

Full Length Research Article
In Silico Study of Innovative Compounds Targeting Protein (Survivin): Understandings into Cancer Therapeutics
Abdullah Hamadi1*, Hyfa Alzahrani2, Fawzyah Albaldi2, Sallwa M Alshehre3, Ali Mahzari4
Adv. life sci., vol. 12, no. 2, pp. 362-370, May 2025
*– Corresponding Author: Abdullah Hamadi (Email: a.aldhafri@ut.edu.sa)
Authors' Affiliations
2. Department of Biology, Faculty of Science, Albaha University – Saudi Arabia.
3. Department of Clinical Laboratory Sciences, Faculty of Applied Medical Sciences, Umm Al-Qura University, PO Box 7607, Makkah – Saudi Arabia
4. Department of Laboratory Medicine, Faculty of Applied Medical Sciences, Al-Baha University, Al-Baha – Saudi Arabia
[Date Received: 20/10/2024; Date Revised: 05/01/2025; Date Available Online: 31/08/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Survivin, also known as Baculoviral Inhibitor of Apoptosis Repeat Containing 5 (BIRC5), is a key protein involved in cell division and the inhibition of apoptosis, making it an essential component of eukaryotic cell biology. Survivin is highly expressed across all human tumor lines and is significant in cancer due to its dual function in promoting cell proliferation and inhibiting apoptosis. Programmed cell death (PCD) is critical for mammalian development and disease regulation. The dysregulation of apoptosis leads to uncontrolled cell proliferation, which contributes to tumor progression and metastasis. Survivin interacts with caspases, predominantly inhibiting Caspase-3 and Caspase-7, thereby playing a key role in dysregulating apoptosis. Given its preferential expression in tumor cells and its decisive role in carcinogenesis and chemoresistance, Survivin emerges as an attractive target for cancer treatment.
Methods: Compounds from the PubChem database were screened using molecular docking. Lead candidates were then evaluated for drug-likeness and ADMET properties, and the stability of the top compound-protein complex was analyzed using molecular dynamics simulations.
Results: The current study highlights the therapeutic potential of novel compounds designed to inhibit Survivin in cancer therapy. This probability is illuminated through the use of innovative techniques such as computer-generated (virtual) screening and molecular docking. The lead compounds have also been filtered for their drug-likeness and ADMET potential.
Conclusion: The stability of the designated compounds has been investigated in-depth through MD simulations. The results from this study suggest that the compound CID:155104111 (1-[2-(difluoromethoxy)ethyl]-2-methyl-3-(pyrazin-2-ylmethyl)benzo[f]benzimidazol-3-ium-4,9-dione) is a potential lead candidate for targeting the Survivin protein, showing promise when compared to the standard inhibitor, YM155.
Keywords: Survivin; Apoptosis; Caspases; YM155; PubChem; Molecular Dynamics
Introduction![]()
The essential eukaryotic protein Survivin, sometimes referred to as BIRC5 (baculoviral inhibitor of apoptosis repeat containing 5), is involved in both cell division and cell death [1,2]. It consists of a total of 142 amino acids. This molecular entity exhibits a molecular weight of 16.5 kDa. Its structural composition encompasses three distinct protein chains, neatly arranged to display its functional properties [3, 4]. In particular, this molecular structure presents seven binding sites, with two among them serving as active sites, critical for its biological activities. Survivin is the smallest member of the Inhibitor of Apoptosis Protein (IAP) family. A family of 8 physically and functionally related proteins make up the human IAPs [2, 4]. The IAP family proteins are known for their multifaceted nature and are present in most eukaryotic organisms. Survivin is vastly expressed in most human cancers. The National Cancer Institute documented that Survivin is expressed across all 60 human tumor lines. As a protein, Survivin assumes a dual responsibility in both cell division and inhibiting programmed cell death also known as apoptosis [5, 6]. Programmed cell death performs a crucial role during mammalian cell development [2, 3]. It is a key protein that is involved with the cell signaling of multiple cancers [6,7]. These signaling molecules regulate and control their function. This allows it to influence cancer initiation and progression [5, 7]. Moreover, it assumes a pivotal role in survival but also the insidious spread of tumor cells and contributes to cancer progression and metastasis [7, 8]. Incomplete execution of apoptosis, a highly regulated process, can lead to hazardous conditions such as uncontrolled cell growth. The failure to fully execute apoptosis sets the stage for uncontrolled cell proliferation, leading the way for potential abnormalities and unwanted extension of cellular population [9-11]. Apoptosis is a conserved mechanism that unfolds uninterrupted across the scale of low to high-order eukaryotes [11, 12]. Currently, two prominent apoptotic pathways are present in mammals:(i) nonreceptor-mediated/intrinsic apoptosis and (ii) Receptor-mediated/extrinsic apoptosis. These complex mechanisms are meticulously controlled by an array of caspase enzymes, encompassing initiator/upstream caspases (Caspases 8 and Caspase 9) and executioner/downstream caspases (caspase3 and caspase 7) [12,13]. Survivin functions fundamentally as an anti-apoptotic protein by inhibiting the activity of caspases. Caspases are significant enzymes (cysteine proteases) that play an essential role in deconstruction of cellular structure during apoptosis. It acts as a molecular brake on apoptosis via directly interacting and inhibiting certain caspases, particularly Caspase 3 and 7. This inhibition serves as an effective indication to Survivin’s crucial role in actively suppressing the machinery of programmed cell death [14,15]. Due to its involvement in cell cycle promotion and apoptosis inhibition, Survivin is careful highly as a perfect candidate for targeted anticancer therapy [14, 16, 17]. Certainly, various strategies aimed at blocking its expression or functionality are appearing as highly promising therapeutic strategies in the field of cancer treatment [17, 18]. Normally, it is practically undetectable in most normal differentiated tissues but conspicuously expressed in malignancies of various cancer types. This distinct expression pattern offers a high level of selectivity for tumor cells, thereby reducing the likelihood of potential side effects in treated patients [19, 20]. The overexpression of Survivin has even now been associated with tumor prognosis. Various studies have highlighted the negative impact of its overexpression on both prognosis and resistance to therapeutic agents. Overall, its presence in tumors and its significant biotic roles in promoting carcinogenesis and resistance to chemotherapy make it a prominent therapeutic target [20-22]. Since it has emerged as an ideal target for cancer drug discovery, numerous Survivin inhibitors have been reported in the literature. Some of them are specifically unswerving inhibitors but others don’t bind directly to Survivin itself, and they target other biomolecules to reduce its expression. YM155 was the initial acknowledged small molecule inhibitor used to specifically target and suppress the activity of Survivin promoters [23, 24]. YM155 inhibits its expression at both protein and mRNA levels, also it is highly selective.YM155 suppresses the upstream transcription factors, SP1 and ILF3, therefore inhibiting their interaction with Survivin promoters [20, 25]. FL118 is another highly auspicious small molecule used for cancer treatment, particularly for colorectal and pancreatic cancer. This was discovered over HTS (high throughput screening) surveyed by in-vitro and in vivo hit to principal analysis [21, 25]. FL118 has displayed high activity in comparison to current treatment preclinical studies. It is a nonselective small molecule inhibitor of Survivin expression. FL118 illustrates effective inhibition of Survivin promoter activity, it also actively suppresses Survivin expression and disrupts cancer cell development at high picomolar to low nanomolar concentration [2, 14]. FL118 similarly inhibits the expressions of other cancer related IAPs, such as Mcl-1, XIAP and cIAP2. The exceptional anticancer efficacy of FL118 is linked to its steric configuration and existence of a free hydroxyl group at position 20 [12, 23, 24]. Thereby, YM155 and FL118 are currently recognized as notable inhibitors targeting the Survivin protein implicated in cancer treatment.
![]() Methods
Methods
The software and ligand library of Discovery Studios
Discovery studio software (Biovia, San-Diego, California, USA) represents a complete set of programs specially made for the simulation both small molecules and macromolecules system. It offers a wide range of functions specified toward Protein modelling, optimization, and drug design. It offers researchers with powerful tools for intricate molecular analysis and innovative therapeutic discoveries. The LibDock module of discovery studios was applied for virtual screening so that it can make use of its advanced capabilities for Molecular docking [15]. Furthermore, The ADME module of discovery studios was employed to carefully analyse the Pharmacological properties, ensuring a full understanding of compounds drug like characteristics and potential therapeutic activities. For ligand library PubChem (https://pubchem.ncbi.nlm.nih.gov/) database was selected. PubChem database is maintained by NCBI (National Centre for Biotechnology Information), It stands as a freely accessible database containing a vast collection of commercially available chemical compounds.
Data retrieval and Preparation
Firstly, the 1.40-Å crystal structure of human Survivin/BIRC5 (Protein databank identifier: 2QFA) was acquired from Protein Data Bank and is introduced in the operational environment of LibDock for further analysis. The protein was prepared by removing crystal water and other heteroatoms. The arranged protein was active to define the binding site. Optimize the protein structure by energy minimization to remove steric clashes and to optimize hydrogen bonding. Furthermore, a set of potential inhibitors are downloaded from PubChem (https://pubchem.ncbi.nlm.nih.gov/ ) in 3D conformer state in SDF format based on structural similarities to known inhibitors. The filtered ligands were then prepared through protonation and energy minimization to generate low-energy conformations.
Adsorption, Distribution, metabolism, Excretion and Toxicity prediction
The ADME module of discovery studios was used to precisely calculate the crucial parameters such as Absorption, distribution, metabolism and excretion (ADME) properties of particular compounds [26,27]. This analysis also includes the evaluation of aqueous solubility, Blood-brain barrier permeability, inhibition of cytochrome p-450 2D6(CYP2D6), hepatotoxicity, human intestinal absorption and plasma-protein binding affinity. These detailed analyses provide critical information about ligand description and potential therapeutic outcomes of compounds [28-30]. The TOPKAT module of Discovery Studio plays an important role in estimating the toxicity and various other properties of the candidate compounds. Through this pharmacological assessment, we can find compounds with therapeutic potential [31].
Molecular Docking
The software (LibDock module of discovery studios) is used for the purpose of MD studies. This module helps high throughput docking, consenting for the quick screening of compounds to assess their probable as inhibitors founded on their aptitude to dock efficiently at the binding site of Survivin. LibDock finds significant regions on a protein, known as hotspots, which serve as the finest locations for ligand binding. These hotspots support the arrangement of ligands, indicating favorable interactions. This alignment is attained through the use of a grid situated within the binding site. For a more active and professional tone, rephrase to: "To refine the ligand alignment and optimize its interaction with the protein, a ligand minimization process was performed. This method uses the CHARMM force field and develops the Smart Minimizer algorithm, ensuring accurate and efficient ligand placement within the binding site. Subsequent to the minimization procedure, the ligand poses were ranked based on their corresponding ligand scores [27, 29, 30]. The preferred compounds endured rigorous screening (virtual) against the target protein, and the results were accurately analyzed. Resulting this examination, the top-performing compound was carefully selected for further assessment. To evaluate its binding affinity with the protein, the compound was docked using the software [20, 29, 31, 32]. Now, higher LibDock scores corresponded to more affinity between the compound and the target protein. Entirely candidate compounds were classified giving to the LibDock score.
Molecular Dynamics Simulation
MD simulation is a bioinformatics-based technique used in the field of structural biology and discovery. It's working to expect the ideal conformation and binding affinity of a small molecule to a protein. Molecular dynamics (MD) simulation operates on the foundational principles of Newton's laws of motion and statistical mechanics, providing a detailed understanding of molecular behaviour in various environments [33,34]. Molecular Dynamics (MD) simulation was applied to thoroughly assess the stability of the protein-ligand complex derived from the Discovery Studio docking procedures. Through MD simulation, intricate molecular interactions and dynamic behaviour within the complex were explored, providing valuable insights into its stability and structural dynamics over time [35,36,37]. The ideal binding conformation of ligand-Survivin complexes among the results projected by the molecular docking database, are carefully designated and are arranged for molecular dynamic simulation using Biovia Discovery Studio software. The results were anlayzed through RMSD and RMSF analysis.
Screening of PubChem database virtually against Survivin
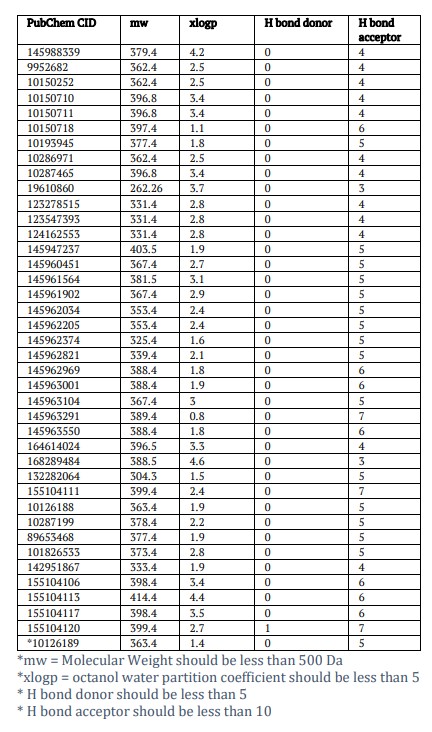
A total of 61 chemical compounds were acquired from the PubChem database based on database similarity searching using YM155 as a standard inhibitor. The structure (chemical) of YM155 was nominated as the standard control and its pharmacological properties were related to additional compounds. After further screening only 39 compounds were shown to demonstrate optimal properties for pharmacological reference. The Lipinski’s rule of five was used to filter these 39 compounds for further ADMET analysis (Table 1).
ADME and Toxicity prediction
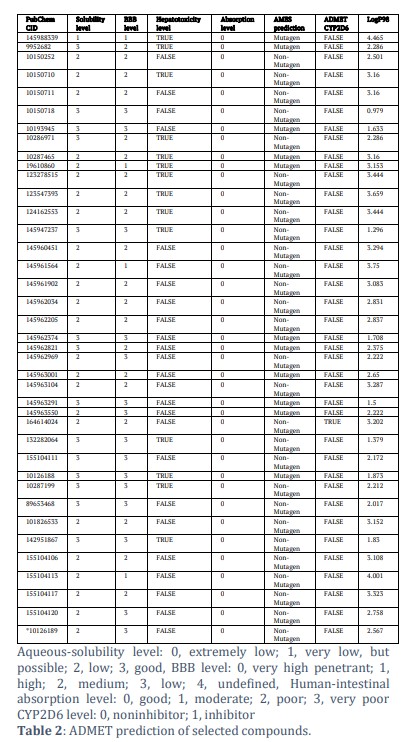
Ensuring safety is also very important in this research endeavour. To guarantee the safety of the chosen compound, a thorough evaluation of its toxicity is very important. Using the TOPKAT module of discovery studios, toxicity indicators of various compounds and YM155 will be carefully analysed. The ADME module of discovery studios was used to expect the pharmacological characteristics of all chosen ligands and YM155. The Aqueous solubility estimate indicates that all compounds show solubility in water [26,28,31]. Furthermore, the prediction suggested that none of the compounds would act as an inhibitor against CYP2D6. For hepatotoxicity 24 compounds are found to be non-toxic, which prove to be less toxic than YM155. The remaining compounds were toxic in nature and were left. For human intestinal absorption, all the compounds have approximately the same level of absorption as YM155 [29,30]. Results show that a total of 27 compounds were discovered to be non-mutagenic. Considering all the above results, we have identified these three lead compounds CID-145962969, 155104111, and 10150718 as ideal candidates, determined by their non-CYP2D6 inhibitory properties and absence of hepatotoxicity (Table 2). Furthermore, our compounds exhibit low Ames mutagenicity and developmental toxicity potential. In total CID-145962969, CID-155104111 and CID-10150718 have been validated as safe candidates as for further studies.
Examination of ligand binding
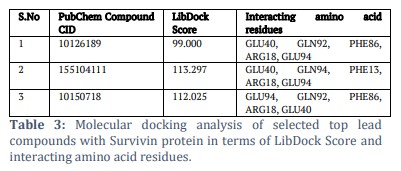
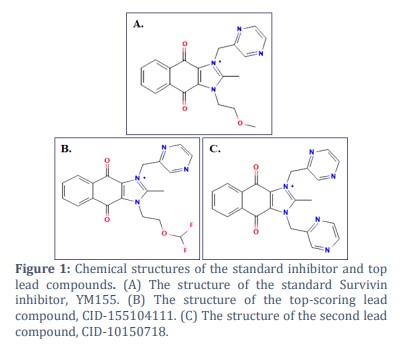
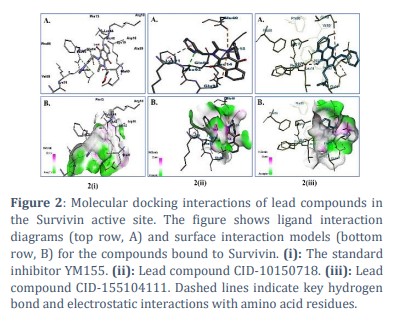
To study the ligand binding mechanism of screened compounds along with YM155, molecular docking was performed against Survivin using the LibDock module of Biovia Discovery Studio, followed by LibDock score calculation [23,24]. The 1.40Å crystal structure of human Survivin retrieved from the PDB (Protein Data Bank) was cleaned and prepared accordingly and seamlessly integrated into the operational environment of LibDock for further analysis and docking simulations (Figure 1) [18, 25]. The ligands were also energy minimized and subjected to the docking interface in parallel. The best leads obtained after docking involved CID-10150718 with LibDock score of 112.025, CID-155104111 with LibDock score 113.297 (Table 3). The LibDock score of CID-10150718 and CID-155104111 was found to be better than the reference ligand YM155 (LibDock Score 99), which demonstrates that proposed compound has a good binding affinity with Survivin (24,26). Furthermore, the proposed leads were found to be docked in the active site of Survivin protein in a like manner like YM155 with surrounding amino acid residues reported as GLN92, PHE86, GLU40, ARG18, PHE93 and LEU14 (Figure 2 (i), (ii) and (iii).
Molecular Dynamics Simulation
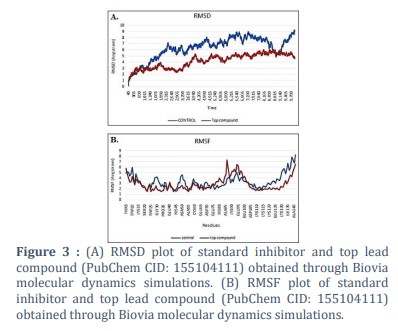
Molecular dynamics (MD) simulation operates on the foundational principles of Newton's laws of motion and statistical mechanics, providing a detailed understanding of molecular behaviour in various environments [38,39,40]. The stability of ligand-Survivin complexes was carefully investigated through MD, faithfully mirroring their behaviour in the ordinary environment. Following the MD procedures, optimum structural conformations of ligand-Survivin complexes are obtained [40,41]. Simulation studies were conducted on the leading compound-complex (CID: 155104111 bound to Survivin) compared with the standard drug YM155 bound to Survivin protein to validate the structural specificity of the protein-ligand interaction, particularly focusing on active site engagements with the protein. Optimal configurations of the primary candidates were chosen for a 10-nanosecond MD simulation. The MD paths were meticulously evaluated to evaluate time-dependent features, as well as the RMSD (root mean square deviation) of all backbone atoms and ligands, as well as the average fluctuations of the residues (RMSF) (Figure 3 A). The protein’s conformational stability throughout the simulation was assessed by computing the RMSD of backbone atoms. Examination of the trajectories revealed that the RMSD values for the top lead complex exhibited superior stability compared to the YM155 complex, with an average RMSD of less than 4.0 Å, indicating comparable stability to the standard drug complex [41,42].
In our study, we meticulously analyzed the RMSF (root mean square fluctuation) of both our top lead complex and the control compound, YM155, utilizing the trajectory module of Biovia Discovery Studio. RMSF serves as a crucial metric for assessing the dynamic behavior of individual residues within a protein structure during molecular dynamics (MD) simulations. Upon scrutinizing the RMSF values, we noted that both the control and the lead complex exhibited deviation from the standard value of 3.0 Å (Figure 3 B). This observation suggests that the protein-ligand complexes maintained a stable conformation throughout the specified simulation duration of 10 nanoseconds [43-45].
Specifically, our analysis revealed slight fluctuations in residues Ala 85, Ala 140, and Lys 90 within both complexes. However, these fluctuations did not significantly deviate from the anticipated range. Notably, residues associated with the active site of the protein displayed consistent RMSF profiles throughout the simulation period. Overall, our findings underscore the robust stability of both the lead complex and the control compound, highlighting their potential suitability for further investigation in the context of cancer therapeutics.
![]() Discussion
Discussion
Survivin, also known as BIRC5, is a eukaryotic protein involved in cell division and apoptosis. As the smallest member of the inhibitor of apoptosis protein (IAP) family, which includes eight structurally related proteins in humans, Survivin is notably overexpressed in many human cancers [16, 17]. Survivin plays a dual role by promoting cell division and inhibiting apoptosis, both critical for mammalian cell development. It is involved in various cancer-related signaling pathways, influencing cancer initiation and progression. Survivin inhibits apoptosis by directly interacting with and inhibiting caspase-3 and caspase-7, effectively suppressing the apoptotic process [16,20]. It operates at the crossroads of numerous cancer cell signaling networks and is regulated by a variety of upstream signaling molecules, including binding proteins, protein regulators, innumerable enzymes (proteases, kinases, phosphatases), transcription factors, miRNAs, transporter and channel proteins, receptors with or without kinase activity, and their corresponding ligands. Conversely, Survivin can also regulate many of these upstream molecules, forming an intricate network of interactions [22,24,25,26]. However further studies are needed to fully appreciate the significance of these interactions and their consequences in cancer to develop effective cancer therapeutic strategies [47-49].
In our study, 61 compounds were initially identified from the PubChem database based on their similarity to YM155, a commonly known inhibitor of Survivin. After further screening, 39 compounds were selected for pharmacological evaluation using Lipinski’s rule of Five and ADMET (Absorption, Distribution, Metabolism, Excretion, and Toxicity) analysis. The ADMET analysis revealed that all compounds were soluble in water, none inhibited CYP2D6 (an enzyme important in drug metabolism), 24 compounds were non-toxic to the liver, demonstrating lower toxicity than YM155, and all compounds had absorption levels similar to YM155. Additionally, 27 compounds were found to be non-mutagenic. Based on these evaluations, two lead compounds were identified as ideal candidates: CID-155104111 and CID-10150718. These compounds also exhibited low Ames mutagenicity and developmental toxicity potential.
To understand the binding mechanisms of the screened compounds, molecular docking was performed against Survivin using the LibDock module of Biovia Discovery Studio. The 1.40Å crystal structure of human Survivin was retrieved from the Protein Data Bank (PDB) and used for docking simulations. The compounds were energy minimized and docked to assess their binding affinity. CID-10150718 and CID-155104111 had better LibDock scores than YM155, indicating good binding affinity with Survivin. These compounds docked in the active site of Survivin similarly to YM155, were found in important interactions with key amino acid residues such as GLN92, PHE86, GLU40, ARG18, PHE93, and LEU14. Furthermore, MD simulations were conducted to investigate the stability of ligand-Survivin complexes. The lead compound (CID-155104111) and the standard drug YM155 bound to Survivin were subjected to 10-nanosecond MD simulations. The RMSD values showed that the lead compound complex exhibited greater stability than the YM155 complex, with an average RMSD of less than 4.0 Å. The MD simulation trajectories were analyzed to evaluate time-dependent features, including the Root Mean Square Deviation (RMSD) of backbone atoms and the Root Mean Square Fluctuation (RMSF) of individual residues. RMSD examination is critical as it makes available understanding into the conformational stability of the protein-ligand complex over time. By means of calculating the RMSD of the backbone atoms, scholars can evaluate how much the protein structure deviates from its early conformation during the simulation.
Moreover, RMSF investigation was performed to assess the average fluctuations of individual residues inside the protein during the simulation. This study supports find regions of the protein that show more flexibility or are more dynamically active, which can be serious for understanding the binding interactions and general stability of the protein-ligand complex. In this study, the RMSF values providing additional validation of the stability and interaction patterns experiential with the prime compound compared to YM155. The full examination of simulation and the subsequent study make available an inclusive picture of how the ligand-Survivin complexes perform over time. This comprises seeing the stability of the complexes, the dynamic comportment of individual residues, and the general conformational changes of the protein. Such understandings are invaluable for drug discovery and development, as they support find compounds with the maximum favorable stability and binding characteristics. Upon studying the RMSF values, we noted that both the control compound and the principal complex exhibited deviation from the standard value of 3.0 Å which recommended that the protein-ligand complexes kept a stable conformation during the specified simulation duration of 10 nanoseconds (ns). The little RMSF values athwart the mainstream of residues also showed that the overall structure of the protein endured comparatively rigid and did not endure important conformational variations through the simulation. The fact that the lead compound showed stability comparable to a known drug highlights its potential efficacy and suitability for further development. The negligible deviations in RMSF values, mainly at perilous residues, submit that the chief compound does not induce momentous structural alarms in the protein, which is a necessary representative for drug candidates. Exactly, our study discovered small variations in residues Ala 85, Ala 140, and Lys 90 inside both complexes.
To précis, the simulations (MD) confirmed the structural specificity and stability of the protein-ligand interactions for the prime compound CID-155104111 bound to the target protein Survivin. The analyses of RMSD and RMSF highlighted the stability of the active site engagements and the overall conformational integrity of the protein-ligand complex. These results underscore the probability of CID-155104111 as a capable candidate for further investigation in the context of cancer therapeutics targeting Surviving protein, providing a strong foundation for future experimental validations and drug development efforts.
In conclusion, Survivin emerges as a highly promising candidate for targeted anticancer therapy due to its involvement in cell cycle promotion and apoptosis inhibition. Its distinct expression pattern, predominantly in various cancer types while being almost undetectable in normal tissues, offers a level of selectivity that minimizes probable side belongings in treated patients. Furthermore, Survivin overexpression has been interrelated with poor tumor prognosis and resistance to therapeutic agents, underscoring its significance as a therapeutic target. Various strategies aimed at blocking Survivin expression or functionality are being explored, with inhibitors like YM155 and FL118 showing considerable promise but lack expected efficacy. By exploiting the vast chemical search space of the PubChem database and using computational techniques, this study proposes a new inhibitor of Survivin protein with PubChem CID as 155104111. The proposed lead expresses considerable binding efficacy as well as conformational stability with Survivin protein in comparison to its standard inhibitor, highlighting it as a promising therapeutic agent against the same.
Tables and Figures
Author Contributions
AH and, HA: Manuscript drafting editing and proof reading, FA, SMS, and, AM: Data analysis and revision.
- Blanco Wheatley SP, , Altieri DC. Survivin at a glance. Journal of Cell Science, (2019); 132(7), jcs223826.
- Martínez-Sifuentes MA, Bassol-Mayagoitia S, Nava-Hernández MP, Ruiz-Flores P, Ramos-Treviño J, et al. Survivin in Breast Cancer: A Review. Genetic Testing and Molecular Biomarkers , (2022); 26(9): 411-421.
- Shojaei F, Yazdani-Nafchi F, Banitalebi-Dehkordi M, Chehelgerdi M, Khorramian-Ghahfarokhi M. Trace of Survivin in cancer. European Journal of Cancer Prevention. (2019); 28(4): 365-372.
- Shomali N, Suliman Maashi M, Baradaran B, Daei Sorkhabi A, Sarkesh A, et al. Dysregulation of Survivin-Targeting microRNAs in Autoimmune Diseases: New Perspectives for Novel Therapies. Frontiers in Immunology. (2022); 13: 839945.
- Bernardo PS, Lemos LGT, de Moraes GN, Maia RC. Unraveling Survivin expression in chronic myeloid leukemia: Molecular interactions and clinical implications. Blood Reviews. (2020); 43: 100671.
- Lyu H, Huang J, He Z, Liu B. Epigenetic mechanism of Survivin dysregulation in human cancer. Science China Life Sciences. (2018); 61(7): 808-814.
- Duffy MJ, O'Donovan N, Brennan DJ, Gallagher WM, Ryan BM. Survivin: a promising tumor biomarker. Cancer Letters. (2007); 249(1): 49-60.
- Braný D, Dvorská D, Kúdela E, Danková Z, Halášová E, Višňovský J. Potential of Survivin for treatment of gynaecological tumour diseases. Ceska gynekologie. (2018); 83(3): 226-231.
- Warrier NM, Agarwal P, Kumar P. Emerging Importance of Survivin in Stem Cells and Cancer: the Development of New Cancer Therapeutics. Journal of Haematology and Stem Cell Research. (2020); 16(5): 828-852.
- Li F. Survivin study: what is the next wave? Journal of cellular physiology, (2003);197(1): 8-29.
- Altieri DC. Validating Survivin as a cancer therapeutic target. Nature reviews Cancer, (2003); 3(1):46-54.
- Fang XL, Cao XP, Xiao J, Hu Y, Chen M, et al. Overview of role of Survivin in cancer: expression, regulation, functions, and its potential as a therapeutic target. Journal of Drug Targeting, (2024); 32(3): 223-240.
- Santarelli A, Mascitti M, Lo Russo L, Sartini D, Troiano G, et al. Survivin-Based Treatment Strategies for Squamous Cell Carcinoma. International Journal of Molecular Sciences, (2018); 19(4): 971
- Albadari N, , Li W. Survivin small molecules inhibitors: Recent advances and challenges. Molecules, (2023); 28(3): 1376.
- Warrier NM, Agarwal P, Kumar P. Emerging importance of Survivin in stem cells and cancer: The development of new cancer therapeutics. tem Cell Reviews and Reports. (2020); 16(5): 828-852.
- Mahmoudian-Sani M, Alghasi A, Saeedi-Boroujeni A, Jalali A, Jamshidi M, Khodadadi A. Survivin as a diagnostic and therapeutic marker for thyroid cancer. Pathology Research and Practice, (2019); 215(4): 619-625.
- Li F, Aljahdali I, Ling X. Cancer therapeutics using Survivin BIRC5 as a target: what can we do after over two decades of study. Journal of Experimental , Clinical Cancer Research, (2019); 38, 368.
- Babkoff A, Cohen-Kfir E, Aharon H, Ronen D, Rosenberg M, et al. A direct interaction between Survivin and myosin II is required for cytokinesis. Journal of Cell Science, (2019); 132(14): jcs233130.
- Park S-H, Shin I, Park S-H, Kim ND, Shin I. An Inhibitor of the Interaction of Survivin with Smac in Mitochondria Promotes Apoptosis. Chemistry – An Asian Journal, (2019); 14(22), 4035-4041.
- Sarvagalla S, Lin T, Kondapuram SK, Cheung CHA, Coumar MS. Survivin – caspase protein-protein interaction: Experimental evidence and computational investigations to decipher the hotspot residues for drug targeting. Journal of Molecular Structure, (2021); 1229(4): 129619.
- Szklarczyk D, Gable AL, Lyon D, Junge A, Wyder S, et al. STRING v11: Protein-protein association networks with increased coverage, supporting functional discovery in genome-wide experimental datasets. Nucleic Acids Research, (2019); 47(D1).
- Martínez‐García D, Manero‐Rupérez N, Quesada R, Korrodi‐Gregório L, , Soto‐Cerrato V. Therapeutic strategies involving Survivin inhibition in cancer. Medicinal Research Reviews, (2019); 39(3): 887-909.
- Wang Q, Arnst K, Xue Y, Lei Z, Ma D, et al. Synthesis and biological evaluation of indole-based UC-112 analogs as potent and selective Survivin inhibitors. European Journal of Medicinal Chemistry, (2018); 149: 211-224.
- Yu X, Zhang Y, Cavazos D, Ma X, Zhao Z, et al. MiR-195 targets cyclin D3 and Survivin to modulate the tumorigenesis of non-small cell lung cancer. Cell Death and Disease, (2018); 9(2): 193.
- Vay C, Babaei S, Safi S, Dizdar L, Rehders A, et al. Clinicopathological and prognostic value of Survivin expression in surgically resected pancreatic ductal adenocarcinoma. Cancers, (2022); 14(14): 3494.
- Adamopoulos P, Tsiakanikas P, Adam E, , Scorilas A. Unraveling novel Survivin mRNA transcripts in cancer cells using an in-house developed targeted high-throughput sequencing approach. Genomics, (2021); 113(1): 573-581.
- Lv X, Jiang Y, Wang X, Xie H, Dou G, et al. Computational study on novel natural inhibitors targeting BCL2. Medical Oncology, (2021); 38(8): 94.
- Komura H, Watanabe R, Mizuguchi K. The Trends and Future Prospective of In Silico Models from the Viewpoint of ADME Evaluation in Drug Discovery. Pharmaceutics, (2023); 15(11): 2619.
- Shou WZ. Status and future directions of high-throughput ADME screening in drug discovery. Journal of Pharmaceutical Analysis, (2020); 10(4): 1.
- Daoud N, Borah P, Deb P, Venugopala K, Hourani W, et al. ADMET Profiling in Drug Discovery and Development: Perspectives of In Silico, In Vitro and Integrated Approaches. Current Drug Metabolism, (2021); 22(7), 503-522.
- Zhong S, Zhang Z, Guo Z, Yang W, Dou G, et al. Identification of novel natural inhibitors targeting AKT Serine/Threonine Kinase 1 (AKT1) by computational study. Bioengineered. (2022); 13(5): 12003-12020.
- Raja T, Ravikumar P, Srinivasan MR, Vijayarani K, Kumanan K. Identification of Potential Novel Inhibitors for Nipah Virus – An in-silico Approach International Journal of Current Microbiology and Applied Sciences. (2020); 9(9): 3377-3390.
- Jia C-Y, Li J-Y, Hao G-F, Yang G-F. A drug-likeness toolbox facilitates ADMET study in drug discovery. Drug Discovery Today, (2020); 25(1): 248-258.
- Barbhuiya S, Das BB. Molecular dynamics simulation in concrete research: A systematic review of techniques, models, and future directions. Journal of Building Engineering, (2023); 107267.
- Ghosh S, Chetia D, Gogoi N, Rudrapal M. Design, molecular docking, drug-likeness, and molecular dynamics studies of 1,2,4-trioxane derivatives as novel plasmodium falciparum falcipain-2 (Fp-2) inhibitors. Biotechnologia, (2021); 102(3): 257-275.
- Raychaudhury C, Srinivasan S, Pal D. Identification of potential oral cancer drugs as Bcl-2 inhibitors from known anti-neoplastic agents through docking studies. Journal of Mathematical Chemistry, (2024); 62(2): 317-329.
- Liu N, Wang X, Wu H, Lv X, Xie H, et al. Computational study of effective matrix metalloproteinase 9 (MMP9) targeting natural inhibitors. Aging, (2021); 13(19). 22867–22882.
- Ferreira LLG, Andricopulo AD. ADMET modeling approaches in drug discovery. Drug Discovery Today, (2019); 24(5): 1157-1165.
- Adelusi TI, Oyedele A-QK, Boyenle ID, Ogunlana AT, Adeyemi RO, et al. Molecular modeling in drug discovery. Informatics in Medicine Unlocked, (2022); 29. 100880.
- Papaleo E, Saladino G, Lambrughi M, Lindorff-Larsen K, Gervasio FL. The role of protein loops and linkers in conformational dynamics and allostery. Chemical Society Reviews, (2016); 45(22), 22-42.
- Roe DR, Cheatham III TE. PTRAJ and CPPTRAJ: software for processing and analysis of molecular dynamics trajectory data. Journal of chemical theory and computation, (2013); 9(7): 3084-3095.
- Gapsys V, de Groot BL, Briones R. Quantifying flexibility in protein structures: comparison of experimental and theoretical B-factor distributions. Biophysical journal, (2015); 108(10): 2585-2595.
- Zhong Q, Qin J, Zhao K, Guo L, Li D. Molecular Docking and Molecular Dynamics Simulation of New Potential JAK3 Inhibitors. Current Computer-Aided Drug Design, (2023); 20 (6): 764-772.
- Miao H, Klein T, Kouřil D, Mindek P, Schatz K, et al. Multiscale Molecular Visualization. Journal of Molecular Biololgy, (2019); 431(6):1049-1070.
- Sargsyan K, Grauffel C, Lim C. How Molecular Size Impacts RMSD Applications in Molecular Dynamics Simulations. Journal of Chemical Theory and Computation, (2017); 13(4): 1518-1524.
- Laasonen K. Ab initio molecular dynamics. Methods in Molecular Biology, (2013); 924: 29-42.
- Giodini A, Kallio MJ, Wall NR, Gorbsky GJ, Tognin S, et al. Regulation of microtubule stability and mitotic progression by Survivin. Cancer Research, (2002); 62(9): 2462-2467.
- Lv Y-G, Yu F, Yao Q, Chen JH, Wang L. Role of Survivin in diagnosis, prognosis, and treatment of breast cancer. Journal of Thoracic Disease, (2010); 2(2): 100-110.
- Savitskaya MA, Onishchenko GE. Mechanisms of Apoptosis. Biochemistry, (2015); 80(11): 1393-405.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0