsid protein. Vaccine, (2012); 30(50): 7153-7156.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0