

Full Length Research Article
Reciprocal relationship between neurodegeneration-related miRNAs and neurohormones in diabetes mellitus patients: The possible mechanism of high glucose neurotoxicity
Toqa Alabyadh, Leila Sadeghi*, Mohammad Ali Hosseinpour Feizi
Adv. life sci., vol. 12, no. 3, pp. 540-546, August 2025
*- Corresponding Author: Leila Sadeghi (Email: l.sadeghi@tabrizu.ac.ir)
Authors' Affiliations
[Date Received: 10/12/2023; Date Revised: 28/02/2025; Available Online: 31/10/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Diabetes mellitus (DM) is a metabolic syndrome that causes blood glucose to remain consistently above normal. High glucose concentrations in the neuronal niche can harm neurons permanently. Some DM-related molecular events are found to cause neurological abnormalities and cognitive dysfunctions. Dopamine and oxytocin are important neurohormones that are regulated by miRNAs and could also reorganize neural function-related miRNA networks. Therefore, the purpose of this study was to assess how neurohormones and miRNAs linked to neurodegeneration interact in DM patients.
Methods: For this purpose, blood samples related to type 1 (T1DM) and type 2 (T2DM) of diabetes and non-diabetic controls were used to measure hormones (dopamine, oxytocin, and thyroid hormones) by using a specific ELISA kit and circulating miRNAs by RT-PCR method.
Results: Data revealed a significant reduction of dopamine and oxytocin in both types of DM, which are accompanied by miR-27a, miR-107, and miR-191 up-regulation. PTEN-induced putative kinase 1 (PINK1) protein expression is primarily inhibited by miR-27a, which leads to dopaminergic cell death and consequently reduces dopamine synthesis and release. We also found a significant decrease in miR-23a and miR-128 levels in DM that may promote dopaminergic cell vulnerability, possibly through attenuation of mitochondrial complex I. Comparing the data related to both types of DM confirmed that miR191 and miR-128 levels in T2DM are higher than in T1DM.
Conclusion: It was discovered that miRNAs and neurohormones have a reciprocal interaction that may make them a promising treatment target for DM.
Keywords: Diabetes mellitus, Neurodegeneration, miRNAs, Dopamine, Oxytocin, Diabetes related neurotoxicity
Introduction![]()
Diabetes mellitus (DM) is a multifactorial metabolic disorder known for increased glucose in circulation [1]. The most common classifications include type 1 (T1DM) and type 2 (T2DM). In contrast to T2DM, which is associated with dyslipidemia, central obesity, hypertension, insulin resistance, and a relative deficiency of insulin secretion, T1DM is caused by an absolute deficiency in pancreatic beta-cell function in most cases [1]. Therefore, diabetic patients experience two main molecular conditions: insulin deficiency and glucose dysregulation, both affecting neural cells [2,3]. Increased glucose disrupts homeostasis throughout the body, particularly in the central and peripheral neurological systems, mostly through the non-enzymatic glycation of functional molecules [4]. Our previous studies confirmed that non-enzymatic glycation in high glucose medium affects acetylcholinesterase enzyme kinetic behavior and changes its interaction with drugs [4,5]. Of course, other proteins and peptides also could be impacted by non-enzymatic glycation, so DM is a systemic disorder accompanied by multiple symptoms in the whole body, especially the nervous system, due to more sensitivity [6]. Insulin is a peptide hormone with pleiotropic actions that play an indispensable role in the regulation of development, dendritogenesis, and metabolism rate in neural cells [3,7]. The mentioned evidence and unknown molecular events cause chronic neurological abnormalities in diabetic patients, which manifest different degrees of diabetic neuropathy and are also more labile against neurodegenerative disease and cognitive dysfunctions [8,9]. The most common type of abnormalities in the nervous system is mediated by neural hormones such as thalamic, hypothalamic, and thyroid hormones, which work together to regulate brain development and function in response to metabolism rate [8,10]. Previous experiments confirmed that the malfunction of oxytocin plays an important role in glucose uptake, insulin resistance, and dyslipidemia [11]. Given the significance of neurohormones in brain function, an assessment of oxytocin and dopamine, thyroxine (T4), and triiodothyronine (T3) hormones in DM could give valuable information about the neuroendocrine impact on the neural abnormalities observed in diabetic patients.
Recent research revealed the crucial role of the microRNAs (miRNAs) in DM pathogenesis, and miR375, miR409, miR155, etc, were introduced as biomarkers of DM [12,13]. MicroRNAs generally interact with the 3′untranslated region (3′-UTR) of target mRNAs and inhibit gene expression. Consequently, one miRNA has the potential to regulate several genes that are involved in distinct biological pathways [14]. Increasing numbers of neurodevelopment-related miRNAs have been found in both the in vitro and in vivo models of DM [15,16]. Consequently, some of the changed miRNAs in DM are responsible for neuropathy and cognitive abnormalities.
This study was designed to investigate the neurohormonal abnormalities in T1DM and T2DM, which had not been studied yet. For this purpose, the concentration of oxytocin, dopamine, T3, and T4 hormones was evaluated in serum samples of DM patients and also non-diabetic controls who did not experience high blood glucose and neurological abnormalities. We also aimed to evaluate some miRNAs related to neurological disease in both types of DM and control. We wanted to investigate possible interactions between neurohormones and miRNAs in DM. This area of research has been more interesting recently because it could produce a reasonable relationship between hormone function and the regulation of synthesis and release by miRNAs and help to clarify the noncoding RNAs’ role in DM.
Methods![]()
Experimental design
This cross-sectional study included 40 T1DM patients, 40 T2DM patients, and 40 non-diabetic controls designated to the Clinical Laboratory of Al Zahraa Teaching Hospital, Kut, Iraq. Our selected samples in the T2DM group were newly diagnosed patients, and a blood sample was drawn before medical treatment (uncontrolled diabetes). Ethical approval and informed consent declarations were acquired from all individuals. Eligible participants for inclusion (T1DM and T2DM) were individuals above 17 years of age. Participants with illnesses like hepatic failure, renal disease, thyroid disease, acute coronary syndrome, Alzheimer’s disease, malignancies, or a family history of dementia were excluded. Controls were healthy subjects matched for sex and did not include individuals with diabetes, prediabetes, cardiovascular disease, thyroid dysfunction, Alzheimer’s disease, or other neurological disorders. The age of patients with T1DM and T2DM was calculated to be 20.30 ± 2.43 years and 45.90 ± 11.40 years, respectively, and the control group was 34.54 ± 15.68 years. The number of men and women was similar in the three experimental groups, and statistical analysis did not reveal any significant differences.
Around 10 milliliters of venous blood were collected in an anticoagulant tube containing sodium EDTA. Every participant had their serum collected in a second shared tube. Before being used for further analysis, the serum underwent centrifugation at 1500 g for 10 minutes at 4°C, subsequently divided into small aliquots, and stored at -20°C.
Hormone measurement method
Hormones, including triiodothyronine (T3), Thyroxine (T4), thyroid-stimulating hormone (TSH), dopamine, and oxytocin, were quantified by using a specific ELISA kit in all the serum samples. Neurohormones like dopamine and oxytocin were quantified using specific ELISA kits manufactured by MyBioSource Company, USA (MBS494471 and MBS2700454). T3, T4, and TSH ELISA kits were prepared by Agappe Company.
Analysis of the miRNA content of blood samples using real-time PCR (RT-PCR)
The RT-PCR technique was used to evaluate miRNA levels in plasma samples, and U6 noncoding RNA was considered as an internal control [17]. According to the guidelines provided by the manufacturer, for this, total RNA was extracted from cells using the TRIzol LS reagent. The quality control of the isolated RNA was evaluated using a NanoDrop UV/VIS spectrophotometer. The TaqMan Reverse Transcription Kit was used to transform total RNA into complementary DNA (cDNA). Then, using a SYBR Green PCR Kit (Toyobo, Tokyo, Japan) and specific primers purchased from Sangon Biotech (Shanghai, China), qRT-PCR was performed on an ABI Prism 7500 Detection System (Applied Biosystems) (Table 1). U6 was employed as an internal reference for miRNAs because of its steady expression. After normalization, the relative expression of miRNAs was evaluated using the 2−ΔΔCt approach [17].
Analysis of statistics
Statistical evaluation was performed using Brown-Forsythe ANOVA test to determine statistical differences between the results of T1 and T2 diabetic patients and non-diabetic samples as controls. The three experimental groups were compared using Tukey's multiple comparison test; asterisks show the matching p-value for significant differences in each figure. GraphPad Prism (version 9.5, GraphPad Software, Inc., San Diego, CA, USA) was used for all statistical analyses.
Results![]()
Considering the important role of hyperglycemia on mood and cognition, especially memory and dementia [2,3], this study aimed to explore the possible mechanisms of neurohormonal and miRNA levels to elucidate some of the neural complexities involved in diabetic patients and related animal models. To this end, we collected data from 40 type 1 diabetic patients, 40 type 2 diabetic patients, and 40 non-diabetic controls, and then biochemical parameters and miRNA content were evaluated in all samples.
Effect on the circulating level regulation of neurohormones in Diabetes
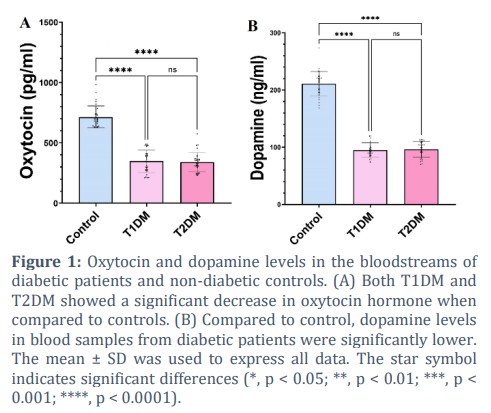
Oxytocin and dopamine are neurohormones that could regulate the cholinergic system; they also regulate energy homeostasis and have interesting potential as a target for metabolic disease treatment according to recent studies [18,19]. However, the regulation and alteration of oxytocin and dopamine in diabetes have not been studied yet, and its role in the neuropathology of diabetes needs additional investigation. The results of the current study showed oxytocin level is 714.85 ± 98.39 pg/mL (0.714 ± 0.098 ng/mL) in non-diabetic control blood samples, which decreased to 348.43 ± 92.98 pg/mL in T1DM; this reduction is significant according to multiple comparisons of one-way ANOVA statistical analysis. This parameter for T2DM patients was 339.56 ± 79.80 pg/mL, which is significantly less than that of the control (Fig. 1A). Although the oxytocin levels reduced in DM patients, the circular concentration of oxytocin in both diabetic groups is similar to each other without a remarkable difference (p > 0.05).
Figure 1B shows the dopamine concentration of blood serum in three experimental groups. According to the results, dopamine concentration is 210.98 ± 21.38 ng/mL in non-diabetic controls, which was reduced more than 2-fold in both of the diabetic groups. Dopamine concentration was measured to be 95.08 ± 12.93 ng/mL and 96.32 ± 13.78 ng/mL in types 1 and 2 of diabetes, respectively.
Effect on thyroid hormones by high glucose toxicity
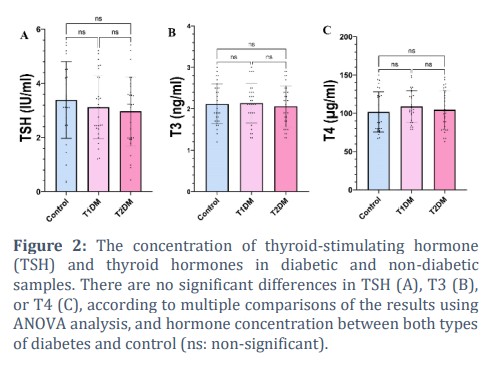
Thyroid hormones also could affect glucose metabolism rate by directly controlling insulin secretion from beta cells and indirectly the clearance of circulating insulin [20]. Previous reports confirmed that underactivity of the thyroid gland reduces glucose-induced insulin secretion, whereas hyperthyroidism enhances the response of beta cells to glucose [21]. Our results revealed thyroid hormone levels don’t have significant changes in the three experimental groups (Fig. 2). This means the average level of TSH, T3, and T4 hormones is similar to each other, and one-way ANOVA could not detect any significant changes.
Effect on circulating level regulation of neurodegeneration-related miRNAs in Diabetes
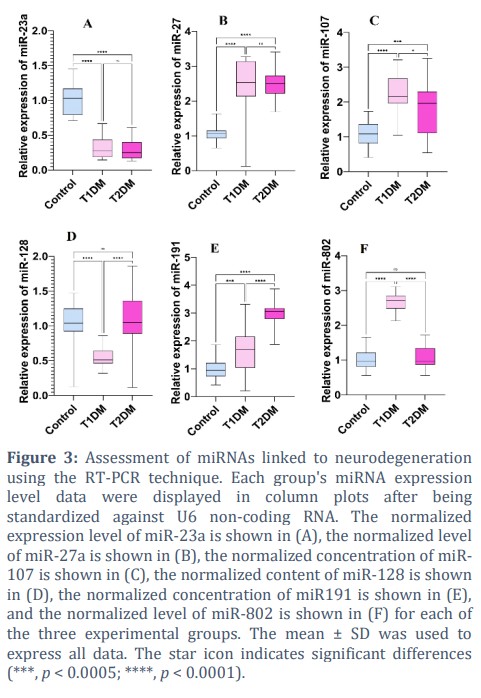
Recent studies confirmed miRNAs are widely changed in high-glucose conditions in patients and animal models, so these non-coding RNAs play important roles in glucose metabolism regulation and also in neurogenesis, neurodevelopment, and neurodegeneration processes [22]. Herein, this study aimed to investigate the possible role of neurodegeneration-related miRNAs in diabetes, so we measured some miRNAs that have regulatory effects in neural death [23], such as miR-23a, miR-27a, miR-107, miR-128, miR-191, and miR-802 in diabetic and non-diabetic samples. According to the results (Fig. 3), high-glucose conditions in diabetic patients cause a harsh dysregulation in miRNA levels. According to the results, miR-23a was reduced in blood samples related to both types of diabetes (p < 0.0001****) while miR-23a expression level is similar in T1DM and T2DM, and statistical analysis could not detect a significant difference (p = 0.86 ns). Conversely, miR-27a remarkably increased by nearly 2-fold in diabetic patients (p < 0.0001****) without any significance according to the type of diabetes (p = 0.99 ns). However, the circular concentration of miR-107 also increased significantly in diabetic patients (p < 0.0001****, p < 0.005***). This miRNA expression level is different between diabetic patients according to the type of diabetes, and Fig. 3 confirmed that miR-107 in patients suffering from T2DM is significantly less than in T1DM (p < 0.0005, **). The results of the current study revealed that miR-128 was reduced in T1DM in comparison with control (p < 0.0001****), and there was no significant difference between T2DM and control (p = 0.749 ns). Therefore, according to the miR-128 expression level, T1DM and T2DM are different from each other, and Fig. 3 revealed a high value of this miRNA in T2DM. Patients suffering from high-glucose disorder revealed a higher concentration of miR191 than non-diabetic samples (control vs T1DM, p = 0.0004*** and control vs T2DM, p < 0.0001****). T1DM samples also have reduced levels of miR191 in comparison with T2DM patients (p < 0.0001****). Finally, miR-802 expression level increased in T1DM by nearly 3-fold rather than in control (p < 0.0001****). While there is no significant difference between control and T2DM (p = 0.829 ns). Therefore, T2DM and T1DM are different from each other as regards miR-802 level (p < 0.0001****).
Figures & Tables

According to previous studies, high glucose toxicity and also insulin deficiency could be considered as the main reasons for neurological abnormalities manifested by patients suffering from different types of DM [2,3,22]. This study aimed to evaluate the possible cross-talk between miRNA regulation and neurohormones, which could affect both the metabolism rate and neural function simultaneously in two common types of DM. One of the main regulators of metabolism is thyroid hormones, and our results revealed no significant differences between the three experimental groups. Previous reports showed thyroid dysfunction is more common in T2DM patients in comparison with the general population [24]. As in previous studies, hyperglycemia could affect thyroid function by mediating the TSH hormone and impairing the conversion of T4 to T3 in the peripheral tissues [21]. These kinds of variances in human samples are usual and possibly are related to the method sensitivity, stage, duration of disease, and also genetic diversity. Within the nervous system, under shifting cellular conditions, neurohormones modify the function and number of miRNAs to meet the needs of specific neurons [25]. On the other hand, by controlling the production and release of neurohormones that drive synaptic plasticity, brain growth, etc., miRNAs influence neuronal communication [26]. Evaluation of interactions between neurodegeneration-related miRNAs and neurohormones revealed that miR-27a, miR-107, and miR-191 upregulation is associated with a significant reduction in dopamine and oxytocin circular content in both types of diabetes. Previous reports confirmed that miR-27a overexpression could suppress dopamine synthesis and release in human dopaminergic-like M17 cells through direct inhibition of PTEN-induced putative kinase 1 (PINK1) protein [27]. According to a similar study, upregulation of miR-27a is one of the main causes of the loss of dopaminergic cells in Parkinson’s disease due to direct inhibition of PINK1, which plays critical roles in mitochondrial homeostasis, spine morphogenesis, and sensitivity against excitotoxicity in neurons [28]. Interestingly, according to previous experiments, PINK1 expression is remarkably reduced in patients suffering from T2DM and also obesity [27]. Our results suggest that PINK1 down-regulation is due to the up-regulation of miR-27a, which was observed in both types of diabetes and could cause mitochondrial-mediated neurodegeneration and dopamine depletion [28]. Incubation of primary epithelial cells in 25 mM glucose medium causes a harsh depolarization in the cell membrane and also a significant reduction in PINK1 expression, which suppresses mitophagy and leads cells to early apoptosis [29]. Prajapati et al., [30] found a decrease in the miR-23a and miR-128 associated with neuroinflammation and dopaminergic cell death by targeting mitochondrial complex I, which also leads to dopamine depletion. According to our knowledge, miRNAs are molecules that regulate genes through base complementarity with their cognate mRNAs; therefore, each miRNA could have multiple targets involved in the regulation of more than one process. According to the published data by Juan et al., [31], miR-128 could also regulate pituitary gland function and oxytocin synthesis and release; the data of the current study showed a positive correlation between miR-128 and oxytocin contents. While we could not find the direct relationship between miR-23a, miR-27a, miR-107, miR191, and oxytocin expression and release in the literature, the current study’s data revealed that reduced circular oxytocin is accompanied by increased concentration of miR-27a, miR-107, miR191, and also reduced level of miR-23a that suggests a possible regulatory relationship between oxytocin and miRNAs needs to be investigated further. According to previous experiments, dopamine and oxytocin contents are in balance with each other, and oxytocin regulates dopamine release in extrahypothalamic regions [32]. Oxytocinergic neurons also express dopamine receptors and are susceptible to modulation by dopamine [32]. Thus, it appears that miRNAs and neurohormones have reciprocal regulation effects. Those that were studied here or those not investigated yet are the main causes of neural abnormalities observed in diabetic patients and related animal models. Accordingly, an imbalance of miRNAs imposed by hyperglycemia is one of the main risk factors for Alzheimer’s, Parkinson’s, and other neurodegenerative diseases. The other risk factor is the non-enzymatic glycation of different macromolecules, such as proteins, that could change enzymes and receptors’ biological functions [4]. The third risk factor is the circulating insulin level, which markedly influences neurogenesis and neurodevelopment [3].
Diabetes-associated neurodegeneration is a challenging problem in patients and related animal models. Here, certain difficulties were resolved by focusing on the interaction between neurohormones and miRNAs. The results of the present study revealed a positive feedback loop between neurodegeneration-related miRNAs, neurohormones, and hyperglycemia. This loop revealed high glucose toxicity as an effective risk factor in the initiation and development of neurodegeneration and some related diseases because the miRNA network reorganization, which is imposed by high glucose toxicity, triggers neural cell death. Consequently, this study emphasizes the important role of neurohormone dysregulation in diabetes, and introduces them as potential therapeutic targets to reduce the neurological side effects of DM. Overall, there is a reciprocal regulatory relationship between miRNAs and neurohormones, which needs further investigation to clarify some ambiguities and could also be considered as a target for treatment in DM.
Author Contributions
Dr. Leila Sadeghi conceived of the presented idea. Dr. Mohammad Ali Hosseinpour Feizi developed the theory, and Toqa Alabyadh performed the experiments. Dr. Leila Sadeghi verified the analytical methods. All authors discussed the results and contributed to the final manuscript.
We would like to certify that there are no known conflicts of interest related to this article and that the work was not significantly funded, which could have influenced its results.![]()
References
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0

