

Full Length Research Article
Plastic Biodegradation Potential of Soil Mangrove Mold Isolated from Wonorejo, Indonesia
Nengah Dwianita Kuswytasari1, Alfia Rahma Kurniawati2, Aunurohim1, Nur Hidayatul Alami1, Enny Zulaika1, Maya Shovitri1, Nimerta Kumari3, Arif Luqman1*
Adv. life sci., vol. 10, no. 2, pp. 228-238, June 2023
*– Corresponding Author: Arif Luqman (arif.luqman@its.ac.id)
Authors' Affiliations
2. Department of Biology Education, Raden Fatah Islamic State University, Palembang – Indonesia
3. Institute of Microbiology, University of Sindh, Jamshoro – Pakistan
[Date Received: 07/12/2022; Date Revised: 15/04/2023; Date Published Online: 30/06/2023; Date Updated: 11/09/2025]
Abstract![]()
Introduction
Methods
Results
Discussion
References
Abstract
Background: Plastic has become one of the most significant environmental threats, especially in mangrove areas due to their non-biodegradable nature. This indicates that there is a need to find alternative degradation methods for these materials, such as the use of mold. Therefore, this study aims to isolate potential plastic degradation mold from the Wonorejo mangrove sediments.
Methods: The isolates were identified based on their morphological characteristic and ITS rDNA sequence. Plastic degradation ability of the samples was evaluated using parameters of plastic dry weight loss, FTIR pattern, SEM, and AFM image.
Results: A total of eight sediment mold were identified, including Perenniporia spp., Trametes polyzona, Aspergillus terreus, Porostereum spadiceum, Leptosphaerulina chartarum, Aspergillus fumigatus, and Hypocreales sp. Among these molds, the best activity was exhibited by Aspergillus terreus, which degraded 12.5% LDPE and 4.9% white plastic after 30 days, followed by Perenniporia sp. with 10.6% LDPE and 3.4% white plastic degradation. Furthermore, the SEM and AFM images showed that the surface of plastic was damaged after incubation. There were also several attenuations of particular peaks, which indicated the occurrence of chemical changes along plastic chains belonging to the C-H alkane group at wavenumber 2914.94, 2847.38, and 1471 cm-1. Enzyme produced by mold in this study were measured qualitatively. Perenniporia sp. produced laccase, manganese peroxidase, alkane hydroxylase, and lipase. Trametes polyzona also secreted similar enzyme, except lipase, while Porostereum spadiceum was negative for alkane hydroxylase. Aspergillus terreus isolates were positive for lipase and alkane hydroxylase, but Aspergillus fumigatus only produced lipase.
Conclusion: In this study, 8 mold with plastic biodegradation potential by producing various enzyme were identified.
Keywords: Biodegradation, Enzyme, Mangrove; Mold; Plastic; Responsible consumption & production
Introduction![]()
Plastic wastes have become a global threat to the environment due to their low degradation rate. Previous reports have also shown that the global production volume of these synthetic materials is likely to increase by 25 million tons per year [1]. Furthermore, plastic wastes have a half-life of tens or even hundreds of years [2], making their presence an environmental issue that needs to be handled seriously by all parties.
One of the ways to manage plastic waste is biodegradation, involving the breakdown of polymers into carbon dioxide, water, and biomass through the action of microorganisms that consume these materials as carbon sources [3]. Moldare the primary decomposers in nature and can form colonies in various environmental conditions. Furthermore, the saprotrophic properties of these decomposers, as well as the ability to live in stressed conditions enable them to produce several enzyme that degrade recalcitrant compounds [4].
Several enzyme have been reported to be involved in plastic biodegradation, including ligninolytic enzyme, such as manganese peroxidase, lignin peroxidase, and laccase, which are used for breaking polyethylene [5-8]. Alkane hydroxylase is often used to degrade hydrocarbons [9-12], while lipase can break downplastic[13-15]. Previous studies showed that each enzyme has its specific catalytic activity on the substrate.
Mangrove areas are located at the border between land and marine environment and are characterized by the presence of mangrove forests. This unique location makes these regions vulnerable to the accumulation of plastic wastes from the mainland. The existence of these wastes in mangrove areas has led to the discovery of microorganisms, particularly mold, that are capable of degrading plastic. Furthermore, Nathania and Kuswytasari (2013) identified some mangrove mold with the ability to degrade bioplastic Poly Hydroxy Butyrate (PHB)[16].
Mold characterization is generally carried out phenotypically based on the macroscopic and microscopic structures [17,18], but these characteristics can be influenced by environmental conditions. An effective alternative to overcome this challenge is the use of genotypic characterization based on the Internal Transcribed Spacer (ITS) rDNA area. This technique is faster and more appropriate for mold analysis compared to 18S rDNA [19] because the degree of variation in the ITS area is higher than the other rDNA genes (Small Subunit (SSU) and Large Subunit (LSU)). Among the various kinds of ITS rDNA standard primers, the universal ITS1 and ITS4 primers are often used because they allow selective amplification of mold sequences [20].Therefore, this study aims to identify and determine the ability of the Wonorejo mangrove soil mold to degrade plastic as well as to detect enzyme-producing mold that play a role in biodegradation process.
Methods![]()
Plastic material
This study used plastic as a substrate for biodegradation assay. Plastic bags were obtained from Bunga Api, Surabaya, Indonesia, while Low-density polyethylene (LDPE, type GF57836537) was purchased from Aldrich in Huntingdon, England. Furthermore, each plastic was cut into small sizes measuring 20 mm x 20 mm squares.
Isolation of soil mangrove mold from Wonorejo
Mold were isolated from plastic surfaces, which were buried in the Wonorejo mangrove soil. Approximately 1 g of partially degraded plastic waste samples from mangrove sediments were then transferred to a 50 ml Falcon tube containing 10 ml sterile distilled water, followed by vigorous vortexing for 5 min. Subsequently, 0.1 ml of sample in the dilution tube was spread onto plates containing Potato Dextrose Agar (PDA) (HiMedia, Mumbai, India) media, with incubation at room temperature for 3-7 days [21]. The growing mold colonies were purified 3 times by colony propagation [22] and then inoculated into a PDA tube medium.
Phenotypic characterization
Mold identification was carried out based on macroscopic and microscopic morphology. Macroscopic observation included surface and reverse side color, colony surface, the presence or absence of exudate drops, radial furrow, and concentric circles [23]. Meanwhile, the microscopic observation consisted of the shape of hyphae or mycelium, the shape of the spore, and the presence or absence of a septum on the hyphae. Phenotypic data were then compared with key books, such as identification keys for the introduction of general tropical fungi [23], Illustrated Genera of Fungi Imperfecti [24], Pictorial Atlas of Soil and Seed Fungi Morphologies of Cultured Fungi and Key to Species [18], and Phylogenetic and taxonomic studies on The Genera Penicillium and Talaromyces [25].
DNA isolation
DNA isolation was performed based on the Promega Kit Extraction protocol with slight modifications. The fungal mycelium (7 days) was taken from the surface of the PDA with a sterile toothpick of 0.1-1 mg and placed into a 1.5 ml microcentrifuge tube. It was then suspended with 300 µl of nuclei lysis solution and 300 µl of glass bead. The mixture obtained was then vortexed vigorously at 3000 rpm for 5 min, followed by the addition of 100 μl protein precipitation solution. Subsequently, the tube was inverted about 10 times and incubated on ice for 5 min. The sample was centrifuged at 13500 for 3 min, and the supernatant containing the DNA was transferred to a clean 1.5ml microcentrifuge tube filled with isopropanol in a ratio of 1:1. The tube content was then gently mixed by inversion until thread-like strands of DNA formed a visible mass. The sample was centrifuged at 13500 for 2 minutes to precipitate the DNA. The supernatant was then discarded using a pipette, while the pellet was mixed with 70% ethanol, and the tube was gently inverted to wash the DNA pellet, followed by centrifugation for 1 minute. Ethanol was discarded and dried at 65 °C for 10 min, and the pellet was mixed with 100 μl of DNA rehydration solution and 1 μl RNase, followed by incubation at 65 °C for 1 hour. The DNA concentration was calculated using a Nanodrop at 260/280 nm, while the remaining DNA was stored at -20 °C.
Amplification of ITS rRNA gene fragment
Isolates were identified based on the ITS area of DNA (rDNA) [26]. Amplification was carried out using the ITS1 universal primer (5′-TCCGTAGGTGAACCTGCGG-3′) and ITS 4 (5′-TCCTCCGCTTATTGAT-GC-3′) [27]. The total volume of the reaction mixture was 50 μl, consisting of 10 µl of 10x amplification reaction buffer, 1.5 μl dNTP (dTTP, dCTP, dATP, and dGTP), 2.5 µl of each primer (100 pmol) forward and reverse, 1 μl of DNA genome, 0.5 μl of polymerase enzyme Q5, as well as H2O, which was added to the final volume.
PCR consisted of initial denaturation at 98ᵒC for 30 seconds, followed by 35 cycles consisting of denaturation at 98ᵒC for 15 seconds, attachment at 63ᵒC for 30 seconds, and extension at 72ᵒC for 1 minute, with a final extension at 72ᵒC for 2 min. The temperature of the annealing step in the cycle was determined using the online program tmcalculator.neb.com, and the PCR amplicons varied by ±500bp. Furthermore, the PCR product was confirmed by loading a sample onto 0.8% gel electrophoresis at 135 Volts for 15 minutes. A total of 5 μl of ladder and sample was mixed with loading dye and loaded into the wells. The DNA migration results were observed under a UV transilluminator after the gel electrophoresis was flooded in ethidium bromide solution for 15 minutes. The PCR products were then purified using NucleoSpin® Gel and PCR Clean-up (Macherey-Nagel).
Bioinformatic analysis of sequence product
DNA sequences from forward and reverse primers were compiled using the Bioedit program. The sequence results were then identified to the taxa level of the species using the Basic Local Alignment Search Tool (BLAST) bioinformatics method online at the website address http://blast.ncbi.nlm.nih.gov/Blast.cgi. Subsequently, multiple alignments of the sequences were carried out using the MEGA X program [28] with the CLUSTAL W method [29]. The phylogenetic trees were constructed using the Neighbor-joining tree method [30] with a 500 bootstrap value.
Biodegradation Assay
For biodegradation assay, 90 ml of Mineral Salt (MS) medium, consisting of 100 mg/L yeast extract, 1 gr/L (NH4)2SO4, 200 mg/L MgSO4.7H2O, 0.5 mg/L Na2MoO4.2H2O, 100 mg/L NaCl, 20 mg/L CaCl2.2H2O, 10 mg/L FeSO4.7H2O, 0.5 mg/L MnSO4, 1.6 gr/L K2HPO4, and 200 mg/L KH2PO4, was added to 250 ml Erlenmeyer flask containing 3 strips of plastic films (20 mm x 20 mm). Furthermore, the flask was inoculated with 10 ml of mold starter and incubated on a rotary shaker (120 rpm) at 30 °C for 30 days[31]. Plastic films were recovered from the broth and then washed thoroughly using sterile water and 70% alcohol. The samples were placed in an oven at 80ºC for 24 hours and subjected to weight loss, SEM, FTIR, and AFM analyses.
Assessment of biodegradation
Percentage Efficiency Degradation (ED)
Biodegradation value was determined by measuring the final weight of plastic film on an analytical balance BBI-31 (BOECO, Germany). The degradation of plastic was evaluated in terms of weight loss percentage, which was calculated using the equation below [32]:

Fourier Transform Infrared (FTIR)
The analysis of chemical structure change was carried out using the FTIR. The infrared spectra of the Functional group structure of plastic films were recorded on a Nicolet Is50 spectrophotometer (Thermo Scientific, Massachusetts, USA). Furthermore, the relative peak differences in the C-C stretch, C-H bending, CH2 deformation, CH2 (symmetrical) bending, CH2 (asymmetric) bending, and CH2 stretching between the control and degraded plastic were compared. Absorbance was measured in the IR-medium region at wave numbers ranging from 400-4000 cm-1.
Scanning Electron Microscope (SEM)
Morphological changes in plastic surface were observed using SEM after the incubation period. For observation of cell morphology, plastic samples with attached cells were washed gently with distilled water gently to remove the remaining medium. Meanwhile, for observation of degradation results on plastic surfaces, the samples were washed with 2% sodium dodecyl sulphate and distilled water to remove the attached cells. The films were sputter-coated with gold using SC7620 sputter coated (EMITECH, Dubai, UAE) at 18 mA and 8 mbar, followed by visualization under SEM Evo Ma 10 (Carl Zeiss).
Atomic Force Microscopy (AFM)
Plastic damage was observed based on the topology obtained from the results of polymer degradation tests using AFM (BRUKER type N8 NEOS, Germany).
Detection of plastic degradation enzyme
Qualitative Tests for Enzyme Laccase and Manganese Peroxidase
Laccase detection was performed on a PDA medium containing 0.02% Guaiacol, while the identification of manganese peroxidase was carried out using a laccase test medium added with hydrogen peroxidase (H2O2). The samples were then incubated at room temperature for 7 days, and the formation of the reddish brown halo on the medium showed positive results [33].
Qualitative Test of Alkana hydroxylase enzyme
A qualitative test of enzyme alkane hydroxylase was carried out on the agar medium, which was coated with n-hexadecane. Furthermore, the basal medium consisted of 3.815 g K2HPO4, 0.5 g KH2PO4, 0.825 g (NH4) 2HPO4, 1.2625 g KNO3, 0.2 g Na2SO4, 0.02 g CaCl2, 0.002 g FeCl3, 0.02 g MgCl2, and 15g/L agar. Mold was inoculated into basal medium + n-hexadecane, and the growth of the colony was observed [19]. The positive control used for this process was isolates grown on a PDA medium.
Qualitative Test of Lipase Enzyme
A qualitative test of the lipase enzyme was carried out using Tween 80 as a substrate [34]. Furthermore, the medium consisted of peptone 10 g/L, 0.5 g NaCl, 0.1 g/L CaCl2.5H2O, 7g/L MgSO4.7H2O, 7 g/L KH2PO4, 20g/L agar, and 80ml/L tween. Mold was then inoculated on the medium and incubated at room temperature for 48 hours. The positive results were indicated by the presence of a clear circular precipitation area around the colony.
Results![]()
Isolation of mold from soil of the Wonorejo mangrove
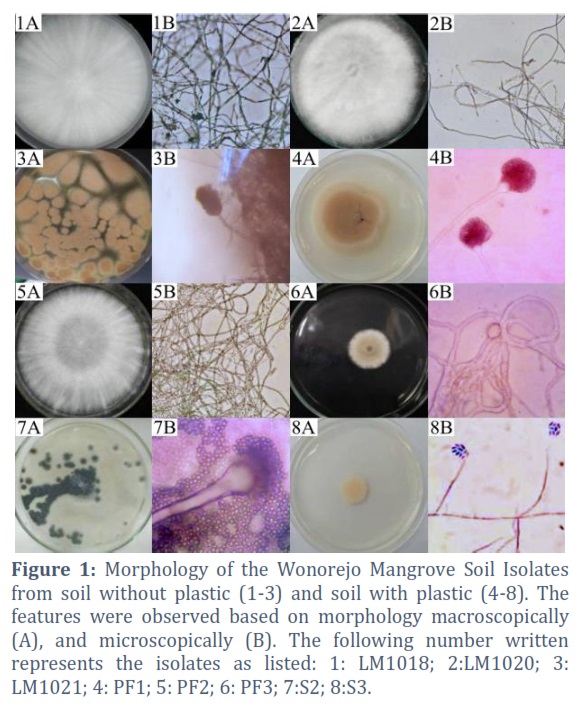
The Wonorejo mangrove area located on the east coast of Surabaya, Indonesia, was selected as the location for isolating plastic degradation soil mold. The coastal conditions, including river estuaries, caused the cause mangrove areas to be polluted by waste, especially plastic. Isolation was carried out on the Wonorejo mangrove soil with and without the intended waste. Furthermore, plastic was added to soil, and the mixture was incubated for 1 year to trigger indigenous mold to utilize the waste as a carbon source. Kuswytasari et al., (2011) had previously reported the isolation of 37 pure isolates from the Wonerejo soil without plastic [35]. In this study, three potential isolates were selected, namely LM1018, LM1020, and LM1021. Based on the isolation results from soil with waste, a total of 5 dominant pure isolates were obtained, including PF1, PF2, PF3, S2, and S3.
Conventional identification of mold was carried out phenotypically based on morphological characteristics [36]. The macroscopic and microscopic morphology of mold is presented in Figure 1. Identification can also be carried out genotypically using molecular characterization [37]. The molecular targets used were sustainableareas with variation, such as ribosomal DNA, which can be used for identification up to the species level [38].
Morphology of the Wonorejo mangrove soil isolate
The Wonorejo mangrove soil isolate colonies showed diversity in terms of color, namely white, brown, green, and yellow. The mycelia from various types of mold were observed macroscopically and microscopically in PDA medium. This morphology was then matched with a mold identification book [18,23,24]. Dark brown and granular-shaped colonies were observed in LM1021 and PF1 isolates, as shown in Figures 1.3A and 4A. Furthermore, their microscopic characteristics showed septate hyphae, yellowish-brown conidium heads, compact appearance, and columnar shape (Figures 1.3B and 4B). Other colonies observed were isolates with S2 codes, which were granular and had a light to old green coloration (Figure 1.7A). The results showed that the S2 isolates had the same microscopic characteristics, namely septate hyphae, conidiophores with a spherical shape, and colored conidia forming elongated chains (Figure 1.7B). The brownish colony surface was observed in the sample with the PF3 code, as shown in Figure 1.6A. Based on microscopic observation, the mycelium appeared transparent with branched septate hyphae, while the conidium had a round to ellipse shape (Figure 1.6B). The S3 isolates had a flat shape with a wet and pale yellow texture, as shown in Figure 1.8A. The microscopic observation showed the presence of a clear yellow coloration with uneptic and conidial mycelia, which were cylindrical or ellipsoidal. The conidiospore of the samples was also observed to form a pin-head ball, as shown in Figure 1.8B. White colonies, which were spread to the entire surface of the Petri dishes and had cotton-like surfaces were observed in LM1018, LM1020, and PF2 isolates (Figures 1.1A, 2A, and 5A). The microscopic morphology of the three samples only showed hyphae that were not septate and branched (Figure 1.1B, 2B, and 5B).
Identification of species using only morphological criteria is quite difficult, it is stated by Sierra and Henricot (2002) that there are several cases requiring information on the biochemistry and ecology of culture, and similarities in the characteristics of mycelium are very similar between different species [39]. For this reason, identification is needed based on molecular character. Molecular identification has been evaluated in several fungi groups to the species level [37].
Molecular characterization
Molecular characterization in this study was carried out to identify genotypic mold isolates. Gherbawy and Voigt (2010) stated that the incorporation of morphological and molecular-based identifications can help mold taxonomy in species-level differentiation, even in varieties [40]. The results of PCR products were obtained using a pair of universal primers, namely ITS1 and ITS4 for forward and reverse, respectively. Furthermore, the sequences obtained from each forward and reverse primer were processed by cutting the samples with low peaks using the Bioedit program.
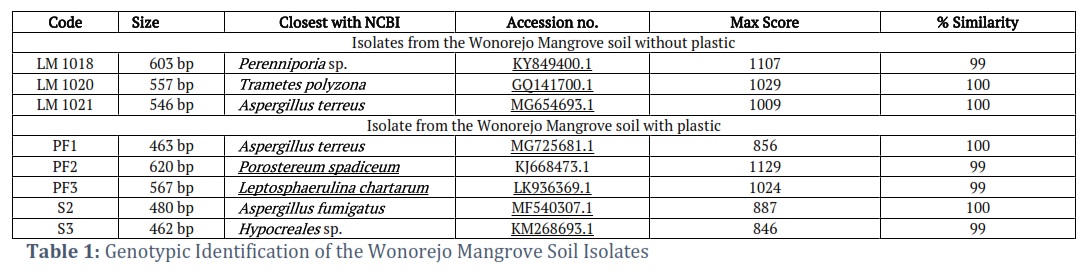
Sequences from the ITS rDNA area were then analyzed for intraspecies similarity with data in Genbank NCBI using the BLAST algorithm, as shown in Table 1. The overall similarity of queries with data on Genbank was found to be 99% and 100%. These results can then be used for identification up to the species level through comparison with the phenotypic observations of mold isolates.
A total of three isolates, namely LM1021, PF1, and S2 were identified as potentially belonging to the Aspergillus genus, with 100% identical results observed for Aspergillus terreus in LM1021 and PF1 isolates, and 100% similarity with Aspergillus fumigatus in S2. Furthermore, PF3 isolates were included in the genus Leptosphaerulina, showing identical results of 99% with Leptosphaerulina chartarum. Some of these samples also showed 99% similarity with Hypocreales sp. The results showed that three isolates included in the Basidiomycota phylum, namely LM1020, LM1018, and PF2 were identical at 100% with Trametes polyzona and 99% with Perenniporia sp. and Porostereum spadiceum.
Similarity and kinship between sequences in the Wonorejo mangrove soil isolates can be observed through phylogenetic tree construction using the neighbor-joining method, which involved distance calculation. The method was utilized to determine the position of the isolate sequence in relation to the closest sequence, which has a small base pair difference. This allowed for the visualization of the distance between each sequence in the samples [41]. The phylogeny tree construction results from the Wonorejo mangrove soil isolates are presented in Figure 2.
Alignment sequences were carried out using CLUSTAL W in the MEGA6 program along with the construction of a phylogenetic tree, as shown in Figure 2. The bootstrap value located in the branch showed the significance of the data set, which had been scrambled in predicting the same branches [41]. The results of phylogenetic trees from the Wonorejo mangrove soil isolates had a value of >70%, indicating that the prediction of kinship was significantly different and can be trusted.
The main branch after the outgroup (Coronilla sistotrema) showed the level of the phylum, with the upper and lower side representing Ascomycota and Basidiomycota phylum, respectively. In the Ascomycota phylum, three branches indicated the class level, namely Eurotiomycetes (isolates code LM1021, PF1, and S2), Dothideomycetes (isolate PF3 code), and Sordariomycetes (S3). Meanwhile, in the phylum Basidiomycota, the sequence showed the same class and order, namely Agaricomycetes and Polyporales. The results also revealed that there were three branches indicating different families, namely Phanerochaetaceae (isolate PF2 code) and Polyporaceae (isolates code LM1018 and LM1020).
Plastic Biodegradation
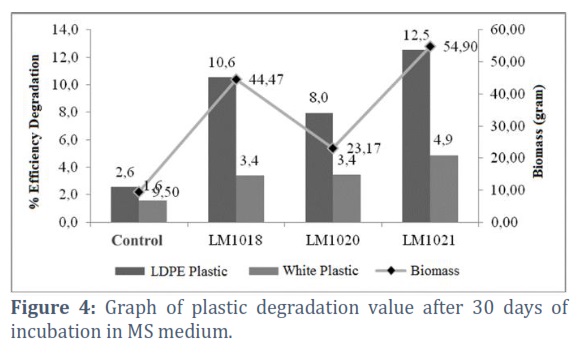
Plastic biodegradation can be seen from the observations of several parameters, such as the formation of % ED, FTIR, SEM, and AFM analysis. Biodegradation process began with biofilm formation and penetration of hyphae into plastic polymers. Furthermore, based on the observations of LDPE plastic using SEM, there was a rod formation embedded in the substrate, as shown in Figure 3. This indicated the penetration of hyphae into plastic, which marked the beginning of the degradation process. The % ED of all the isolates obtained in this study is presented in Figure 4. Based on the ANOVA test, the three isolates had a significant effect on the degradation value, but in ordinary plastic, only LM1021 had an influence. The % ED data showed higher values in LDPE plastic compared to the white variant. This was because the LDPE plastic contained a purer composition compared to the white variant containing additives.
Aspergillus terreus LM1021 had the highest %ED of 12.2% and 4.9% in LDPE and white plastic, respectively, with an incubation period of 30 days. This was positively correlated with the highest biomass produced between the two other isolates, namely 45.7 gr. Perenniporia sp. LM1018 showed % ED of 10.8% and 3.4% in LDPE white plastic respectively. Meanwhile, Trametes polyzona LM1020 produced values of 8.1% and 3.4% in LDPE and white plastic, respectively.
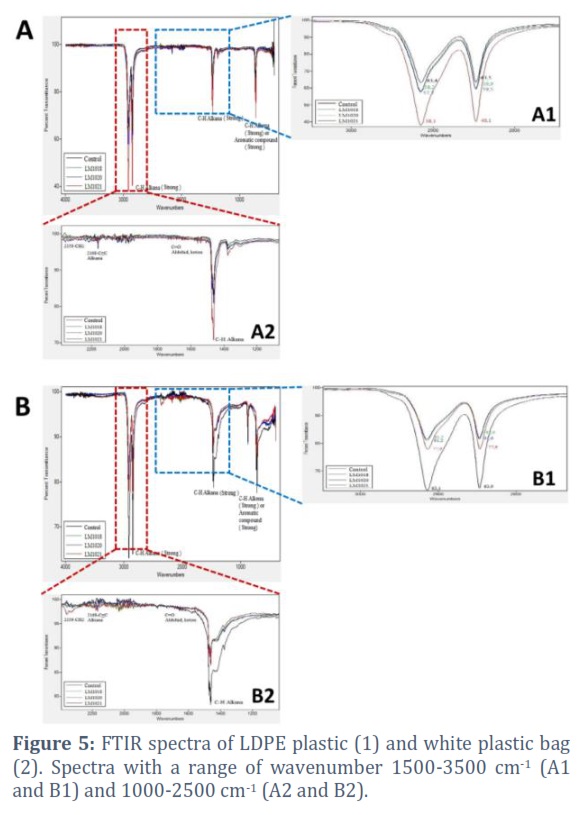
FTIR analysis on test plastic was carried out to determine structural changes that occurred in the functional group. The sensitivity of FTIR to the local molecular environment had been widely used to analyze interactions between molecules. The LDPE and white plastic showed that there was a stretch in the C-H alkane group at wavenumber 2914.94, 2847.38, and 1471 cm-1. This indicated that mold isolate catalyzed the breakdown of the polymer in the polyethylene chain. Stretching also occurred in the symmetrical C-C=C group of the aromatic ring at 1462.43 cm-1 and the C=C from the alkyne group at 2159.93 cm-1. The highest stretch was shown by isolates Aspergillus terreusLM1021, and this was in line with the high ED. The FTIR test results on degraded white plastic provide a quite different picture from the results of LDPE plastic FTIR. The transmittance intensity of the C-H alkane group on LDPE plastic after biodegradation was lower than the control, whereas in the white plastic the transmittance intensity was higher than the control. Stretching also shown on the C=C aromatic ring in 1461.30 and 1471.67cm-1 and the C-H alkane group at 2913.70 and 2846.60 cm-1 (Figure 5). SEM analysis showed changes in plastic surface after biodegradation process. Changes in plastic morphology can also be seen from plastic surface roughness using the Atomic Force Microscopic (AFM). Table 2 showed variations in the sample morphology based on AFM and SEM analyses. SEM results revealed the presence of significant changes in all films with different destructive patterns. The LDPE plastic surface after biodegradation exhibited grinding and peeling, while the white plastic showed holes (Table 2). Changes in the surface of LDPE and white plastic by isolates Perenniporia spp. LM1018 (Table 2) showed evenly distributed damage. The structural variations in the Trametes polyzona LM1020 (Table 2) contained rod fragments attached to the surface, which allowed hyphae penetration. The initial plastic peeling stage was characterized by indentations on plastic surface. Furthermore, surface changes were evident in isolates Aspergillus terreusLM1021, which illustrated the occurrence of erosion by random peeling. Similar forms of damage are also reported by a previous study [42].
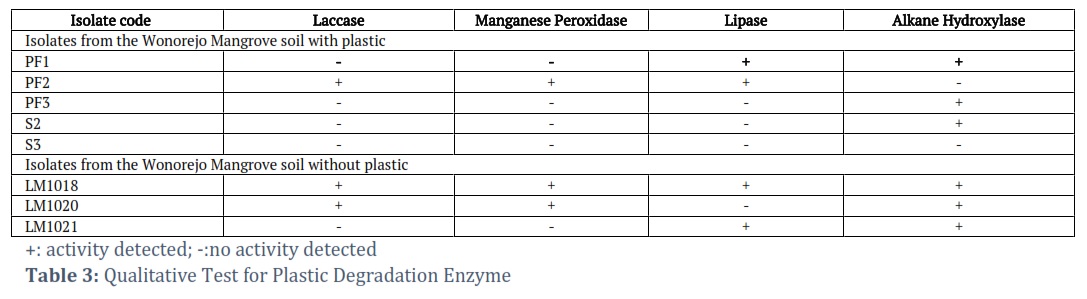
The 3-dimensional topography of LDPE plastic is presented in Table 2 (Left). The topography was indicated by color changes, where the lighter the surface, the higher the topographic condition, and vice versa. The AFM test results showed that the topography of the LDPE plastic from biodegradation test results (Table 2) had several dark-colored valley areas compared to LDPE plastic control with more bright regions. The AFM observations reported by Ojha et al. also showed similar results [43]. The results of enzyme detection test of laccase, manganese peroxidase, lipase, and alkane hydroxylase are presented in Table 3. The detection of lipase was carried out using Tween 80, where positive results were indicated by the presence of a zone of precipitation around the microorganisms that produced enzyme in the agar medium. The Wonorejo mangrove soil isolates with the ability to produce lipases included Perenniporia sp. (LM1018), Aspergillus terreus (LM1021 and PF1), Porostereum spadiceum (PF2), and Aspergillus fumigatus (S2).
Alkane hydroxylase detection was carried out based on the method proposed by Liu et al. (2014), which involved growing mold isolates on BS (Basal Salt) agar medium coated with n-hexadecane [19]. Positive results were indicated by growth in the test medium. The positive controls used for the procedure consisted of mold isolates grown on a PDA medium. The test results showed the six Wonorejo mangrove mold isolates with the ability to grow on the UI medium, namely LM1018, LM1020, PF1, PF3, and S2. However, the observation showed that PF2 and S3 could not grow on the test medium.
Perenniporia sp. (LM1018), Trametes polyzona (LM1020), and Porostereum spadiceum (PF2) were classified as white rot fungi. Based on the results, they had potential to produce lignin peroxidase enzyme, as indicated by the positive results in the qualitative laccase and manganese peroxidase tests. The qualitative test results of laccase and manganese peroxidase in isolates capable of producing ligninolytic enzyme showed a brown zone around the microorganisms. This was due to the oxidation reaction on the guaiacol substrate (laccase) and guaiacol + H2O2 substrate (manganese peroxidase).
Figures & Tables



The phenotypic characteristics of LM1021 and PF1 showed that they can be included in the genus Aspergillus, as reported by Gandjar et al. [23]. Based on this morphology, there was a high possibility that the S2 isolates belonged to the same genus Aspergillus. There was also a possibility that the PF3 code was a member of Leptosphaerulina, and this was consistent with the report of Wu et al., [44]. Furthermore, the morphological characteristics showed that isolate S3 can be included in the genus Hypocreales. The characteristics of LM1018, LM1020, and PF2 were observed to be almost the same as the phylum Basidiomycota. Advanced identification of the phylum can be carried out genotypically based on the ITS rDNA area. Molecular characterization was carried out in the Internal area of the ITS rDNA. There were more than 90,000 fungal sequences in the ITS area, which was commonly used for fungi barcoding [45]. The results showed that the more biomass produced, the higher enzyme released, thereby increasing the degradation value. Potential of Aspergillus terreusLM1021 in degrading plastic had been reported by several studies [46-48], but there was limited information on the plasticdegradationproperties of LM1018 and LM1020 isolates. However, LM1018 and LM1020 isolates were known to have potential as lignin-degrading fungi [49-52]. The transformation of the LDPE structure pattern from the results of degradation was similar to the findings of Ojha et al., [43]. Detection of the formation or removal of carbonyl groups and double bonds using FTIR showed the occurrence of a biodegradation process [42]. The results showed that the different types of plastic had their unique degradation mechanism. Bonhomme et al., stated that polymer biodegradation was divided into two pathways, namely hydro and/or oxo biodegradation [53]. The initiation of the breakdown of the polyethylene chain was the longest and most difficult step in the degradation process. Consequently, a longer incubation time was required to produce sufficient quantities of carbonyl groups to continue the decomposition process. Albertsson et al., (2004) reported that only a few biologically oxidized polyethylene chains were used by intracellular enzyme in β-oxidation and cutting of methylene units [54]. According to Bonhomme et al., the peaks became wider along with the rate of degradation due to production of several monomeric forms and oxidative yields of polyethylene [53]. Morphological damage in the form of indentation and erosion that appears on biodegradation results occurred due to the absence of uniform distribution of the polymer matrix. The morphological changes observed served as the confirmation of plastic fragility caused by mold culture. Lipase was one of the hydrolase enzyme that was responsible for the degradation of hydrocarbons [13-15]. It also had the ability to catalyze hydrolytic cleavage, leading to a decrease in hydrophobicity on the polymer surface [55]. The precipitation zone formed was the crystallization of calcium salt with fatty acids produced from the hydrolysis of Tween [34]. The involvement of alkane hydroxylase in the degradation process of plastic, especially polyethylene had been reported by Jeon and Kim (2015)[56]. The degradation process occurred through the alkane hydroxylase system, which can reduce the alkane chain with the alkane monooxygenase present in the first stage of the pathway, leading to alkane terminal carbon hydroxylation [57]. Lignin was a natural aromatic polymer of woody and recalcitrant biomass. Furthermore, white rot fungi had been reported to be the most effective and widely studied lignin-degrading microorganisms for plastic degradation [58]. The three main enzyme involved in the ligninolytic system included Lignin peroxidase (LiP), Manganese peroxidase (MnP), and laccase [15]. Several studies have proven that the degradation of polyethylene was carried out by ligninolytic enzyme. Laccase was the main enzyme secreted by lignin-degrading mold, and it had the ability to catalyze various polyaromatic compounds and non-aromatic substrates. According to Mayer and Staple (2002), it was responsible for the oxidation of hydrocarbon chains from polyethylene. Manganese peroxidase had been reported to be a lignin-degrading enzyme that used manganese as an important cofactor, especially as a regulator of MnP production and an active mediator in the MnP catalytic cycle [59]. Manganese played an important role in the degradation system of polyethylene, and MnP was involved in the degradation activities by fungi in a culture containing sufficient manganese [60].
Based on the results, the eight Wonorejo mangrove soil isolates had been identified using their morphological and molecular characteristics, namely Perenniporia sp. (LM1018), Trametes polyzona (LM1020), Aspergillus terreus (LM1021), Aspergillus terreus (PF1), Porostereum spadiceum (PF2), Leptosphaerulina chartarum (PF3), Aspergillus fumigatus (S2) and Hypocreales sp. (S3). Isolate Aspergillus terreus LM1021 showed the highest potential in plastic degradation with a % ED value of 12.5% and 4.9% in LDPE and white plastic, respectively. Perenniporia sp. LM1018 had a % ED of 10.6% and 3.4% in LDPE and white plastic, respectively. Meanwhile, Trametes polyzonaLM1020 isolates showed values of 8% and 3.4% in LDPE and white plastic. The Wonorejo mangrove soil isolates, which were capable of producing laccase and manganese peroxidase were LM1018, 1020, and PF2. LM1018, LM1021, PF1, and PF2 can produce lipase enzyme, while LM1018, LM1020, LM1021, PF1, PF3, and S2 secreated Alkana hydroxylase.
Acknowledgement
The authors are grateful to Indonesia Endowment Fund for Education (LPDP-Lembaga Pengelola Dana Pendidikan) and Kemenristekdikti for the financial support. The authors are also grateful to Dr.Ali Rohman, M.Si., Ph.D. for the assistance in tutorial data sequencing, as well as Prof.Dr. Purkan, S.Si.,M.Si. and Prof. Dr. Afaf Baktir, MS for assisting with enzyme data analysis.
Conflict of Interest
The authors declare that there is no conflict of interest.
![]()
References
- Kaseem M, Hamad K, Deri F. Thermoplastic starch blends: A review of recent works. Polymer Science Series A, (2012); 54(2): 165-176.
- Obruča S. Controlled production and degradation of selected biomaterials. Brno: Vysoké učení technické v Brně, Fakulta chemická, (2010).
- Klun U, Friedrich J, Kržan A. Polyamide-6 fibre degradation by a lignolytic fungus. Polymer degradation and stability, (2003); 79(1): 99-104.
- Naima A, Safia A, Bashir A, Geoffery R. Isolation and identification of polystyrene biodegrading bacteria from soil. African Journal of Microbiology Research, (2010); 4(14): 1537-1541.
- Ehara K, Iiyoshi Y, Tsutsumi Y, Nishida T. Polyethylene degradation by manganese peroxidase in the absence of hydrogen peroxide. Journal of wood science, (2000); 46(2): 180-183.
- Premraj R, Doble M. Biodegradation of polymers. Indian Journal of Biotechnology, (2005); 4(2): 186-193.
- Ameen F, Moslem M, Hadi S, Al-Sabri AE. Biodegradation of Low Density Polyethylene (LDPE) by Mangrove fungi from the red sea coast. Progress in Rubber Plastics and Recycling Technology, (2015); 31(2): 125-143.
- Sowmya H, Ramalingappa B, Nayanashree G, Thippeswamy B, Krishnappa M. Polyethylene degradation by fungal consortium. International Journal of Environmental Research, (2015); 9(3): 823-830.
- Fish NM, Harbron S, Allenby DJ, Lilly MD. Oxidation of n-alkanes: isolation of alkane hydroxylase from Pseudomonas putida. European journal of applied microbiology and biotechnology, (1983); 17(1): 57-63.
- Van Beilen JB, Funhoff EG. Alkane hydroxylases involved in microbial alkane degradation. Applied microbiology and biotechnology, (2007); 74(1): 13-21.
- Van Beilen JB, Li Z, Duetz WA, Smits TH, Witholt B. Diversity of alkane hydroxylase systems in the environment. Oil & gas science and technology, (2003); 58(4): 427-440.
- Mishra S, Singh S. Microbial degradation of n-hexadecane in mineral salt medium as mediated by degradative enzymes. Bioresource Technology, (2012); 111148-154.
- Skariyachan S, Megha M, Kini MN, Mukund KM, Rizvi A, et al. Selection and screening of microbial consortia for efficient and ecofriendly degradation of plastic garbage collected from urban and rural areas of Bangalore, India. Environmental monitoring and assessment, (2015); 187(1): 1-14.
- Eberl A, Heumann S, Brückner T, Araujo R, Cavaco-Paulo A, et al. Enzymatic surface hydrolysis of poly (ethylene terephthalate) and bis (benzoyloxyethyl) terephthalate by lipase and cutinase in the presence of surface active molecules. Journal of biotechnology, (2009); 143(3): 207-212.
- Bhardwaj H, Gupta R, Tiwari A. Communities of microbial enzymes associated with biodegradation of plastics. Journal of Polymers and the Environment, (2013); 21(2): 575-579.
- Nathania TR, Kuswytasari ND. Studi Potensi Isolat Kapang Wonorejo Surabaya dalam Mendegradasi Polimer Bioplastik Poly Hydroxy Butyrate (PHB). Jurnal Sains dan Seni ITS, (2013); 2(2): E55-E58.
- Larone DH, Larone DH Medically important fungi: a guide to identification. Chapter: Book Name. 1987 of publication; 196; Citeseer.
- Watanabe T Pictorial atlas of soil and seed fungi: morphologies of cultured fungi and key to species. Chapter: Book Name. 2002 of publication; CRC press.
- Liu H, Xu J, Liang R, Liu J. Characterization of the medium-and long-chain n-alkanes degrading Pseudomonas aeruginosa strain SJTD-1 and its alkane hydroxylase genes. PloS one, (2014); 9(8): e105506.
- Gardes M, Bruns TD. ITS primers with enhanced specificity for basidiomycetes‐application to the identification of mycorrhizae and rusts. Molecular ecology, (1993); 2(2): 113-118.
- Ilyas M. Isolasi dan Identifikasi Mikoflora Kapang pada Sampel Serasah Daun Tumbuhan di Kawasan Gunung Lawu, Surakarta, Jawa Tengah. Biodiversitas, (2007); 8(2): 105-110.
- Alexopoulos CJ, Mims CW, Blackwell M Introductory mycology. Chapter: Book Name. 1996 of publication; John Wiley and Sons.
- Gandjar I, Rifai MA Pengenalan kapang tropik umum. Chapter: Book Name. 1999 of publication; Yayasan Obor Indonesia.
- Barnett HL Illustrated genera of imperfect fungi. Chapter: Book Name. 2013 of publication; Burgess Publishing Company.
- Samson RA, Houbraken J Phylogenetic and taxonomic studies on the genera Penicillium and Talaromyces. Chapter: Book Name. 2011 of publication; CBS-KNAW Fungal Biodiversity Centre.
- Webb JS, Nixon M, Eastwood IM, Greenhalgh M, Robson GD, et al. Fungal colonization and biodeterioration of plasticized polyvinyl chloride. Applied and environmental microbiology, (2000); 66(8): 3194-3200.
- White T, Bruns T, Lee S, Taylor J (1990) Amplification and direct sequencing of fungal RNA genes for phylogenetics: 315-322 (in) MA INNIS, DH GELGARD, JJ SNINSKY & TJ WHITE (eds.) PCR Protocols: a Guide to Methods and Applications. San Diego, CA: Academic Press.
- Tamura K, Stecher G, Peterson D, Filipski A, Kumar S. MEGA6: molecular evolutionary genetics analysis version 6.0. Molecular biology and evolution, (2013); 30(12): 2725-2729.
- Thompson JD, Higgins DG, Gibson TJ. CLUSTAL W: improving the sensitivity of progressive multiple sequence alignment through sequence weighting, position-specific gap penalties and weight matrix choice. Nucleic acids research, (1994); 22(22): 4673-4680.
- Saitou N, Nei M. The neighbor-joining method: a new method for reconstructing phylogenetic trees. Molecular biology and evolution, (1987); 4(4): 406-425.
- Pramila R, Ramesh KV. Biodegradation of low density polyethylene (LDPE) by fungi isolated from marine water a SEM analysis. Afr J Microbiol Res, (2011); 5(28): 5013-5018.
- Hosseini S, Salari M, Jamalizadeh E, Khezripoor S, Seifi M. Inhibition of mild steel corrosion in sulfuric acid by some newly synthesized organic compounds. Materials Chemistry and Physics, (2010); 119(1-2): 100-105.
- Vantamuri A, Kaliwal B. Isolation, screening and identification of laccase producing fungi. Int j pharm bio sci, (2015); 6(3): 242-250.
- Gupta P, Upadhyay L, Shrivastava R. Lipase Catalyzed-transesterification of Vegetable Oils by Lipolytic. Research Journal of Microbiology, (2011); 6(3): 281-288.
- Kuswytasari N, Shovitri M, Andriyadi R. Soil Molds Diversity in The Coastal Wonorejo Surabaya; 2011.
- Burdsall HH, Banik M, Cook ME. Serological Differentiation of Three Species of Armillaria and Lentinula Edodes by Enzyme-Linked Immunosorbent Assay Using Immunized Chickens as a Source of Antibodies. Mycologia, (1990); 82(4): 415-423.
- Iwen PC, Hinrichs SH, Rupp ME. Utilization of the internal transcribed spacer regions as molecular targets to detect and identify human fungal pathogens. Medical mycology, (2002); 40(1): 87-109.
- Schwarz P, Bretagne S, Gantier J-C, Garcia-Hermoso D, Lortholary O, et al. Molecular identification of zygomycetes from culture and experimentally infected tissues. Journal of Clinical Microbiology, (2006); 44(2): 340-349.
- PÉRez-Sierra A, Henricot B. Identification of fungal species beyond morphology. Mycologist, (2002); 16(2): 42-46.
- Gherbawy Y, Kesselboth C, Elhariry H, Hoffmann K (2010) Molecular barcoding of microscopic fungi with emphasis on the mucoralean genera Mucor and Rhizopus. Molecular identification of fungi: Springer. pp. 213-250.
- Dharmayanti N. Filogenetika molekuler: metode taksonomi organisme berdasarkan sejarah evolusi. Wartazoa, (2011); 21(1): 1-10.
- Esmaeili A, Pourbabaee AA, Alikhani HA, Shabani F, Esmaeili E. Biodegradation of low-density polyethylene (LDPE) by mixed culture of Lysinibacillus xylanilyticus and Aspergillus niger in soil. Plos one, (2013); 8(9): e71720.
- Ojha N, Pradhan N, Singh S, Barla A, Shrivastava A, et al. Evaluation of HDPE and LDPE degradation by fungus, implemented by statistical optimization. Scientific Reports, (2017); 7(1): 1-13.
- Wu Q, Li Y, Li Y, Gao S, Wang M, et al. Identification of a novel fungus, Leptosphaerulina chartarum SJTU59 and characterization of its xylanolytic enzymes. PLoS One, (2013); 8(9): e73729.
- Hibbett DS, Ohman A, Glotzer D, Nuhn M, Kirk P, et al. Progress in molecular and morphological taxon discovery in Fungi and options for formal classification of environmental sequences. Fungal biology reviews, (2011); 25(1): 38-47.
- Raaman N, Rajitha N, Jayshree A, Jegadeesh R. Biodegradation of plastic by Aspergillus spp. isolated from polythene polluted sites around Chennai. J Acad Indus Res, (2012); 1(6): 313-316.
- Ibrahim IN, Maraqa A, Hameed KM, Saadoun IM, Maswadeh HM. Assessment of potential plastic-degrading fungi in Jordanian habitats. Turkish Journal of Biology, (2011); 35(5): 551-557.
- Singh V, Dubey M, Bhadauria S. Biodeterioration of polyethylene high density by Aspergillus versicolor and Aspergillus terreus. Journal of Advanced Laboratory Research in Biology, (2012); 3(1): 47-49.
- Arantes V, Milagres AM, Filley TR, Goodell B. Lignocellulosic polysaccharides and lignin degradation by wood decay fungi: the relevance of nonenzymatic Fenton-based reactions. Journal of Industrial Microbiology and Biotechnology, (2011); 38(4): 541-555.
- Dashtban M, Schraft H, Syed TA, Qin W. Fungal biodegradation and enzymatic modification of lignin. International journal of biochemistry and molecular biology, (2010); 1(1): 36.
- Vasina DV, Moiseenko KV, Fedorova TV, Tyazhelova TV. Lignin-degrading peroxidases in white-rot fungus Trametes hirsuta 072. Absolute expression quantification of full multigene family. PLoS One, (2017); 12(3): e0173813.
- Knežević A, Milovanović I, Stajić M, Vukojević J. Potential of Trametes species to degrade lignin. International Biodeterioration & Biodegradation, (2013); 8552-56.
- Bonhomme S, Cuer A, Delort A, Lemaire J, Sancelme M, et al. Environmental biodegradation of polyethylene. Polymer degradation and Stability, (2003); 81(3): 441-452.
- Albertsson A-C, Karlsson S. The influence of biotic and abiotic environments on the degradation of polyethylene. Progress in Polymer science, (1990); 15(2): 177-192.
- Tribedi P, Sil AK. Bioaugmentation of polyethylene succinate-contaminated soil with Pseudomonas sp. AKS2 results in increased microbial activity and better polymer degradation. Environmental Science and Pollution Research, (2013); 20(3): 1318-1326.
- Jeon HJ, Kim MN. Functional analysis of alkane hydroxylase system derived from Pseudomonas aeruginosa E7 for low molecular weight polyethylene biodegradation. International Biodeterioration & Biodegradation, (2015); 103141-146.
- Rojo F. Degradation of alkanes by bacteria. Environmental Microbiology, (2009); 11(10): 2477-2490.
- Kale SK, Deshmukh AG, Dudhare MS, Patil VB. Microbial degradation of plastic: a review. Journal of Biochemical Technology, (2015); 6(2): 952-961.
- Mayer AM, Staples RC. Laccase: new functions for an old enzyme. Phytochemistry, (2002); 60(6): 551-565.
- Iiyoshi Y, Tsutsumi Y, Nishida T. Polyethylene degradation by lignin-degrading fungi and manganese peroxidase. Journal of wood science, (1998); 44(3): 222-229.
This work is licensed under a Creative Commons Attribution-Non Commercial 4.0 International License. To read the copy of this license please visit: https://creativecommons.org/licenses/by-nc/4.0